The autoactivity of tomato helper NLR immune proteins of the NRC clade is unaltered in Nicotiana benthamiana prf mutants
Daniel Lüdke, Hsuan Pai, AmirAli Toghani, Adeline Harant, Chih-Hang Wu, Sophien Kamoun

TL;DR
This paper shows that tomato NRC clade immune proteins can trigger cell death independently in Nicotiana benthamiana prf mutants.
Contribution
It demonstrates that autoactivity of NRC clade proteins is not affected by prf mutations in Nicotiana benthamiana.
Findings
Tomato NRC clade proteins trigger hypersensitive cell death without sensor partners.
Autoactivity remains unchanged in Nicotiana benthamiana prf mutants.
NRC clade function is independent of prf in this system.
Abstract
Nucleotide-binding and leucine-rich repeat immune receptors of the NRC clade act in one direction, with autoactive helpers able to trigger hypersensitive cell death without their sensor partners.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1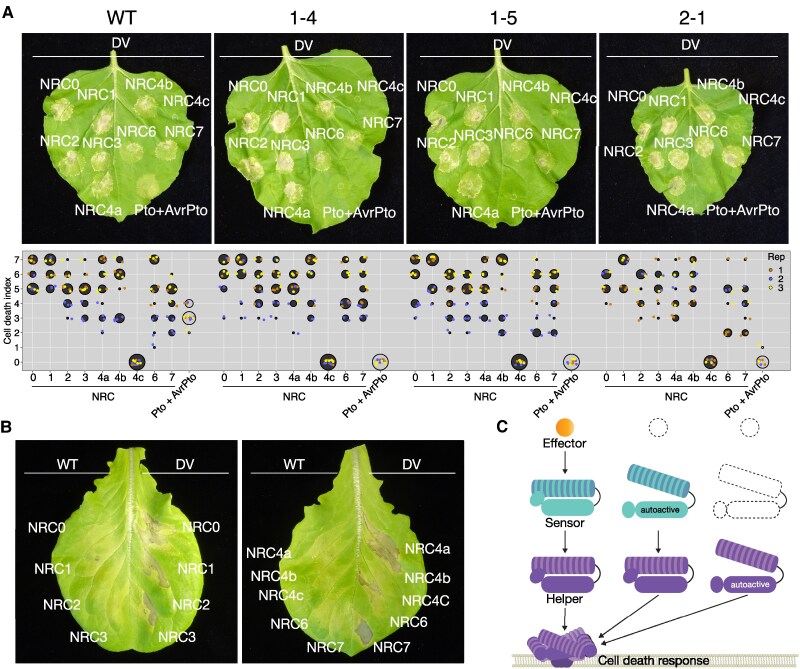 Figure 2
Figure 2- —Gatsby Charitable Foundation10.13039/501100000324
- —European Research Council10.13039/100010663
- —DFG Walter Benjamin Programme
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsToxin Mechanisms and Immunotoxins · Transgenic Plants and Applications · Plant-Microbe Interactions and Immunity
Dear Editor,
Nucleotide-binding domain and leucine-rich repeat immune receptors (NLRs) can function in networks of sensors and helpers to induce hypersensitive cell death and immunity against pathogens. The tomato (Solanum lycopersicum) sensor NLR Prf guards the Pto kinase from AvrPto and AvrPtoB effector perturbation and activates the downstream helpers NRC2 and NRC3 (Mucyn et al. 2006; Wu et al. 2017; Zhang et al. 2024). Prf is conserved across the Solanaceae and its ortholog in the model species Nicotiana benthamiana is also required for detection of AvrPto/AvrPtoB function on Pto (Lu et al. 2003; Mucyn et al. 2006). A recent study reported that cell death induction after transient expression of an autoactive mutant of tomato NRC3 is abolished upon RNAi silencing of Prf in N. benthamiana (Zhang et al. 2024). Here, we generated loss-of-function prf mutants in N. benthamiana and demonstrate that autoactive mutants of eight canonical tomato NRCs (NRC0, NRC1, NRC2, NRC3, NRC4a, NRC4b, NRC6, and NRC7) still induce hypersensitive cell death when expressed transiently in the prf mutant background. Autoactive tomato NRCs also triggered cell death when expressed in lettuce (Lactuca sativa), an Asteraceae plant that does not have a Prf ortholog (Pai et al. 2025). These results confirm a unidirectional dependency of sensors and helpers in the NRC network and underscore the value of the N. benthamiana and lettuce model systems for studying functional relationships between paired and networked NLRs.
Intracellular recognition of pathogen effector proteins by NLRs typically leads to induction of cell death (Jones and Dangl 2006). While singleton NLRs can detect effectors and induce cell death, paired and networked NLRs can be distinguished into phylogenetic and functional clades of sensor and helper NLRs (Contreras et al. 2023a). In the NRC (NLR required for cell death) network of asterid plants, sensors detect effectors and induce the oligomerization of downstream helpers. Activated helpers form membrane localized resistosomes, which act as calcium channels to induce cell death (Wu et al. 2017; Ahn et al. 2023; Contreras et al. 2023b; Liu et al. 2024; Madhuprakash et al. 2024). The activation of NRC helpers by sensors follows the activation-and-release model, in which sensors are not part of activated helper resistosomes (Ahn et al. 2023; Contreras et al. 2023b; Madhuprakash et al. 2024). The cell death function of helpers is mediated by the N-terminal coiled-coil domain, encoding the conserved MADA motif (Adachi et al. 2019), while sensors have degenerated N-termini and can encode an extended N-terminus of variable length prior to a solanaceous-domain (SD) (Mucyn et al. 2006; Seong et al. 2020), which is present in a subclade of NRC sensors (Contreras et al. 2023a).
NRC helpers are widely present across Solanaceae and can be grouped into 11 distinct phylogenetic sub-clades (Madhuprakash et al. 2024; Lüdke et al. 2025). Several disease resistance genes are NRC sensors which require NRCs from the helper clades for cell death induction and immunity. Previously studied NRC network components include the sensors Rpi-amr1a and Gpa2 which signal through NRC2/3, Rpi-amr1e, Rpi-amr3, Bs2, Rx, R1, Sw-5b, and R8 signaling through NRC2/3/4, as well as Rpi-blb2 and Mi-1.2 which exclusively signal through NRC4 (Wu et al. 2017; Witek et al. 2021; Lin et al. 2022, 2023). While the Hero resistance gene signals specifically through NRC6 (Lüdke et al. 2025), the pepper (Capsicum annuum) CaRpi-blb2 signals through NRC8/9 (Oh et al. 2023). The Pseudomonas syringae pv. tomato (Pst) resistance protein Prf from tomato (Solanum lycopersicum) contains an extended N-terminal domain prior to the SD-domain, which is required for interaction with the Pto kinase (Mucyn et al. 2006; Ntoukakis et al. 2014). The Prf/Pto complex binds and detects the Pst effectors AvrPto and AvrPtoB, leading to activation of NRC2 and NRC3 for cell death induction and immunity (Wu et al. 2004, 2016, 2017; Wu and Kamoun 2021; Sheikh et al. 2023; Zhang et al. 2024). A Prf ortholog is present in the model species N. benthamiana and is also required for detection of AvrPto/AvrPtoB function on Pto (Lu et al. 2003; Mucyn et al. 2006).
A recent study reported that cell death induction after transient expression of SlNRC3^H478AD479V^, an autoactive mutant of SlNRC3, is abolished upon RNAi silencing of Prf in N. benthamiana (Zhang et al. 2024). This work suggests that the cell death of autoactive NRC3 is genetically dependent on the Prf sensor, in sharp contrast with the previously reported unidirectional network architecture (Wu et al. 2017). Here, we revisited these experiments using mutant N. benthamiana plants rather than RNAi. We used CRISPR/Cas9 to generate three independent loss-of-function prf mutant lines to determine the extent to which autoactive mutants of helper NLRs of the NRC phylogenetic clade can induce cell death independently of Prf. In addition, we expressed autoactive helper NRCs in lettuce (Lactuca sativa var. Fenston), an Asteraceae plant that does not encode for a Prf homolog (Pai et al. 2025). Our results revealed that autoactive variants of eight tomato NRC helpers, including SlNRC3, can still trigger cell death in the absence of functional Prf. These findings confirm the unidirectional nature of the NRC sensor-helper networks of NLRs and demonstrate the utility of N. benthamiana genetic mutants and lettuce as a model system for dissecting sensor-helper interactions.
The N. benthamiana genome encodes two Prf copies, Prfa (Niben101Scf00650) and Prfb (Niben101Scf01984) (Kourelis et al. 2021; Supplementary Fig. S1). We used CRISPR/Cas9 to generate N. benthamiana lines with loss-of-function mutations in both copies, targeting the first 300 nucleotides, respectively (Fig. 1A). Based on AlphaFold3 predictions, these loci encode a coiled-coil domain structure within the extended N-terminal domain (Supplementary Fig. S2), which has been shown to be important for Pto interaction (Saur et al. 2015). While prf 1-4 contains an in-frame deletion in Prfa and Prfb, prf 1-5 and prf 2-1 contain frame shifts in Prfa and Prfb, respectively (Fig. 1A). To test the prf lines for AvrPto recognition, we performed Agrobacterium-mediated transient expression. While AvrPto, and Pto/AvrPto expression leads to cell death in wild type plants, no cell death was observed in prf mutant lines (Fig. 1B, Supplementary Fig. S3). Loss of cell death was complemented by co-expressing SlPrf with Pto/AvrPto, while neither Pto, nor SlPrf expression induced cell death (Fig. 1B, Supplementary Fig. S4). Co-expression of SlPrf with Pto induced a weak cell death response independent of AvrPto, as previously reported (Mucyn et al. 2006), which was strongly increased upon AvrPto co-expression (Supplementary Fig. S4) Co-expression of Rx with Potato virus X coat protein (CP), was used as cell death control induced by an NRC sensor-helper pair (Fig. 1B). These results indicate that all generated mutant lines, including the in-frame prf 1-4 mutant, are Prf loss-of-function mutants.
The tomato reference genome encodes a total of 11 proteins in the NRC phylogenetic helper clade (NRCX, NRC0, NRC1, NRC2, NRC3, NRC4a, NRC4b, NRC4c, NRC5, NRC6, and NRC7), all of which have canonical signatures of functional helper NLR proteins, except for NRC5, which does not encode an MHD motif (Lüdke et al. 2025) and NRCX, which acts as a modulator protein (Adachi et al. 2019). We tested if transient expression of the 9 canonical tomato NRC helpers as autoactive MHV mutant variants leads to the induction of a cell death response in the prf mutant lines. Expression of tomato NRC0, NRC1, NRC2, NRC3, NRC4a, NRC4b, NRC6, and NRC7 as autoactive MHV mutant variants induced a cell death response in the prf mutant lines similar to the wildtype plants (Fig. 2A). In contrast to results received by RNAi silencing of Prf in N. benthamiana, we could not detect a reduction of cell death after transient expression of autoactive versions of NRC helpers in prf mutant lines (Fig. 2A, Supplementary Fig. S5). In all experiments, the NRC4c autoactive MHV mutant variant did not induce a cell death response, which is in line with reports showing that NRC4c could not be activated by NRC4-dependent sensor NLRs (Huang et al. 2024; Lüdke et al. 2025). Next, we tested if autoactive tomato NRC mutant variants induce a cell death response when expressed transiently in lettuce (Lactuca sativa), which does not encode a Prf homolog (Pai et al. 2025). Similar to transient expression in N. benthamiana, tomato NRC0, NRC1, NRC2, NRC3, NRC4a, NRC4b, NRC6, and NRC7 induced a cell death response in lettuce when expressed as autoactive MHV mutant variants, but not as wildtype protein (Fig. 2B). We conclude that autoactive NRC helper proteins can function in the absence of functional sensor NLRs like Prf, consistent with the activation-and-release model (Ahn et al. 2023; Contreras et al. 2023b).
Several studies outlined that Prf signals through NRC2 and NRC3 for the induction of cell death and immunity. We showed that NRCs can induce cell death when expressed as autoactive mutants in the absence of functional Prf, either in N. benthamiana or in lettuce. Our results are consistent with the activation-and-release model (Ahn et al. 2023; Contreras et al. 2023b) and the observation that activated Prf sensors and NRC2/NRC3 helpers do not form a stable complex (Sheikh et al. 2023). In addition, NRC-dependent sensors are not part of NRC helper hexameric resistosomes (Contreras et al. 2023b; Madhuprakash et al. 2024). Our phylogenomic analyses are also consistent with the view that NRCs do not require Prf to function. Whereas NRC3 is present in all 35 examined Solanaceae genomes, Prf genes are missing in six of the 35 Solanaceae genomes, further suggesting that Prf is dispensable for NRC3 function (Supplementary Fig. S1). In addition, autoactive tomato NRC mutants can trigger cell death when expressed in lettuce, which does not carry a Prf or an NRC3 ortholog and diverged from tomato 97.5 to 109.2 mYA (Pai et al. 2025).
Altogether, these findings are consistent with the view that the edges in the NRC network of sensors and helpers have unidirectional relationships. While effector activated or autoactive sensors require a downstream helper for the induction of cell death, autoactive helpers can act independently of upstream sensors for the induction of a cell death response (Fig. 2C).
Supplementary Material
kiaf506_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adachi H, Contreras MP, Harant A, Wu C-H, Derevnina L, Sakai T, Duggan C, Moratto E, Bozkurt TO, Maqbool A, et al An N-terminal motif in NLR immune receptors is functionally conserved across distantly related plant species. Elife. 2019:8:e 49956. 10.7554/e Life.4995631774397 PMC 6944444 · doi ↗ · pubmed ↗
- 2Ahn H-K, Lin X, Olave-Achury AC, Derevnina L, Contreras MP, Kourelis J, Wu C-H, Kamoun S, Jones JDG. Effector-dependent activation and oligomerization of plant NRC class helper NL Rs by sensor NLR immune receptors Rpi-amr 3 and Rpi-amr 1. EMBO J. 2023:42(5):e 111484. 10.15252/embj.202211148436592032 PMC 9975942 · doi ↗ · pubmed ↗
- 3Contreras MP, Lüdke D, Pai H, Toghani A, Kamoun S. NLR receptors in plant immunity: making sense of the alphabet soup. EMBO Rep. 2023 a:24(10):e 57495. 10.15252/embr.20235749537602936 PMC 10561179 · doi ↗ · pubmed ↗
- 4Contreras MP, Pai H, Tumtas Y, Duggan C, Yuen ELH, Cruces AV, Kourelis J, Ahn H-K, Lee K-T, Wu C-H, et al Sensor NLR immune proteins activate oligomerization of their NRC helpers in response to plant pathogens. EMBO J. 2023 b:42(5):e 111519. 10.15252/embj.202211151936579501 PMC 9975940 · doi ↗ · pubmed ↗
- 5Huang C-Y, Huang Y-S, Sugihara Y, Wang H-Y, Huang L-T, Lopez-Agudelo JC, Chen Y-F, Lin K-Y, Chiang B-J, Toghani A, et al Subfunctionalization of NRC 3 altered the genetic structure of the Nicotiana NRC network. P Lo S Genet. 2024:20(9):e 1011402. 10.1371/journal.pgen.101140239264953 PMC 11421798 · doi ↗ · pubmed ↗
- 6Jones JDG, Dangl JL. The plant immune system. Nature. 2006:444(7117):323–329. 10.1038/nature 0528617108957 · doi ↗ · pubmed ↗
- 7Kourelis J, Sakai T, Adachi H, Kamoun S. Ref Plant NLR is a comprehensive collection of experimentally validated plant disease resistance proteins from the NLR family. P Lo S Biol. 2021:19(10):e 3001124. 10.1371/journal.pbio.300112434669691 PMC 8559963 · doi ↗ · pubmed ↗
- 8Lin X, Jia Y, Heal R, Prokchorchik M, Sindalovskaya M, Olave-Achury A, Makechemu M, Fairhead S, Noureen A, Heo J, et al Solanum americanum genome-assisted discovery of immune receptors that detect potato late blight pathogen effectors. Nat Genet. 2023:55(9):1579–1588. 10.1038/s 41588-023-01486-937640880 PMC 10484786 · doi ↗ · pubmed ↗