Extracellular vesicle-derived microRNAs in renal cell carcinoma: biological roles and clinical applications
Jiahui He, Lanfeng Wang, Bingqing Yu, Wengbin Huang, Chen Ouyang, Mingcan Zhou, Rong Hu, Zhiping Chen

TL;DR
This review explores how microRNAs in extracellular vesicles contribute to kidney cancer and their potential for diagnosis and treatment.
Contribution
The paper systematically reviews the biological roles and clinical potential of EV-derived miRNAs in renal cell carcinoma.
Findings
EV-derived miRNAs play roles in tumor progression and drug resistance in RCC.
These miRNAs show promise as biomarkers for diagnosis and prognosis in kidney cancer.
They may also help predict treatment response and guide immune regulation strategies.
Abstract
Renal cell carcinoma (RCC) is a common malignant tumour of the urinary system, characterised by high heterogeneity and a tendency to metastasise, with poor prognosis in advanced patients. Although surgical resection and targeted therapies such as tyrosine kinase inhibitors and immune checkpoint inhibitors have significantly improved survival outcomes in some patients, drug resistance and recurrence remain clinical challenges. In recent years, extracellular vesicles (EVs) and the microRNAs (miRNAs) they carry have emerged as a research hotspot due to their critical roles in tumour initiation, progression, immune regulation, and drug resistance. This systematic review summarises the biological functions of EVs-derived miRNAs in renal cell carcinoma and their potential applications in clinical diagnosis and treatment, with a focus on their value in diagnosis, prognosis, immune regulation,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
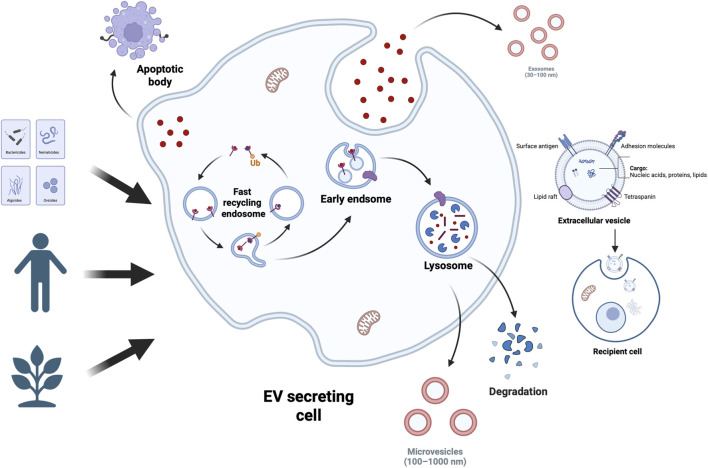 FIGURE 1
FIGURE 1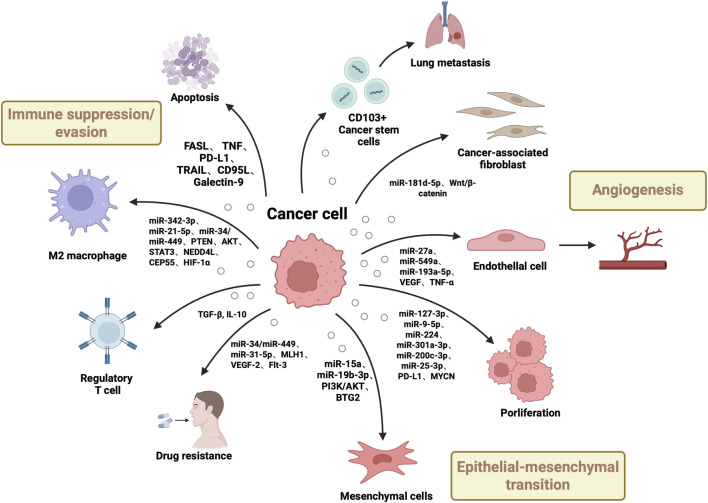 FIGURE 2
FIGURE 2| Sample source | Number of samples | Dysregulated miRNAs | Physiological effect | Target genes | miRNA profiling | REF |
|---|---|---|---|---|---|---|
| Plasma-derived Evs | 32 ccRCC patients with localized disease (before and after surgery) and in 37 patients with metastatic disease | miR-301a-3p↑miR-1293↓ | miR-301a-3p activates PI3K/AKT to promote progression and metastasismiR-1293 acts as a tumor suppressor | PTEN | Small RNA-seq, RT-qPCR |
|
| Cell-derived Evs | 3 RCC patient pairs (tumor tissues: CAFs/adjacent normal tissues: NFs) | miR-181d-5p↑ | Promotes tumor cell proliferation, migration, and invasion | RNF43 | Small RNA-seq, qRT-PCR |
|
| Cell-derived Evs | N/A | miR-127-3p↑ | Promotes tumor cell migration, invasion, and metastasis and is associated with poor prognosis | MYCN | RT-qPCR、Next-Generation Sequencing |
|
| Serum-derived EVs | 35 ccRCC patients (high expression)/31 ccRCC patients (low expression) | miR-9-5p↑ | Promotes tumor cell proliferation and migration, positively associated with TNM stage and Fuhrman grade | SOCS4 | qRT-PCR |
|
| Cell-derived Evs | N/A | miR-549a↓ | Enhances vascular endothelial permeability and angiogenesis and promotes lung metastasis formation | HIF-1α | RT-qPCR, Small RNA-seq |
|
| Tissue-derived Evs | 53 paired ccRCC tissues and adjacent normal tissues (31 stage I–II/22 stage III–IV patients) | miR-15a↑ | Promotes tumor cell proliferation, migration, invasion, and EMT and aggravates ccRCC progression via PI3K/AKT activation | BTG2 | Microarray, RT-qPCR |
|
| Tissue-derived Evs | 133 CCRCC patients (stage I–II, non-metastatic)/76 CCRCC patients (stage III–IV, metastatic) | miR-19b-3p↑ | Enhances tumor cell migration, invasion, and metastasis and CD103⁺ exosomes preferentially target lung tissue to promote metastasis | PTEN | RT-qPCR, Small RNA-seq |
|
| Tissue-derived Evs | 51 histologically confirmed ccRCC tissue samples (including 23 paired adjacent non-cancerous tissues) | miR-193a-5p↑ | Enhances tumor cell migration and angiogenesis and promotes tumor progression and metastasis | TIMP2 | RT-qPCR |
|
| Cell-derived Evs | N/A | miR-342-3p↑ | Promotes tumor cell proliferation, migration, invasion, and metastasis by inhibiting NEDD4L and stabilizing CEP55 to activate PI3K/AKT/mTOR signaling | NEDD4L, CEP55 | RT-qPCR |
|
| Cell-derived Evs | N/A | miR-21-5p↑ | Promotes tumor cell proliferation, migration, invasion, and distant metastasis by downregulating PTEN and activating Akt signaling to induce EMT. | PTEN | RT-qPCR |
|
| Cell-derived Evs | N/A | lncARSR↑→miR-34/miR-449 | Induces macrophage polarization from M1 to M2 and promotes phagocytosis, angiogenesis, and tumor progression | STAT3 | RT-qPCR |
|
| Plasma-derived Evs | 40 metastatic RCC patients (sorafenib treatment) | miR-31-5p↑ | Promotes sorafenib resistance and enhances tumor cell proliferation and survival under drug pressure | MLH1 | RT-qPCR, TaqMan miRNA probes |
|
| Biomarker | Clinical application | Source | Number of cases/ | Sensitivity% | Specificity% | AUC | Characterization | Quantification technique | Normalizer | REF |
|---|---|---|---|---|---|---|---|---|---|---|
| miR-210 | Diagnostic of CCRCC | Serum | 82 ccRCC patients/80 healthy controls | 70.000 | 62.200 | NA | Flow cytometry analysis and immunofluorescence | qRT-PCR | U6 |
|
| miR-1233 | 81.000 | 76.000 | ||||||||
| miR-210 | Diagnostic of CCRCC | Serum | 45 ccRCC patients/30 healthy controls | 82.500 | 80.000 | NA | TME; Western blot | qRT-PCR | miR-16-5p |
|
| miR-224 | Diagnostic of CCRCC | Serum | 108 ccRCC patients | NA | NA | 0.833 | TME; Immunoprecipitation and western analysis | qRT-PCR | U6 |
|
| miR-9-5 | Diagnostic of CCRCC | Serum | 35 ccRCC patients (high expression)/31 ccRCC patients (low expression) | NA | NA | NA | Western blot | qRT-PCR | snRNAU6 |
|
| miR-149-3p | Diagnostic of CCRCC | Serum | 22 ccRCC patients/16 healthy controls | 75.000 | 77.300 | 0.719 | TEM; NTA | qRT-PCR | miR-16-5p |
|
| miR-424-3p | 75.000 | 72.700 | 0.773 | |||||||
| miR-92a-1-5p | 87.500 | 81.800 | 0.832 | |||||||
| hsa-miR-200c-3p | Diagnostic of CCRCC | 2D and 3D cell culture | NA | NA | NA | NA | TEM | qRT-PCR | GAPDH |
|
| hsa-miR-25-3p | ||||||||||
| has-301a-3p | ||||||||||
| miR-224-5p | Diagnostic of CCRCC | urine | NA | NA | NA | NA | TE,; NTA; Western blot | qRT-PCR | miR-16; RNU48 |
|
| miR-30c-5p | Diagnostic biomarker of early-stage ccRCC | urine | 70 early-stage (T1aN0M0) ccRCC patients/30 early-stage prostate cancer (T1N0M0) patients/30 early-stage bladder cancer (T1N0M0) patients/30 hralthy controls | 68.57 | 100 | 0.819 | TEM; NTA | qRT-PCR | Not Specified |
|
| Combination of miR-126-3p-miR-449a | Diagnostic of CCRCC | Serum | 81 ccRCC patients/24 patients with benign lesions/33 healthy controls | 60.6 | 100 | 0.82 | TEM | qRT-PCR | miR-16-5p + miR-106a-5p |
|
| Combination of miR-126-3p-miR-34b-5p | 67.3 | 82.8 | 0.8 | |||||||
| Combination of miR-126-3p-miR-486-5p | 52.9 | 95.8 | 0.79 | |||||||
| Combination of miR-25-3p-miR-34b-5p | 73.1 | 79.3 | 0.76 | |||||||
| Combination of miR-21-5p-miR-34b-5p | 74 | 72.4 | 0.76 | |||||||
| Combination of miR-150-5p-miR-126-3p | 61.5 | 82.8 | 0.76 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsExtracellular vesicles in disease · MicroRNA in disease regulation · Circular RNAs in diseases
1 Introduction
Renal cell carcinoma (RCC) is one of the most common malignant tumours of the urinary system, with over 400,000 new cases diagnosed globally each year, making it one of the leading causes of cancer-related deaths. The incidence of RCC is on the rise in both developed and developing countries (Chen et al., 2023; Sun et al., 2023; Grünwald et al., 2024). Based on histological characteristics, the most common subtypes of RCC are clear cell type, papillary type (Type I and Type II), and chromophobe type, accounting for 70%–90%, 10%–15%, and 3%–5% of all renal malignant tumours, respectively. RCC accounts for 2%–3% of all cancers and is the most lethal urogenital system cancer, with a mortality rate of 30%–40%, compared to approximately 20% for bladder cancer and prostate cancer. The incidence of RCC continues to rise, varying globally with higher rates in developed countries than in developing countries (Stepanovska Tanturovska et al., 2023). Despite recent advances in surgical techniques and the introduction of novel targeted therapies such as tyrosine kinase inhibitors (TKIs) and immune checkpoint inhibitors (ICIs), the prognosis for patients with advanced RCC remains poor, with a five-year survival rate below 15%. The primary challenges in current treatment include the presence of drug resistance mechanisms and the lack of reliable early diagnostic and prognostic biomarkers (Chen et al., 2020; Motzer et al., 2022; Delcuratolo et al., 2023).
Extracellular vesicles (EVs) have been demonstrated to serve as crucial intercellular communication mediators within the tumour microenvironment (TME) (Kalluri and LeBleu, 2020; Urabe et al., 2020; Xiao Y. et al., 2020; Huang et al., 2021; Marar et al., 2021; Fusco et al., 2024). Enclosing diverse bioactive components including proteins, lipids, and nucleic acids, EVs modulate recipient cell behaviour, thereby influencing numerous physiological and pathological processes such as TME formation and evolution (Maacha et al., 2019; Ortiz, 2021; Kita and Shimomura, 2022; Kumar et al., 2024; Rai et al., 2024). The mechanisms of EV-mediated cellular communication are diverse, encompassing activation of membrane surface receptors, Messenger RNA (mRNA) transport and translation, transfer of microRNA (miRNAs) and target mRNAs, delivery of functional proteins, and activation of signalling pathways via internalisation (D'Souza-Schorey and Clancy, 2012; Svensson and Belting, 2013; Maeda et al., 2023). Among these, miRNAs have emerged as a current research focus due to their crucial role in gene expression regulation and their extensive involvement in tumour progression, metastasis, angiogenesis, and immunomodulation. Valadi et al. were the first to demonstrate that miRNAs can be transported between cells via extracellular vesicles while retaining biological activity in recipient cells (Valadi et al., 2007). miRNA constitutes one of the most abundant RNA components within EVs, accounting for up to 40% of RNA in human plasma EVs as revealed by RNA sequencing (Mittelbrunn et al., 2011; Makarova et al., 2021; Yang et al., 2021; Xiong et al., 2023). Studies have shown that miRNAs are predominantly enclosed within extracellular vesicles (EVs), where vesicular encapsulation protects them from nuclease degradation in body fluids, thereby facilitating intercellular communication (Kogure et al., 2019). EVs are widely distributed across various biological fluids, including plasma, saliva, urine, milk, and cerebrospinal fluid. Notably, the abundance of miRNAs carried by EVs from milk and cerebrospinal fluid may exceed that of plasma-derived EVs (Tabatabai et al., 2025). Whether derived from normal or tumour tissues, EVs serve as carriers for both miRNA and pathogenic proteins, potentially contributing to the onset and progression of certain diseases (Saeedi et al., 2021; Doncheva et al., 2022; Kumar et al., 2024) (Figure 1).
EV Types and Sources. Extracellular vesicles (EVs) encapsulate bioactive components like proteins, nucleic acids, and lipids—lipids act as structural elements and reflect cellular traits. Cells across tissues use EVs for intercellular communication, releasing them into body fluids. A large share of human EVs comes from stem cells; EVs are also actively produced and released by organisms from plants to bacteria.
microRNAs (miRNAs) are a class of small non-coding RNAs that primarily suppress gene translation by targeting messenger RNAs (mRNAs), and participate in diverse biological processes including cell differentiation, proliferation, apoptosis, and development (Lu et al., 2005; Filella and Foj, 2017; Selvaskandan et al., 2023). In malignant tumours, miRNAs may function as either tumour suppressors or oncogenes, exhibiting characteristic alterations in their expression levels (Guil and Esteller, 2009; Hill and Tran, 2021; Li B. et al., 2021). The differential expression of miRNAs between normal and cancerous cells renders them ideal candidate molecules for tumour biomarkers (Chow et al., 2010). miRNAs within tumour-derived EVs can further enhance the invasive and metastatic capabilities of tumour cells by modulating the tumour microenvironment (TME) (Nishida-Aoki and Ochiya, 2015; Tkach and Théry, 2016).
Recent studies have revealed significant differences in the expression of EVs-miRNA between healthy individuals and RCC patients (Nawaz et al., 2014), suggesting broad application prospects in non-invasive diagnosis and prognostic assessment. Furthermore, engineered miRNA delivery systems based on EVs have demonstrated potential therapeutic value (Rädler et al., 2023). The high stability of miRNAs in bodily fluids such as plasma and urine render them ideal molecules for liquid biopsy. Nevertheless, technical challenges persist in clinical implementation, including standardization of isolation methods, target specificity, and potential off-target effects. This paper aims to provide a systematic review of research progress on EVs-miRNA in RCC, focusing on their role in tumour biology, potential as diagnostic biomarkers, and therapeutic applications. It also explores challenges and future directions in their clinical translation.
2 The biological basis of EVs and miRNAs
Extracellular vesicles are a class of membrane-bound nanovesicles actively secreted by cells, carrying diverse biomolecules such as proteins and nucleic acids, particularly microRNA (miRNA), which play crucial roles in tumourigenesis and progression. These vesicles can be readily extracted from peripheral blood and other bodily fluids, rendering them potential tools for non-invasive tumour diagnosis (Raposo and Stoorvogel, 2013; Kogure et al., 2019; Munir et al., 2020). The biological effects of extracellular vesicles on surrounding or distant target cells are primarily determined by their biomolecular cargo. Their lipid composition includes sphingolipids, cholesterol, phosphatidylserine, saturated fatty acids, and sphingosine—substances also prevalent in the plasma membrane (Trajkovic et al., 2008; Skotland et al., 2020). Research indicates that sphingosine plays a direct role in the formation of internalised vesicles (ILVs) within the lumen of multivesicular bodies (MVBs). Inhibition of neutral sphingomyelinase significantly reduces extracellular vesicle release, further validating this lipid’s critical function in extracellular vesicle biogenesis (Menck et al., 2017).
The proteome of extracellular vesicles primarily comprises proteins involved in membrane transport, such as tetramembrane proteins (CD63, CD81, CD82, and CD9), whose recruitment depends on the ALIX and ESCRT-III pathways (Larios et al., 2020). Furthermore, extracellular vesicles are rich in heat shock proteins (Hsp60, Hsp70, Hsp90), integrins, and class II major histocompatibility complex (MHC II) molecules (Clayton et al., 2005). Notably, extracellular vesicles do not simply represent the protein expression profile of the parent cell; rather, they selectively enrich certain proteins through specific mechanisms. Among these, ubiquitination is recognised as a key regulatory step mediating protein binding to the ESCRT complex and facilitating their incorporation into vesicles (Larios et al., 2020).
miRNAs are regulated both by their biogenesis mechanisms and by the sequence characteristics of the miRNAs themselves. Although the ESCRT system plays a central role in the formation of multi-vesicular bodies (MVBs) and EV release, studies indicate that knocking down key ESCRT proteins does not affect miRNA content (Kosaka et al., 2010). Interestingly, while knocking down the ESCRT-III-associated protein Alix does not affect total extracellular vesicle release, it leads to a significant reduction in miRNA content (Iavello et al., 2016).
Certain miRNA sequences contain conserved domains known as EXO-motifs, which bind to specific RNA-binding proteins such as hnRNPA2B1 and SYNCRIP, thereby facilitating their incorporation into extracellular vesicles (Santangelo et al., 2016). Previous studies have identified short motif sequences (e.g., GGAG in miR-198 and UGCA in miR-601) that regulate miRNA loading into extracellular vesicles, with point mutations altering these motifs significantly impacting miRNA efficiency (Villarroya-Beltri et al., 2013). Recent studies have further revealed an association between AGO2 and extracellular vesicle-associated miRNAs, with AGO2 being identified in extracellular vesicle proteomics via mass spectrometry (MS) or Western blotting analyses (Goldie et al., 2014; Zhang J. et al., 2015). Knockout of AGO2 was found to reduce the types or abundance of preferentially exported miRNAs in extracellular vesicles derived from HEK293T cells (Guduric-Fuchs et al., 2012). Further investigations indicate that YBX1 (Y-box protein I) also binds to miR-223 and miR-144, regulating their packaging into vesicles (Ung et al., 2014; Shurtleff et al., 2016). Collectively, these findings suggest that specific sequences within miRNAs may guide their incorporation into extracellular vesicles.
Extensive research indicates that miRNAs within extracellular vesicles participate in the formation of multiple tumour types and serve as sensitive biomarkers for cancer diagnosis (Aguilar-Hernandez et al., 2021; Lu et al., 2021; Romano et al., 2021; Maruoka et al., 2022; Wang et al., 2022; Genova et al., 2024; Kural et al., 2024). Compared to free-floating miRNAs, those derived from extracellular vesicles exhibit enhanced stability, owing to the protective double-layered membrane structure that shields them from degradation by endogenous nucleases or phagocytes such as macrophages. The nanoscale dimensions and membrane protective mechanisms of extracellular vesicles prolong the half-life of miRNAs in bodily fluids, enhancing their biological functionality (Sun et al., 2022). Upon release into bodily fluids, extracellular vesicles may be internalised by neighbouring or distant cells, where the encapsulated miRNAs can regulate a range of biological processes including immune evasion, tumour microenvironment modulation, angiogenesis, metastasis, and drug resistance development (Sun et al., 2018). Consequently, the role of EVs-miRNAs in cancer progression has garnered significant attention. For instance, Fabbri (2012) demonstrated that miRNAs within lung cancer-derived extracellular vesicles promote tumour cell proliferation and metastasis by activating Toll-like receptors (TLRs). Zhou et al. (2014) further reported that extracellular vesicles secreted by tumour cells carrying miR-105 disrupt tight junctions between vascular endothelial cells, compromising the natural barrier and thereby promoting metastasis. Additionally, studies on renal cell carcinoma (RCC) have identified circulating EVs enriched with multiple miRNAs associated with metastasis and invasion, including miR-200c, miR-92, miR-141,miR-19b, miR-29a, miR-29c, miR-650, and miR-151 (Chow et al., 2010; Grange et al., 2011). These miRNAs hold promise as biomarkers for renal carcinoma progression, demonstrating significant research and clinical application value.
3 The function of EVs carrying miRNAs in renal cell carcinoma
3.1 Tumorigenesis and progression
In recent years, multiple studies have progressively revealed the pivotal regulatory role of miRNAs carried by EVs in the development and progression of clear cell renal cell carcinoma (ccRCC). DISA et al. discovered that PTEN serves as a common target gene for hsa-miR-301a-3p, hsa-miR-200c-3p, and hsa-miR-25-3p. Studies indicate that these miRNAs are significantly enriched in plasma extracellular vesicles during the presence of primary tumours, with their expression levels markedly decreasing following tumour resection. This suggests that EVs-miRNAs may regulate tumour progression at the post-transcriptional level by activating the PI3K/AKT signalling pathway (Dias et al., 2020). Notably, hsa-mir-301a-3p exhibited a declining trend post-tumour resection, with its levels continuing to decrease during follow-up. However, its expression significantly increased in the metastasis cohort, suggesting this miRNA may play a crucial role in metastasis and holds potential as a prognostic biomarker. In contrast, the EV-derived levels of hsa-miR-1293 progressively increased after tumour resection until follow-up yet were markedly reduced in the metastatic cohort. This indicates the miRNA’s potential tumour-suppressing function and its possibility as a biomarker for metastatic disease in ccRCC patients (Dias et al., 2020). Among these, hsa-miR-301a-3p exhibited a consistent decrease in expression following surgery but was significantly elevated in patients with metastasis during follow-up, indicating its potential involvement in metastatic dissemination and its utility as a prognostic biomarker. Conversely, hsa-miR-1293 displayed increased EV expression post-surgery but was significantly downregulated in metastatic cases, suggesting its potential tumor-suppressive role and prognostic relevance in identifying metastatic ccRCC (Dias et al., 2020). Ding et al. using small RNA sequencing, demonstrated that miR-181d-5p was highly enriched in EVs derived from cancer-associated fibroblasts (CAFs) in RCC patients. This miRNA directly suppressed RNF43 expression in RCC cells and activated the Wnt/β-catenin pathway, thereby enhancing cancer stemness and promoting tumor progression (Ding et al., 2022). Additionally, RAB27 A/B has been shown to regulate EV-miRNA secretion. Decreased RAB27A expression was closely associated with lymph node metastasis and poor prognosis in RCC (Chen et al., 2012; An et al., 2019). Song et al. further reported that elevated RAB27A expression promoted RCC cell secretion of miR-127-3p, which, through EV-mediated transfer, upregulated MYCN expression and enhanced tumor invasiveness (Song et al., 2024). miR-9-5p has also been implicated in RCC progression. Song et al. found that it was significantly elevated in serum from advanced RCC patients and positively correlated with TNM stage and Fuhrman grade. Functional assays demonstrated that EV-derived miR-9-5p promoted proliferation and invasion of A-704 cells by downregulating SOCS4, both in vitro and in vivo, supporting its role as a diagnostic and prognostic biomarker (Song et al., 2020).
Further in vitro and in vivo studies confirmed that ccRCC-derived EVs transmit miR-27a, which suppresses its target gene SFRP1 while enhancing vascular endothelial growth factor (VEGF) and tumour necrosis factor-α (TNF-α) expression, thereby promoting RCC cell viability, migration, and angiogenesis (Hou et al., 2021). Xuan et al. observed significantly downregulated miR-549a expression in TKI-resistant ccRCC cells and their extracellular vesicles. Extracellular vesicle-derived miR-549a inhibits tumour angiogenesis and reduces endothelial cell migration by binding to the 3′-UTR region of HIF-1α, thereby suppressing its expression (Xuan et al., 2021).
Li et al. further observed that miR-15a, upregulated in EVs, enhances the epithelial-mesenchymal transition (EMT) capacity of ccRCC cells by downregulating BTG2 and activating the PI3K/AKT pathway (Li D. Y. et al., 2021). Furthermore, Wang et al. observed that in metastatic ccRCC patients, cancer stem cell (CSC) extracellular vesicles induce EMT by transporting miR-19b-3p to tumour cells and suppressing PTEN gene expression. CD103+-guided CSC extracellular vesicles target cancer cells and organs, conferring enhanced lung metastatic potential to ccRCC. Consequently, CD103+ extracellular vesicles also emerge as a potential metastatic diagnostic biomarker (Wang L. et al., 2019).
3.2 Immune regulation
Tumour-associated macrophages (TAMs) play a pivotal role in regulating the tumour microenvironment (TME) and promoting tumour initiation and progression (Pollard, 2004). Increasing evidence indicates that TAMs are not only one of the predominant cell types within the TME, but also participate in the complex processes of cancer through multiple mechanisms including immune suppression, promotion of tumour progression, metastasis, and drug resistance (Pathria et al., 2019; Wei et al., 2019). Macrophages can be categorised into pro-inflammatory M1 and immunosuppressive M2 types based on their functional state. TAMs exhibit phenotypic and functional characteristics closer to M2 macrophages, which are strongly associated with tumour-promoting properties (Boutilier and Elsawa, 2021). Multiple studies demonstrate that TAM infiltration correlates closely with poor prognosis across various cancer types, including RCC (Komohara et al., 2011; Zhou et al., 2015; Chen et al., 2017). Consequently, targeting TAMs presents a potential therapeutic strategy that may offer novel insights into the interactions between the tumour microenvironment and tumour cells (Pyonteck et al., 2013; Ries et al., 2014). In RCC, TAMs have been demonstrated to promote tumour cell migration and tumour growth (Kadomoto et al., 2019; Schnetz et al., 2020).
Regarding specific mechanisms, research has revealed that upregulation of HIF-1α in macrophages promotes miR-193a-5p expression. This microRNA is subsequently transported to RCC cells via extracellular vesicles, targeting the 3′-untranslated region (3′-UTR) of TIMP2 mRNA. This downregulates TIMP2 expression, thereby enhancing tumour angiogenesis and invasive capacity. Inhibition of miR-193a-5p in extracellular vesicles derived from TAMs has been shown to significantly attenuate RCC progression and metastasis, offering a novel therapeutic approach targeting TAM-associated miRNAs (Liu et al., 2022). In a ccRCC model, Feng et al. discovered that extracellular vesicles derived from M2-polarised macrophages transport miR-342-3p to target and inhibit NEDD4L, thereby blocking the ubiquitination and degradation of CEP55 and activating the PI3K/AKT/mTOR pathway. This ultimately enhances the proliferation, migration, and invasive capacity of RCC cells. This research offers fresh perspectives for developing therapeutic targets in RCC (Feng et al., 2021). Zhang et al. further indicated that extracellular vesicles derived from M2 macrophages serve as key mediators enhancing RCC cell migration and invasive potential. The miR-21-5p they enrich promotes distant tumour metastasis by downregulating PTEN expression and activating the Akt signalling pathway. In vitro and in xenograft models, this miRNA consistently demonstrated metastatic-promoting capabilities. Notably, application of miR-21-5p inhibitors reversed these pro-metastatic effects, opening novel pathways for intervening in TAM-mediated metastatic behaviour and offering a novel therapeutic strategy for preventing RCC metastasis (Zhang Z. et al., 2022). Furthermore, research indicates that extracellular vesicles derived from ccRCC cells can transport long non-coding RNA (lncARSR), activating the miR-34/miR-449-STAT3 signalling pathway. This induces the transformation of M1 macrophages towards an M2 phenotype, enhancing their phagocytic activity and promoting angiogenesis, thereby accelerating tumour progression (Zhang W. et al., 2022).
3.3 Drug resistance
With the widespread application of multi-targeted kinase inhibitors in the treatment of advanced ccRCC, the issue of drug resistance has progressively become a key obstacle affecting therapeutic efficacy. Sorafenib is a commonly used oral multi-targeted tyrosine kinase inhibitor in clinical practice. Its mechanism of action primarily involves inhibiting multiple angiogenesis and tumour proliferation-related signalling pathways, including: VEGFR-2, VEGFR-3, platelet-derived growth factor receptor-β (PDGFR-β), RAF-1,c-Kit, and FMS-like tyrosine kinase 3 (Flt-3) (Yu et al., 2015).
Research by He et al. revealed that tumour-derived extracellular vesicles can promote sorafenib resistance by transporting miR-31-5p. They further demonstrated that this microRNA targets the 3′-UTR of the MLH1 gene, leading to its downregulation and thereby inducing a sorafenib-resistant response (Yu et al., 2015). Additional studies have reported that miR-31-5p-enriched extracellular vesicles in ccRCC models mediate resistance signals by directly targeting the DNA mismatch repair-associated gene MutL homolog 1 (MLH1), thereby enhancing the sorafenib-resistant phenotype. Consequently, both miR-31-5p and its target genes may serve as predictive biomarkers and therapeutic targets for sorafenib resistance (He et al., 2020). Qu et al. further discovered that extracellular vesicles secreted by RCC cells deliver IncARSR, which promotes sunitinib resistance by competitively binding miR-34/miR-449, thereby enhancing AXL and c-MET expression in RCC (Qu et al., 2016).
In summary, EV-carried miRNAs exert core regulatory roles in RCC initiation, progression, immune modulation, and drug resistance mechanisms. By targeting multiple signalling pathways (e.g., PI3K/AKT, Wnt/β-catenin, STAT3), they influence cancer cell proliferation, migration, invasion, and metastatic nodule formation. Certain miRNAs (e.g., miR-301a-3p, miR-21-5p,miR-342-3p) are closely associated with prognosis, whilst miR-193a-5p, miR-9-5p, and miR-127-3p play crucial roles in immune regulation and invasive metastasis mediated by tumour-associated macrophages (TAMs) and cancer stem cells (CSCs). Furthermore, miR-31-5p has been demonstrated to contribute to sorafenib resistance by downregulating MLH1 expression, suggesting its potential for predicting and overcoming targeted therapy resistance. As the functional role of EV-associated miRNAs is increasingly elucidated, their clinical value as non-invasive biomarkers and therapeutic intervention targets becomes increasingly evident, offering broad prospects for the early diagnosis, dynamic monitoring, and personalised treatment of renal cell carcinoma (Figure 2, Table 1).
Roles of Renal Cell Carcinoma-Derived Extracellular Vesicles (EVs) in Tumorigenesis. This figure shows renal carcinoma EVs drive tumor development via multiple pathways: delivering growth-promoting miRNAs/oncogenes to boost cancer cell proliferation/dissemination, transferring molecules to induce epithelial-mesenchymal transition, conveying pro-angiogenic factors for neovascularization, and transporting immunosuppressive molecules to enable immune evasion, while interacting with stromal cells to remodel the tumor microenvironment.
4 EVs-miRNA serves as a biomarker for RCC
In recent years, liquid biopsy has garnered significant attention in both clinical practice and fundamental research as a non-invasive tool for tumour detection. Particularly in the early diagnosis of renal tumours, accurately distinguishing between benign and malignant small renal masses (SRMs) is crucial for determining the need for surgical intervention. Although the diagnostic accuracy of tissue biopsy continues to improve (Marconi et al., 2016), its invasive nature and procedural limitations remain obstacles to widespread clinical adoption. In contrast, miRNAs carried by EVs possess inherent protective mechanisms due to their membrane-enclosed structure, exhibiting enhanced stability and detection reproducibility in biological fluids such as plasma, serum, and urine. This offers promising clinical application prospects. Consequently, liquid biopsy strategies based on EVs-miRNA hold potential as a safer, more sensitive, and reproducible diagnostic approach for renal cancer. Simultaneously, identifying ccRCC patients at high metastatic risk is crucial for developing personalised monitoring plans, optimising adjuvant treatment decisions, and enabling early intervention against metastatic lesions. However, clinically validated molecular biomarkers for risk stratification in this patient cohort remain scarce. Notably, despite advances in targeted therapies such as TKIs and ICIs, predictive indicators for treatment efficacy remain limited (Linxweiler and Junker, 2020). Consequently, exploring EVs-miRNAs with high stability, specificity, and reproducibility as liquid biopsy biomarkers has become a core research direction for precision diagnosis and treatment of RCC. Previous studies have demonstrated that EV-associated miRNAs remain stable without degradation in preservation solutions under hypothermic ischemia conditions (4 °C) during liver transplantation (Vidal-Correoso et al., 2024). Ma et al. further showed that glycosylated extracellular vesicles preserve their miRNA cargo without degradation under storage at −80 °C and remain stable for up to 7 days at 4 °C (Ma et al., 2023). In addition, Muth et al. evaluated the effects of freeze–thaw cycles and room temperature incubation on plasma EV-miRNAs, and found that appropriate pre-analytical processing (e.g., platelet removal) markedly improves the stability and accuracy of EV-miRNA analysis (Muth et al., 2018). Collectively, these findings suggest that EV-associated miRNAs are feasible candidates for use as routine biomarkers. Numerous studies have reported that miRNAs in extracellular vesicles demonstrate significant potential in RCC diagnosis, prognostic assessment, and treatment response prediction (Butz et al., 2016; De Palma et al., 2016; Zhang et al., 2018; Song et al., 2019; Wang X. et al., 2019).
miR-210 and miR-1233, isolated from serum extracellular vesicles, have been validated as diagnostic biomarkers for clear cell renal cell carcinoma (ccRCC), exhibiting sensitivities of 70% and 81%, respectively, alongside specificities of 62.2% and 76.0%. Notably, these biomarkers demonstrate a significant decline following nephrectomy. Furthermore, Wang and colleagues reported that miR-210 could identify RCC with 82.5% sensitivity and 80.0% specificity. In this context, higher miR-210 levels were observed in more advanced cases and those with higher Fuhrman grading, independent of gender or age (Zhang et al., 2018; Wang X. et al., 2019). Fujii et al. further demonstrated that high expression of Exo-miR-224 correlates strongly with poor prognosis in ccRCC patients, manifesting as reduced survival times and accelerated tumour progression. Compared to the low-expression group, the high-expression Exo-miR-224 group exhibited significantly shorter progression-free survival, cancer-specific survival, and overall survival. In multivariate analysis, elevated Exo-miR-224 levels emerged as a significant prognostic risk factor across all studies. Co-incubation of primary renal cell carcinoma (RCC) cell lines with extracellular vesicles derived from metastatic RCC cell lines enhanced cellular proliferation and invasive capacity while markedly reducing apoptotic cell proportions. Intracellular miR-224 levels were significantly upregulated in primary renal carcinoma cell lines. Extracellular miR-224 in extracellular vesicles influences patient prognosis and represents a potential prognostic biomarker in ccRCC patients (Fujii et al., 2017). Dias et al. confirmed that multiple miRNAs (miR-301a-3p, miR-200c-3p, miR-25-3p) play a crucial role in sustaining ccRCC cell proliferation by targeting PTEN and activating the PI3K/AKT pathway. The expression level of hsa-miR-301a-3p derived from EVS was again elevated in metastatic patients, highlighting its potential as a biomarker for metastatic disease (Dias et al., 2020; Alves et al., 2024). Findings by Alves et al. further support hsa-miR-200c-3p, hsa-miR-25-3p, and hsa-miR-301a-3p in extracellular vesicles as potential biomarkers for monitoring disease aggressiveness. Experiments revealed that co-inhibiting these EVs-miRNAs significantly increased PTEN expression, reduced tumour cell proliferation and migration in 2D models, and diminished spheroid size and metabolic capacity in 3D models. These EVs-miRNAs demonstrate potential as biomarkers for monitoring disease invasiveness and as therapeutic targets for ccRCC, potentially enabling more effective and personalised treatments for patients (Alves et al., 2025). Song et al. identified miR-9-5 as highly expressed in renal cancer patients’ serum, correlating with advanced TNM staging (tumour size, lymph node metastasis) and Fuhrman grade. Both in vitro and in vivo studies suggest its utility as a diagnostic biomarker and treatment response monitor (Song et al., 2020). Furthermore, Xiao et al. reported upregulation of miR-149-3p and miR-424-3p alongside significant downregulation of miR-92a-1-5p in plasma EVs, suggesting their combination holds potential for RCC screening. Plasma extracellular vesicles containing hsa-miR-92a-1-5p,hsa-miR-149-3p, and hsa-miR-424-3p in plasma extracellular vesicles may serve as potential biomarkers for detecting RCC (Xiao C. T. et al., 2020).
In urinary EVs, researchers developed a nanowire-based EV enrichment technique capable of capturing over 99% of EVs and detecting approximately 2,500 miRNAs. This approach revealed a miRNA profile similar to that in serum, with urine being a more suitable sampling source due to its proximity to the kidneys (Yasui et al., 2024). Small extracellular vesicles secreting distinct miRNA combinations, including miR-126-3p + miR-449a, miR-126-3p + miR-34b-5p, miR-126-3p + miR-486-5p, miR-25-3p + miR-34b-5p,miR-21-5p + miR-34b-5p, and miR-150-5p + miR-126-3p, have been reported as diagnostic biomarkers for clear cell renal cell carcinoma (ccRCC),with sensitivities of 60.6%, 67.3%, 52.9%, 73.1%, 74%, and 61.5%, and specificities of 100%, 82.8%, 95.8%, 79.3%, 72.4%, and 82.8% respectively (Butz et al., 2016). Qin et al. demonstrated that miR-224-5p was significantly upregulated in urine EVs from RCC patients compared to healthy volunteers. Overexpression of miR-224-5p inhibited RCC cell proliferation and induced cell cycle arrest. Urinary EVs containing miR-224-5p were identified as a potential biomarker for RCC (Qin et al., 2021). Butz et al. observed significant downregulation of miR-126-3p (P = 0.004) in urinary EVs, alongside upregulation of miRNA-150-5p, suggesting potential for tumour diagnosis using these miRNAs in SRMs.Moreover, several distinct EVs-miRNA combinations (including miR-126-3p, miR-486-5p, and miR-34b-5p) not only differentiated general ccRCC patients and SRMs from healthy participants but also distinguished benign tumour patients from ccRCC patients. These data are highly promising and may improve future surgical management decisions for SRMs (Butz et al., 2016). Moreover, Song et al. discovered that miR-30c-5p in urinary extracellular vesicles targets heat shock protein 5 (HSPA5) and inhibits ccRCC progression, exhibiting a sensitivity of 68.57% and specificity of 100%, demonstrating significant potential as a diagnostic biomarker for ccRCC (Song et al., 2019). Crentsil et al. identified miR-205 and miR-150 in extracellular vesicles as significantly differentially expressed in 786-O cells compared to controls, suggesting their utility as ccRCC extracellular vesicle biomarkers. Results from in vitro models corroborated this finding, though only miR-205 achieved statistical significance (Crentsil et al., 2018). Consequently, miR-224-5p, miR-126-3p, miR-150-5p, miR-30c-5p, miR-205,miR-486-5p, and miR-34b-5p have been reported to exhibit altered expression in urinary EVs, showing promise for the early diagnosis and differentiation of RCC and SRMs (Butz et al., 2016; Crentsil et al., 2018; Song et al., 2019). Notably, miR-224-5p demonstrates novel predictive value in immunotherapy by influencing the stability of Cyclin D1 and PD-L1 (Qin et al., 2021).
Several candidate miRNAs detected in RCC have also been implicated in other renal disorders, which challenges their disease specificity. For instance, miR-9-5p has been proposed as a non-invasive biomarker for idiopathic membranous nephropathy (IMN) (Guo et al., 2022), and miR-210 has shown diagnostic relevance in IgA nephropathy (IgAN) (Zhao et al., 2022). Such evidence indicates that single miRNAs may lack sufficient specificity for RCC diagnosis. However, their clinical value remains promising, particularly when applied as part of multi-marker panels or in combination with imaging and clinicopathological features to enhance diagnostic precision.
In summary, EVs-miRNAs in serum and urine have demonstrated favourable specificity and stability, offering broad prospects for non-invasive detection, prognostic assessment, and treatment response prediction in RCC. Future large-scale prospective studies and technical standardisation are required to advance their clinical translation, thereby providing more personalised and precise management strategies for RCC patients (Table 2).
5 The potential of EVs-miRNA in the treatment of RCC
With ongoing innovations in treatment strategies, the clinical management of RCC, particularly metastatic renal cell carcinoma (mRCC), is progressively shifting towards individualisation. However, the lack of stable therapeutic predictive biomarkers, coupled with significant tumour heterogeneity, substantially increases uncertainty regarding treatment response. In recent years, the role of miRNAs within EVs in regulating RCC biological behaviour and guiding therapy has garnered considerable attention, offering novel avenues for clinical intervention.
Indeed, for stage I tumours confined to the kidney, the five-year survival rate exceeds 90%. However, this rate drops to approximately 72.5% in stages II/III. More concerning is that around 30% of RCC patients are already at stage IV at diagnosis, with a five-year survival rate of merely 12% (Padala et al., 2020). RCC exhibits widespread resistance to conventional chemotherapy and radiotherapy, with radical or partial nephrectomy remaining the standard treatment approach (Yang and Liao, 2018). However, radical surgery may lead to renal insufficiency, increased cardiovascular event risk, and elevated mortality (Alam et al., 2019). Surgical approach must be balanced against factors including tumour stage, size, and grade (Padala et al., 2020; Marchioni et al., 2021; Spadaccino et al., 2021). Despite aggressive treatment, approximately 20%–50% of patients will progress to advanced disease (Padala et al., 2020).
In recent years, combination therapy with ICIs and anti-angiogenic TKIs has significantly improved treatment response in mRCC (Borchiellini and Maillet, 2022). Nevertheless, therapeutic efficacy remains highly variable, potentially constrained by the absence of a unified molecular classification, intratumoural heterogeneity, and differences between RCC subtypes. Currently, the IMDC risk scoring model is the only prospectively validated prognostic indicator for mRCC (Dudani et al., 2020). Concurrently, a substantial proportion of patients develop primary or acquired resistance to targeted therapies (Makhov et al., 2018). Consequently, identifying novel, highly effective, and predictable molecular biomarkers is imperative.
EVs-miRNA, as key information molecules secreted by tumour cells and released into body fluids, participate in regulating multiple tumour biological processes, particularly in remodelling the tumour microenvironment (TME) (D'Souza-Schorey and Clancy, 2012). EVs modulate the state of local fibroblasts, macrophages, and vascular endothelial cells, inducing their transformation into pro-tumour subtypes that support tumour progression (Kosaka et al., 2016; Madeo et al., 2018; Ringuette Goulet et al., 2018). Furthermore, tumour derived EVs exert remote effects, inducing the formation of ‘pre-metastatic niches’ in target organs, enhancing vascular permeability, recruiting mesenchymal stem cells, and reprogramming the local matrix (Hoshino et al., 2015; Zhang L. et al., 2015). Conversely, EVs released by stromal cells can influence tumour cell behaviour, either enhancing their invasive capacity or inducing a dormant state (Roccaro et al., 2013; Ono et al., 2014). EVs also transmit drug resistance signals between primary and metastatic sites, participating in the establishment and maintenance of resistance (Qu et al., 2016; Lobb et al., 2017).
Specific studies indicate that elevated serum or plasma expression of miR-1233, miR-221, and miR-210 correlates significantly with RCC-specific mortality risk (Wulfken et al., 2011; Dias et al., 2017),with miR-1233 further proposed as a potential therapeutic target. Yoshino et al. discovered that EVs-miR-1 significantly inhibits RCC cell proliferation, migration, and invasion. Its downregulation in RCC tissues correlates with reduced patient survival, suggesting therapeutic potential (Yoshino et al., 2022). Furthermore, upregulation of HIF-1α expression in TAMs induces high expression of miR-193a-5p, which is transported to RCC cells via extracellular vesicles. This miRNA inhibits TIMP2 expression by targeting its 3′-UTR, thereby enhancing angiogenesis and tumour invasiveness. Notably, suppressing miR-193a-5p in TAM-derived extracellular vesicles significantly slows RCC progression and metastasis, offering a novel therapeutic direction targeting TAM-associated miRNAs (Liu et al., 2022). Regarding RCC resistance mechanisms, studies reveal that resistant RCC cells secrete EVs carrying long non-coding RNAs (lncRNAs). These lncRNAs competitively bind miR-34 and miR-449, activating MET and AXL pathways to mediate distant dissemination of sorafenib resistance (Qu et al., 2016). He et al. further demonstrated that tumour-derived extracellular vesicles promote sorafenib resistance by transporting miR-31-5p. This miRNA binds to the 3′-UTR of the MLH1 gene, suppressing its expression and inducing the formation of a resistant phenotype (Yu et al., 2015). Similar studies revealed that miR-31-5p enriched in extracellular vesicles within ccRCC models directly targets the DNA mismatch repair gene MLH1, acting as a key factor in resistance signalling. This suggests miR-31-5p and its target gene MLH1 may serve as important biomarkers for predicting sorafenib resistance and therapeutic targets (He et al., 2020). Furthermore, Song et al. reported that elevated RAB27A expression promotes miR-127-3p secretion by RCC cells. This miRNA, delivered via extracellular vesicles, enhances MYCN expression levels, thereby increasing tumour invasiveness. Consequently, engineered disruption of miR-127-3p′s extracellular vesicle transport holds promise as an effective therapeutic intervention strategy for metastatic RCC (Song et al., 2024).
In summary, the role of EVs-miRNAs in regulating therapeutic response to RCC is becoming increasingly evident. Their involvement in processes such as angiogenesis, immune regulation, and drug resistance signalling confers significant potential as predictive and therapeutic biomarkers, offering new avenues for achieving precision treatment in RCC.
6 Summary
MicroRNAs (miRNAs) derived from extracellular vesicles (EVs) play a pivotal regulatory role in multiple aspects of renal cell carcinoma (RCC), including its initiation and progression, immune modulation, and the development of drug resistance. Extensive research confirms that miR-210, miR-1233, miR-224, miR-301a-3p, and miR-31-5p are significantly enriched in RCC-associated EVs, exhibiting strong correlations with tumour staging, metastatic potential, and therapeutic response. These miRNAs regulate tumour cell proliferation, migration, and immune evasion by participating in signalling pathways such as PI3K/AKT, MET/AXL, and Wnt/β-catenin. Furthermore, they influence tumour-associated fibroblasts, macrophages, and vascular endothelial cells through extracellular vesicle-mediated intercellular communication, thereby shaping the tumour microenvironment. Moreover, EVs-miRNA persist stably in bodily fluids such as blood and urine, exhibiting excellent reproducibility and detection reliability, making them highly promising non-invasive biomarkers in the liquid biopsy field.
Nevertheless, current research faces several challenges. Firstly, standardisation of EVs extraction, identification, and miRNA analysis methods remains elusive, compromising the comparability and reproducibility of research outcomes. Secondly, the inherent high heterogeneity of RCC, with miRNA expression across distinct subtypes, different patients, and even between distinct lesions within the same patient, limits the establishment of uniform biomarkers. Furthermore, many reports remain at the correlation level, with insufficient in-depth investigation into the mechanisms of action of EVs-miRNA, particularly regarding their roles in tumour immune regulation and drug resistance pathways.
Future research should prioritise multicentre, large-scale, prospective clinical studies to systematically evaluate the accuracy and clinical utility of candidate EVs-miRNA in RCC diagnosis, prognosis, and treatment response prediction. Integrating multi-omics data—including transcriptomics, proteomics, and metabolomics—holds promise for enhancing biomarker screening precision. Furthermore, artificially synthesised or engineered extracellular vesicles offer technical feasibility for targeted miRNA delivery, potentially emerging as novel therapeutic strategies against drug resistant and metastatic RCC. Combining artificial intelligence to construct multi-factor models could further elevate the clinical utility of EVs-miRNA in personalised management.
In summary, EVs-miRNA, as a stable, specific, and reproducible molecular biomarker, is progressively emerging as a crucial breakthrough for precision diagnosis and treatment of RCC. With ongoing technological advancements and deepening mechanistic research, its clinical translational application in renal cancer holds considerable promise.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aguilar-Hernandez M. M.Rincon Camacho J. C.Galicia Garcia G. (2021). Extracellular vesicles and their associated mi RN As as potential prognostic biomarkers in chronic lymphocytic leukemia. Curr. Oncol. Rep. 23 (6), 66. 10.1007/s 11912-021-01058-2 33855607 · doi ↗ · pubmed ↗
- 2Alam R.Patel H. D.Osumah T.Srivastava A.Gorin M. A.Johnson M. H. (2019). Comparative effectiveness of management options for patients with small renal masses: a prospective cohort study. BJU Int. 123 (1), 42–50. 10.1111/bju.14490 30019484 PMC 6301094 · doi ↗ · pubmed ↗
- 3AlvesÂ.Medeiros R.Teixeira A. L.Dias F. (2024). Decoding PTEN regulation in clear cell renal cell carcinoma: pathway for biomarker discovery and therapeutic insights. Biochim. Biophys. Acta Rev. Cancer 1879 (5), 189165. 10.1016/j.bbcan.2024.189165 39117092 · doi ↗ · pubmed ↗
- 4AlvesÂ.Ferreira M.Eiras M.Lima L.Medeiros R.Teixeira A. L. (2025). Exosome-derived hsa-mi R-200c-3p, hsa-mi R-25-3p and hsa-mi R-301a-3p as potential biomarkers and therapeutic targets for restoration of PTEN expression in clear cell renal cell carcinoma. Int. J. Biol. Macromol. 302, 140607. 10.1016/j.ijbiomac.2025.140607 39900161 · doi ↗ · pubmed ↗
- 5An H. J.Song D. H.Koh H. M.Ko G. H.Lee J. H.Kim D. C. (2019). RAB 27A is an independent prognostic factor in clear cell renal cell carcinoma. Biomark. Med. 13 (4), 239–247. 10.2217/bmm-2018-0336 30661368 · doi ↗ · pubmed ↗
- 6Borchiellini D.Maillet D. (2022). Clinical activity of immunotherapy-based combination first-line therapies for metastatic renal cell carcinoma: the right treatment for the right patient. Bull. Cancer 109 (2s), 2s 4–2s 18. 10.1016/s 0007-4551(22)00234-x 35760470 · doi ↗ · pubmed ↗
- 7Boutilier A. J.Elsawa S. F. (2021). Macrophage polarization states in the tumor microenvironment. Int. J. Mol. Sci. 22 (13), 6995. 10.3390/ijms 22136995 34209703 PMC 8268869 · doi ↗ · pubmed ↗
- 8Butz H.Nofech-Mozes R.Ding Q.Khella H. W. Z.SzabóP. M.Jewett M. (2016). Exosomal Micro RN As are diagnostic biomarkers and can mediate cell-cell communication in renal cell carcinoma. Eur. Urol. Focus 2 (2), 210–218. 10.1016/j.euf.2015.11.006 28723537 · doi ↗ · pubmed ↗