AmpC β-Lactamase-Producing Microorganisms in South American Hospitals: A Meta-Regression Analysis, Meta-Analysis, and Review of Prevalence
Valmir Nascimento Rastely-Junior, Hosanea Santos Nascimento Rocha, Mitermayer Galvão Reis

TL;DR
This study reviews the prevalence of AmpC β-lactamase-producing bacteria in South American hospitals and finds significant variation across species and study types.
Contribution
The study provides a comprehensive meta-analysis of AmpC β-lactamase prevalence in South American hospitals, identifying key factors influencing detection rates.
Findings
AmpC β-lactamases were detected in 11.7% of isolates across 48,801 samples.
Enterobacter species had the highest AmpC prevalence (~46%), while Escherichia spp. had the lowest (~4.5%).
Meta-regression showed higher prevalence in single-genus studies and lower prevalence in studies including pediatric patients.
Abstract
AmpC β-lactamases are class C enzymes that hydrolyze penicillins, cephalosporins, and monobactams. The WHO recently classified third-generation cephalosporin-resistant and carbapenem-resistant Enterobacterales as critical pathogens. We conducted a systematic review and meta-analysis to evaluate AmpC prevalence in hospital isolates across South America. We searched PubMed/MEDLINE, SciELO, and Google Scholar. We included 69 observational studies that phenotypically or genotypically identified AmpC producers. A random-effects generalized linear mixed model with logit transformation estimated pooled prevalence; heterogeneity and moderators were explored through subgroup analyses and meta-regression. Seventy studies, including 48,801 isolates, were eligible. AmpC β-lactamases were detected in 11.7% of isolates (95% CI 11.4–12.0), with extreme heterogeneity (I2 ≈ 97%). Enterobacter species…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
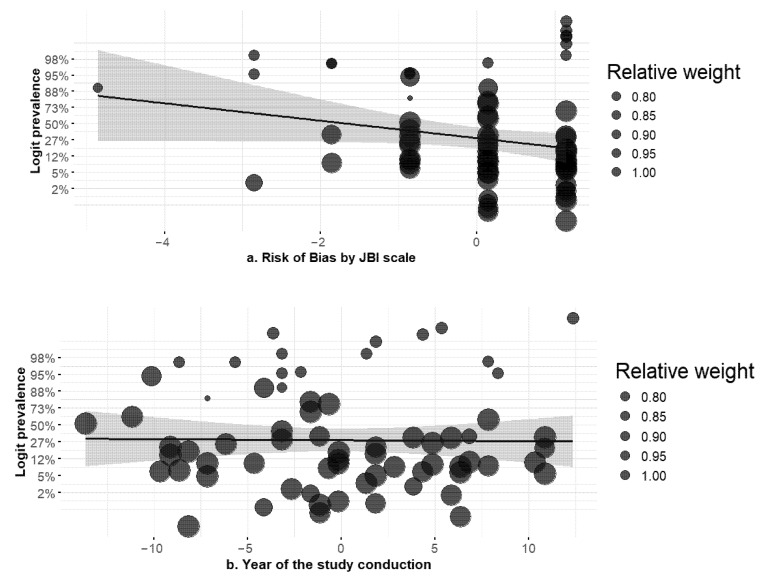 Figure 1
Figure 1 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Antibiotic Use and Resistance · Vibrio bacteria research studies
1. Introduction
Antimicrobial resistance (AMR) is one of the biggest threats to global public health in the 21st century. It undermines decades of medical progress and puts the foundations of modern medicine at risk [1]. There are many ways that bacteria have learned to avoid antimicrobial agents, but the production of β-lactamase enzymes by Gram-negative pathogens is one of the main reasons why they are resistant to the β-lactam class of antibiotics [2]. These enzymes work by hydrolytically breaking the amide bond in the four-atom β-lactam ring, which makes the antibiotic molecule inactive [3]. While considerable research and clinical focus have been directed towards specific classes of β-lactamases, notably extended-spectrum β-lactamases (ESBLs), other equally significant enzymes have been relatively underexplored in numerous areas. AmpC β-lactamases constitute a unique and particularly formidable resistance mechanism, characterized by their distinct biochemical properties, intricate genetic regulation, and the substantial diagnostic and therapeutic challenges they pose in clinical contexts. AmpC β-lactamases are very good at breaking down a lot of β-lactam agents, such as most penicillins, monobactams like aztreonam, and, most importantly, first-, second-, and third-generation cephalosporins like cefotaxime, ceftazidime, and ceftriaxone [4].
The global crisis of AMR is not evenly spread out. Its causes and effects are often made worse by social and economic factors, which puts a disproportionate burden on countries with low and middle incomes [1]. The rise of drug-resistant pathogens is especially harmful for South America, which has a wide range of healthcare systems and populations. To find out what public health actions are most important, we need to know the details of this regional crisis. AMR is seen as a big threat in the area, but the scientific and public health response has mostly been based on the dramatic and well-documented rise in ESBLs. This may have made it easier to miss other, less well-known resistance mechanisms like AmpC [4]. The wealth of detailed and fairly recent epidemiological data on ESBLs stands in stark contrast to the thin, fragmented, and mostly out-of-date information on AmpC β-lactamases.
This fragmented and outdated dataset is fundamentally inadequate for guiding evidence-based public health policy, clinical practice, and infection control across a continent as large and diverse as South America. It is impossible to accurately assess the true public health burden of AmpC-mediated resistance and justify or direct such resources effectively without a consolidated, contemporary overview. Policymakers cannot determine whether AmpC prevalence is a low-level, stable issue or a rapidly emerging crisis that is simply going unmeasured [5].
This uncertainty has direct consequences for the patient’s bedside. The development of effective empirical antibiotic prescribing guidelines relies on accurate and up-to-date local and regional antibiograms [6]. In the absence of data, particularly data addressing AmpC prevalence, clinicians managing patients with severe Gram-negative infections must make judgments based on insufficient knowledge. This may result in the improper utilization of third-generation cephalosporins or β-lactam/β-lactamase inhibitor combos, leading to therapeutic failure, heightened morbidity, and extended hospitalizations [7].
This systematic review seeks to systematically identify, critically evaluate, and synthesize the existing evidence about the prevalence of AmpC β-lactamases in clinical isolates from hospital environments throughout South America.
2. Materials and Methods
2.1. Register and Guidelines
This systematic review and meta-analysis followed PRISMA 2020 statement guidelines [8] and was prospectively registered in PROSPERO (CRD420251084285). The study commenced on 2 July 2025 and was completed on 12 August 2025.
2.2. Eligibility Criteria
Observational studies conducted in hospitals in South America that identified AmpC-type β-lactamase-producing bacteria by phenotypic and genotypic methods were included. Studies with patients of any age, including mixed populations (inpatient and outpatient), were accepted, provided the data were predominantly related to the hospital setting or allowed for an estimation of prevalence in this context. The outcome of interest was AmpC prevalence, including studies providing indirect data (total number of isolates and number of positive isolates). There were no language or time restrictions. We excluded non-original studies (such as reviews, editorials, or letters), environmental investigations, case reports, in vitro models, and exclusively outpatient studies.
2.3. Information Sources
Studies were searched without language or time restrictions in MEDLINE via Pubmed, SciELO, and Google Scholar. The strategy used a sensitive string combining South American countries, hospital terms, and AmpC synonyms, adapted to each database’s syntax. In Google Scholar, there was no limit on the number of results. The search was complemented by citation tracking, reference lists, gray literature (theses, conference proceedings), and contact with experts. All references were organized in Zotero and deduplicated in Rayyan AI before screening. We considered including LILACS, but it was excluded because it yielded no unique eligible studies beyond those found in the combination of final chosen databases.
2.4. Search Strategy
Construction of the search string—A sensitive string was developed combining South American countries, hospital-related terms, and AmpC synonyms. The search string used in PubMed was as follows:
(Argentina OR Bolivia OR Brazil OR Chile OR Colombia OR Ecuador OR Guyana OR Paraguay OR Peru OR Suriname OR Uruguay OR Venezuela OR “South America*” OR “South America”[Mesh]) AND (“critical care*” OR “critical care*”[mesh] OR “ICU” OR “intensive care*” OR “Intensive Care Units”[Mesh] OR “neurocritic*” OR “critically ill” OR “Critical Illness*” OR “Critical Illness”[mesh] OR “hospital*” OR “Hospital Units”[Mesh] OR “Hospitals”[Mesh] OR “Hospital Departments”[Mesh] OR “emergenc*” OR “Emergency Medical Services”[Mesh] OR “nosocomial”) AND (“AmpC*” OR “pAmpC*” OR “cAmpC*” OR “C β-lactamase” OR “C beta-lactamase” OR “ampicillinase Cs” OR “ampicillinase C”).
The search strategies used in other databases are included in Supplementary Materials.
2.5. Study Selection Process
All retrieved records were imported into Zotero and Rayyan, and duplicates were removed. Two reviewers then independently screened titles/abstracts and full texts. A third, independent screen was performed with Claude Sonnet 3.7, which classified records against pre-specified eligibility criteria using the labels include/exclude/uncertain. Disagreements about the three screening procedures were reviewed by two authors, and final inclusion decisions were made by human consensus.
2.6. Data Extraction
Initially, two reviewers independently extracted the data. Secondly, a third extraction was produced and compared with the first ones using assistance from Claude Sonnet 4.7. After that, all fields and any disagreements were reviewed by two authors and final decisions were made by human consensus, using a standardized spreadsheet with a data dictionary, piloted beforehand to calibrate interpretations. For each study, the following were recorded: identification (first author, year of publication, and study period), country, study design, definition and hospital setting, sample size, age range (mean or median and range), type of diagnostic test for AmpC and eligibility criteria applied, isolated bacterial species and genera, year of sampling, total number of microorganisms isolated, and total number of AmpC-positive isolates. We classified isolates as AmpC producers when the source study provided (a) phenotypic confirmation using recognized approaches—typically cefoxitin-based screening followed by inhibitor-based confirmation (e.g., boronic acid or cloxacillin), AmpC disk tests, or equivalent confirmatory assays—or (b) genotypic confirmation by PCR and/or sequencing of ampC determinants (e.g., blaCMY, blaDHA, blaFOX, blaACC, blaMOX, blaEBC/ACT/MIR). When both phenotypic and genotypic results were available, genotypic confirmation took precedence. This process minimized extraction errors and ensured independence between reviewers.
2.7. Risk-of-Bias Assessment
Risk of bias in the included studies was independently assessed by two reviewers using the “JBI Critical Appraisal Checklists for Prevalence Study” for each study design [8]. In each study, reviewers classified risk as low, moderate, or high; disagreements were discussed until consensus was reached.
2.8. Data Synthesis
All analyses were conducted in R version 4.3.2 (2023-10-31 ucrt) using the meta (v8.2-0), metafor (v4.8-0), mice (v3.17.0), clubSandwich (v0.6.1), ggplot2 (v3.5.2) and dplyr (v1.1.4) packages.
Prevalence estimates were pooled using a binomial–normal generalized linear mixed model (GLMM) fitted on the logit scale [9,10]. For each outcome (number of AmpC-positive isolates divided by total isolates), we used meta::metaprop() (R package meta, v8.2-0) with method = “GLMM” and summary measure sm = “PLOGIT” to fit a random-effects GLMM with a logistic link and maximum likelihood estimator for between-study variance τ^2^. Confidence intervals for the pooled proportion and τ^2^ were obtained via the Hartung–Knapp adjustment. The GLMM does not require continuity corrections; however, if the model failed to converge (e.g., due to sparse data or quasi-separation), we used a Freeman–Tukey double-arcsine transformation (sm = “PFT”) with restricted maximum likelihood (REML) for τ^2^ as a fallback [11]. Heterogeneity was quantified by the Q statistic, τ^2^, and I^2^. When at least ten studies were available, publication bias was assessed using funnel plots and Egger’s test. A sensitivity analysis excluded studies with 0% or 100% observed prevalence.
Univariable meta-regression models were fitted to explore heterogeneity. For each moderator (year of sampling, country, diagnostic test type, inclusion of outpatients, inclusion of children/adolescents, sample origin, focused bacterial group, and detection objective), a random-intercept GLMM was fitted using metafor::rma.glmm() (package metafor, v v4.8-0) with the measure set to PLO (logit of the proportion) and maximum likelihood estimation [11]. Categorical moderators were coded with the most common category as reference, and levels represented by <5 studies were collapsed to “Other”. The year variable was centered, and the risk-of-bias score was centered for interpretation. Model fit and the reduction in τ^2^ relative to the intercept-only model (pseudo-R^2^) were reported. For each model, we attempted several approximation methods (CM, CM.EL, CM.AL, UM.FS) and optimisers (BFGS, Nelder–Mead, nlminb) with 20 000 iterations and relative tolerance 10^−10^; if no GLMM converged, we fitted a REML random-effects model on the logit-transformed proportions with a continuity correction of 0.5 added to all cells. Robust standard errors were obtained via the HC3 method using clubSandwich [12,13].
For the multivariable meta-regression, we simultaneously evaluated three prespecified moderators: inclusion of children/adolescents (yes/no), the risk-of-bias score (centered), and whether the study focused on a specific bacterial group (specific vs. various/any). Because these covariates were missing for some studies, we used multiple imputation (mice, v3.17.0) under the missing-at-random assumption. The imputation dataset included the logit effect size (yi), its variance (vi), study identifiers, total isolates, total AmpC-positive isolates, and the three moderators. Missing values were imputed using logistic regression (logreg) or polytomous regression (polyreg) for categorical variables and predictive mean matching (pmm) for the continuous risk-of-bias score. We generated 20 imputed datasets (m = 20), each with 20 iterations (maxit = 20), and used the default predictor matrix. Each imputed dataset was analyzed with the GLMM described above; if a GLMM failed, the REML fallback was used. Regression coefficients and their standard errors were combined across imputations using Rubin’s rules via mice::pool(), and pooled confidence intervals were calculated. Between-study heterogeneity (τ^2^) and pseudo-R^2^ were averaged across imputations. Diagnostics included inspection of imputation distributions, fraction of missing information, influence analysis, sensitivity to exclusion of zero- and full-prevalence studies, and collinearity diagnostics (Pearson correlations, Cramér’s V, variance inflation factors, condition indices) [13,14]. A reproducible R code and all analysis scripts are available from the authors on request.
2.9. Certainty of Evidence Assessment (GRADE)
The certainty of the evidence was assessed using the GRADE approach [14], applied to the primary outcome of prevalence. Each pooled estimate was rated as high, moderate, low, or very low based on five domains: methodological limitations of the included studies, inconsistency across results, imprecision of estimates, indirectness relative to the review question, and potential publication bias.
3. Results
A systematic search across Medline, Scielo, and Google Scholar yielded over 15,000 records, but rigorous screening and duplication checks left 336 articles for full-text appraisal and 69 studies meeting the inclusion criteria, as shown in Figure S1. These studies collectively span nearly thirty years, indicating that AmpC β-lactamase producers have been present in South American hospitals since the late 1990s [15,16,17,18,19] and continue to be detected into the 2020s [20,21,22,23,24,25,26,27,28]. Surveillance intensified in the mid-2000s through sequential cross-sectional series and prospective cohorts, showing persistent circulation across institutions [29,30,31,32,33]. More recent point-prevalence studies and multicentric surveys from 2013 onward confirm that AmpC remains a contemporary challenge [21,34,35,36,37].
Table 1 shows that the evidence base includes a wide range of methods. Cross-sectional studies are predominant, indicating dependence on routine laboratory surveillance and facilitating prevalence comparisons across hospitals and time [29,38,39,40,41]. Although fewer in number, prospective and retrospective cohort studies elucidate temporal dynamics and correlations with clinical outcomes [16,21,25,42,43,44,45]. Molecular studies and outbreak investigations elucidate genetic mechanisms and transmission pathways [22,24,31,46,47]. Study durations varied from short outbreaks lasting a few weeks to extensive surveillance spanning nearly a decade [27,41,48,49], demonstrating that AmpC detection occurs regardless of follow-up duration.
Brazil and Colombia provide most of the data [39,45,60,64,65,68,69,70,71,77,81,84], with additions from large series from Argentina and Chile. Reports from Peru, Ecuador, Bolivia, Venezuela, and Uruguay also add to the data [31,54,59,75,82]. The settings examined are similarly diverse, encompassing adult and neonatal/pediatric ICUs and specialized units—such as oncology and transplant services—and extending to general wards and emergency departments. This variety shows that AmpC producers are not limited to high-risk units; they can be found throughout the hospital. While numerous studies concentrated on adults, a notable subset examined pediatric patients [56,63,72,76,83], especially in neonatal and pediatric ICUs, demonstrating that children are also impacted.
The majority of researchers evaluated clinical infections, corresponding with the clinical relevance of AmpC enzymes, whereas a lesser number examined both colonization and infection [29,35,53,61,62,66,74,78,80]. Only one study concentrated exclusively on colonization, underscoring a deficiency in the comprehension of asymptomatic reservoirs [43]. There were different ways to find things: phenotypic tests were the most common, but they were often used with genotypic tests to make them more accurate. The coexistence of multi-species and species-specific studies allowed for both broad and targeted insights. Stratification by species is important in meta-analyses. Most included articles were judged to be at low risk of bias using the JBI tool, indicating a generally robust evidence base, although a few studies exhibited moderate risk due to methodological limitations.
3.1. Meta-Analysis Results
The meta-analysis included 69 studies and 48,801 isolates of Gram-negative bacteria from South American hospitals. Using a random-effects GLMM with a logit transformation and maximum-likelihood estimator, the pooled prevalence of AmpC β-lactamase production across all microorganisms was 11.7% (95% CI 11.4–12.0%; τ^2^ = 0 on the logit scale; I^2^ = 97.2%; Q = 2429.55, p < 0.001; 95% prediction interval [PI] 11.4–12.0%), as shown in Table 2. Egger’s test showed no evidence of small-study bias (intercept ≈ 0; p = 0.779). The high I^2^ may reflect marked between-study and between-country variability (Figure S2). When studies reporting 0% or 100% prevalence were excluded, the pooled prevalence decreased modestly to 10.0% (95% CI 9.8–10.3%), with τ^2^ still 0 and I^2^ remaining high, indicating that heterogeneity was not driven solely by extreme findings.
Subgroup analyses by genus revealed substantial differences in intra-genus prevalences. Among Enterobacterales, Enterobacter spp. had the highest intra-genus pooled prevalence, at 46.0% (95% CI 43.7–48.4%; τ^2^ = 0; I^2^ = 30.9%; Q = 41.96, p = 0.057; PI 43.6–48.5%; k = 30). Although heterogeneity was moderate, Egger’s test indicated mild small-study effects (p = 0.037). Salmonella spp. (k = 7) showed a pooled prevalence of 28.5% (95% CI 21.2–37.0%; τ^2^ = 0; I^2^ = 0%; PI 19.6–39.3%); Egger’s test was not performed because of the small number of studies. Morganella spp. (k = 18) had a prevalence of 20.2% (95% CI 14.8–27.0%; τ^2^ = 0; I^2^ = 0%; PI 14.5–27.6%), with no evidence of small-study bias (p = 0.499). Serratia spp. (k = 21) showed 20.0% (95% CI 17.2–23.0%; τ^2^ = 0; I^2^ = 0%; PI 17.1–23.2%; Egger p = 0.852). Citrobacter spp. (k = 19) had a prevalence of 18.9% (95% CI 14.0–25.2%; τ^2^ = 0; I^2^ = 0%; PI 13.7–25.6%; Egger p = 0.086). Klebsiella spp. (k = 42) yielded 7.05% (95% CI 6.60–7.53%; τ^2^ = 0; I^2^ = 78.7%; Q = 192.15, p < 0.001; PI 6.6–7.6%), with significant small-study effects (p = 0.005). Proteus spp. (k = 27) showed 5.23% (95% CI 3.87–7.03%; τ^2^ = 0; I^2^ = 0%; PI 3.8–7.1%; Egger p = 0.584). Escherichia spp. (k = 39) had the lowest prevalence at 4.48% (95% CI 4.03–4.98%; τ^2^ = 0; I^2^ = 93.4%; Q = 575.38, p < 0.001; PI 4.0–5.0%), and Egger’s test suggested small-study bias (p = 0.028).
Among non-Enterobacterales, Acinetobacter spp. (k = 12) had a prevalence of 16.7% (95% CI 14.8–18.7%; τ^2^ = 0; I^2^ = 0%; PI 14.6–19.0%; Egger p = 0.046), while Pseudomonas spp. (k = 28) showed 19.3% (95% CI 18.0–20.7%; τ^2^ = 0; I^2^ = 83.4%; Q = 162.31, p < 0.001; PI 18.0–20.7%; Egger p = 0.249). Sensitivity analyses excluding 0%/100% studies yielded similar or modestly lower prevalence estimates for most genera, indicating that extreme proportions were not driving the pooled results.
3.2. Meta-Regression Results
The univariable meta-regressions evaluated each moderator separately (Table S1). Years of data collection showed no evidence of a temporal trend (Figure 1). Each additional year was associated with β = −0.03 (95% CI −0.14 to 0.08) and an odds ratio (OR) of 0.97 (95% CI 0.87–1.08); the p-value was 0.566 and the pseudo-R^2^ was 0%. This indicates that AmpC prevalence did not systematically increase or decrease over time, and nearly all variability remained unexplained.
As shown in Figure 2, country effects were suggestive but not statistically robust: compared with Brazil, studies from Argentina had β = −1.67 (95% CI −3.60 to 0.26), OR = 0.19 (95% CI 0.03–1.30), and p = 0.090; Chile showed β = 1.68 (95% CI −0.75 to 4.10), OR = 5.34 (95% CI 0.47–60.46), p = 0.176; Colombia had β = −0.70 (95% CI −2.55 to 1.15), OR = 0.50 (95% CI 0.08–3.14), p = 0.457. The omnibus test (QM = 8.989, p = 0.061) and pseudo-R^2^ of 6.6% suggest some geographic heterogeneity, but with wide confidence intervals and substantial residual heterogeneity (τ^2^ ≈ 75.4, I^2^ ≈ 99.3%).
The type of AmpC test did not account for variability. Compared with purely phenotypic testing, using both phenotypic and genotypic methods had β = 0.39 (95% CI −1.21 to 1.99), OR = 1.47 (95% CI 0.30–7.29), p = 0.634, and using genotypic methods alone had β = 1.07 (95% CI −0.70 to 2.83), OR = 2.90 (95% CI 0.50–17.01), p = 0.237. Including outpatients likewise had no significant effect (β = −0.98, 95% CI −2.81 to 0.86; OR = 0.38 [95% CI 0.06–2.36], p = 0.297). Methodological quality (risk of bias) showed a marginal association: each one-point increase in the JBI score corresponded to β = −0.53 (95% CI −1.10 to 0.04), OR = 0.59 (95% CI 0.33–1.04), and p = 0.070, with a pseudo-R^2^ of 4.1%. Bubble plots showed a slight downward slope. The sample origin (infection vs. colonization) showed no effect: comparing “both infection and colonization” versus infection had β = −0.25 (95% CI −2.13 to 1.64), OR = 0.78 (95% CI 0.12–5.14), p = 0.798, and comparing colonization alone versus infection had β = −0.17 (95% CI −6.08 to 5.73), OR = 0.84 (95% CI effectively undefined), p = 0.954.
Inclusion of children and adolescents was a statistically significant moderator. Studies that included pediatric populations had β = −1.77 (95% CI −3.31 to −0.22), OR = 0.17 (95% CI 0.04–0.80), and p = 0.025, explaining 9.7% of the heterogeneity; prevalence was roughly one-third of that in adult-only studies. Taxonomic focus emerged as a strong determinant: studies focused on a single bacterial genus or species had much higher prevalence than those screening a variety of genera. The contrast of “various/any” versus “specific” yielded β = −1.99 (95% CI −3.29 to −0.68), corresponding to OR = 0.14 (95% CI 0.04–0.51), p = 0.003, and a pseudo-R^2^ of 11.1%. This means that the odds of detecting an AmpC producer were reduced by ~86% when the study sampled multiple genera. The detection objective showed a borderline effect: when AmpC was a secondary rather than primary outcome, β = −1.42 (95% CI −3.00 to 0.16), OR = 0.24 (95% CI 0.05–1.17), p = 0.078, and pseudo-R^2^ = 2.3%.
In the multivariable meta-regression (Table S2), three moderators were entered simultaneously. Focusing on a variety of genera remained a strong independent predictor of lower prevalence, with β = −1.96 (95% CI −3.35 to −0.57), OR = 0.14 (95% CI 0.04–0.57), and p = 0.007. Inclusion of children/adolescents retained a large effect (β = −1.74, 95% CI −3.64 to 0.15; OR = 0.17 [95% CI 0.03–1.16]), but the p-value increased to 0.069, suggesting partial confounding with study focus. Study quality (risk of bias) was no longer significant (β = −0.04, 95% CI −0.70 to 0.62; OR = 0.96 [95% CI 0.50–1.85], p = 0.901). The multivariable model explained 19.5 % of between-study variance, yet residual heterogeneity remained extreme (τ^2^ ≈ 62.6, I^2^ ≈ 99.3%).
In summary, while most potential moderators—such as year of study, detection method, inclusion of outpatients, and sample origin—did not meaningfully explain the wide variation in AmpC β-lactamase prevalence across South American hospital studies, two factors consistently stood out. Studies targeting a single bacterial genus (often known high-risk groups) reported substantially higher prevalence than studies sampling multiple genera, and studies that included children or adolescents tended to find much lower prevalence. The findings indicate that variations in AmpC prevalence are primarily influenced by taxonomic emphasis and population age rather than by temporal trends, geographic disparities, or methodological factors, underscoring the necessity to meticulously evaluate study scope and demographics when comparing prevalence estimates.
4. Discussion
4.1. Principal Findings
A total of 69 studies encompassing 48,801 isolates were also eligible for meta-analysis. Using a random-effects generalized linear mixed model, we found that roughly one in ten isolates carried AmpC β-lactamase enzymes (pooled prevalence 11.7%, 95% CI 11.4–12.0%). The heterogeneity among studies was substantial (I^2^ = 97%), indicating significant variance in occurrence across different environments. Genus stratification indicated that AmpC producers were most prevalent on an intra-genus basis in Enterobacter spp. (~46%), moderate in Serratia spp. (~20%, almost no heterogeneity) and Pseudomonas spp. (~19%), appreciable in Salmonella spp. (~28%, few isolates), but much lower in Klebsiella spp. (~7%) and Escherichia spp. (~4.5%).
AmpC β-lactamase prevalence in South America appears elevated compared to most other regions. Europe and North America have some of the lowest documented AmpC rates globally (often only ~1–5% of Enterobacterales isolates) [85]. In contrast, parts of Asia and the Middle East show much greater AmpC burdens—certain local studies (for example, reports from India and Nepal) have observed AmpC prevalence rates exceeding 20–40% in clinical isolates. Africa’s experience seems more intermediate, with considerable variability by country; for instance, plasmid-mediated AmpC in E. coli has ranged from under 1% in some areas to nearly 15% in Egypt [86]. This places South America’s prevalence at an upper-middling level globally—lower than the highest rates seen in Asian hotspots, but significantly above the prevalence typically found in European or North American settings.
Addressing prevalence and geographic heterogeneity, the included studies indicate that AmpC β-lactamases are less frequent than ESBLs in South American hospital isolates, with global estimates for ESBLs typically at 10–40%. Clinically and phenotypically, AmpC differs from ESBLs by resistance to clavulanate and hydrolysis of cephamycins, and co-occurrence with ESBLs is reported—features that confound phenotypic algorithms and may inflate or obscure AmpC prevalence [87,88]. Regionally, the marked heterogeneity documented for ESBLs (e.g., shifting CTX-M variants) underscores the likelihood of locally variable AmpC epidemiology that current evidence cannot resolve [89]. Methodologically, underdetection is likely: widely used phenotypic screens have suboptimal sensitivity/specificity—especially for plasmid-mediated AmpC. CLSI lacks a standardized detection method. Multiplex PCR, while more reliable, is rarely routine, collectively biasing prevalence downward [4,87]. In parallel, South American hospitals face high burdens of ESBLs and carbapenemase producers, intensifying the clinical relevance of any unmeasured AmpC contribution and complicating interpretation of resistance phenotypes and empiric coverage decisions [90,91,92].
Finally, One Health linkages—documented across human, animal, and environmental interfaces—reinforce that hospital burdens are both recipients and amplifiers of broader transmission networks, further justifying coordinated interventions beyond the acute care setting [89].
4.2. Interpretation of Heterogeneity
Despite stratification and meta-regression, between-study variability remained very high. The taxonomic focus of studies consistently explained some heterogeneity: investigations targeting a single high-risk genus (e.g., Enterobacter spp.) reported far higher prevalence than those screening multiple genera. Multivariable meta-regression estimated that sampling a variety of genera reduced the odds of detecting an AmpC producer by roughly 86%. Population age also moderated prevalence: studies that included children and adolescents had markedly lower prevalence rates (odds ratio ≈ 0.17), possibly reflecting different exposure patterns, microbiota, or prescribing practices in pediatric wards. Other variables—such as year of conduction, country, detection method, inclusion of outpatients, and sample origin (infection vs. colonization cases)—did not significantly account for heterogeneity. The persistence of high between-study variance may reflect differences in species ecology and sampling frames, ward types (ICU vs. general), selective culturing, thresholds for screening, and case mix. Focusing on high-risk species can inflate prevalence, whereas broad screening across genera may dilute detection, underscoring the need for species-aware surveillance.
4.3. Implications for Policy and Surveillance
The high and heterogeneous burden of AmpC β-lactamases underscores the need for regional standardization of detection and reporting. South American health networks should develop unified phenotypic algorithms and build capacity for targeted molecular confirmation, ensuring that results are comparable across laboratories. Surveillance should integrate colonization and ambulatory settings to capture spill-over between hospitals and communities. Data sharing across countries will enable benchmarking of species-stratified prevalence and inform region-wide antibiograms. Public health authorities should incorporate AmpC reporting into national antimicrobial resistance plans, linking microbiology data with stewardship and infection control programs.
It is essential to recognize that the region possesses a robust policy framework and surveillance capacity for antimicrobial resistance—led by the PAHO/WHO and the ReLAVRA+ network. Yet hospital-level implementation of antimicrobial stewardship programs (ASPs) remains heterogeneous. Evidence syntheses report widespread adoption of national action plans and highlight ReLAVRA+ as the epidemiological backbone but also show that ASP maturity is concentrated in a few countries and that many institutions lack the infrastructure to monitor antimicrobial use and align prescribing with local microbiology [93,94,95]. Nevertheless, where well designed, ASPs are associated with lower antibiotic consumption and improved clinical outcomes, while strengthening pathogen detection and guiding de-escalation—strategies particularly relevant to reducing selective pressure from cephalosporins and carbapenems, implicated in the emergence of AmpC [96]. Recent advances include PAHO–GARDP partnerships to expand rational access to antimicrobials and build regional capacity, while feasible interventions such as multiprofessional education and prospective audit with feedback have demonstrated measurable impacts (e.g., shorter time to first dose in septic patients in Latin American ICUs) [97]. In sum, mitigating the burden of AmpC in the region requires translating intent into operational capacity—investing in multidisciplinary teams, monitoring technologies, and the continuous use of ReLAVRA+ data to inform local protocols and evaluate impact [98].
4.4. Clinical and Microbiological Implications
High-risk genera such as Enterobacter spp. account for nearly half of isolates tested, indicating that clinicians should remain vigilant when treating infections caused by these taxa. Species with intrinsic inducible ampC (Enterobacter cloacae complex, Citrobacter freundii, Klebsiella aerogenes) can initially appear susceptible to third-generation cephalosporins but become resistant after induction [4]. Clinically, misclassification of AmpC as ESBL risks inappropriate empiric therapy and failure given frequent multidrug resistance and cephamycin hydrolysis [88]. For inducible chromosomal AmpC in high-risk species, on-therapy derepression can raise β-lactam MICs and threaten clinical efficacy [99]. Quantitative data show substantial cefepime MIC shifts with derepression in Enterobacter cloacae complex [100]. Conversely, low-risk species (Serratia marcescens, Providencia spp., Morganella morganii) have a lower propensity for clinically significant derepression, permitting carbapenem-sparing regimens when susceptibility is confirmed [90]. Among alternatives, cefepime is supported by a meta-analysis of observational studies in bloodstream infection showing no significant mortality difference versus carbapenems; IDSA suggests cefepime for high-risk organisms when MIC ≤ 2 μg/mL. Important caveats include potential inoculum effect and diminished reliability with ESBL co-production [91,92,101]. Evidence for piperacillin–tazobactam is mixed and generally weaker, with lower reported susceptibility among AmpC producers [99]. For selected scenarios (e.g., uncomplicated urinary tract infection or step-down), non-β-lactam agents have observational support without increased mortality [102]. Finally, laboratory reporting strategies can materially alter carbapenem use, underscoring the need to pair species-aware advice with stewardship and reporting practices in a dedicated “Practice Implications” subsection [103]. Therapeutic considerations in this review are therefore framed as observational/indirect and should be interpreted as guidance rather than directives.
The rising antimicrobial resistance (AMR) among nosocomial Gram-negative pathogens increases the clinical significance of AmpC β-lactamase-producing Enterobacterales isolated from hospital settings in South America and highlights the necessity for strategies that transcend mere incremental adjustments to current medications [104]. Due to the limited pipeline and the prevalence of resistance mechanisms that compromise traditional β-lactams, strategies that leverage bacterial metal homeostasis should be prioritized [104,105]. The most important of these are “Trojan horse” siderophore–antibiotic conjugates (SACs), which steal iron transport to get around porin loss and efflux and deliver active drugs into the periplasm [104]. The siderophore–cephalosporin cefiderocol represents this paradigm; its catechol moiety facilitates active absorption, while its β-lactam core maintains efficacy against targets such as PBP3 and demonstrates stability against Class C β-lactamases pertinent to AmpC manufacturers [106,107,108,109]. However, resistance to the strategy itself is emerging via mutations or downregulation of siderophore receptors (e.g., cirA, fiu, piuA/piuD/pirA) and via co-expression of multiple β-lactamases with permeability defects, necessitating surveillance for receptor alterations alongside routine resistance testing [110]. Moreover, co-selection dynamics—where plasmids carry both siderophore and resistance determinants—suggest that iron-limited conditions selecting for metallophore systems may also enrich resistance traits, reinforcing the need for integrated stewardship and genomic surveillance [111].
Iron chelation can be used as an adjuvant to break up biofilms and make traditional antibiotics work better [112,113]. Metal mimics (e.g., gallium) can also be used to stop iron-dependent metabolism [114]. Inhibition of metallophore production pathways is another method [114,115,116,117]. Resistance will develop through alterations in siderophore–receptor interactions, efflux mechanisms, and co-/cross-resistance associated with metal exposure, necessitating monitoring and management that are sensitive to these processes [110,118].
4.5. Infection Prevention and Stewardship
The genus-specific risks observed in this review translate into actionable surveillance priorities. Hospitals should focus screening resources on ICUs, oncology/transplant wards and neonatal/pediatric units, particularly where Enterobacter or Serratia spp. predominate. Preventive strategies should bundle device stewardship and contact precautions during clusters and environmental hygiene audits with antimicrobial stewardship (AMS) protocols that flag likely AmpC-producing organisms. Regular review of AmpC trends integrated with antibiotic consumption metrics will help evaluate the impact of interventions and adjust empiric therapy. Because AmpC overexpression can be induced by third-generation cephalosporins [99], stewardship programs should promote carbapenem-sparing regimens (high-dose cefepime or β-lactam/β-lactamase inhibitor combinations) where appropriate, while reserving carbapenems for severe infections or high-inoculum states [99].
4.6. Limitations
The evidence is mainly observational and hospital-based, limiting generalizability to community settings. Methodological heterogeneity (assay types, breakpoints, sampling frames) and varied species mixtures may contribute to residual I^2^ despite subgrouping and meta-regression. In addition, most primary studies did not consistently distinguish chromosomally mediated, inducible AmpC from plasmid-mediated AmpC (pAmpC). Consequently, our prevalence estimates reflect combined AmpC mechanisms, which may contribute to heterogeneity and should be interpreted with caution when comparing across genera. Generalizability may be further constrained by geographic and genera imbalance: the dataset is dominated by Brazil and Colombia—the two most populous South American countries—and some genera (e.g., Salmonella spp.) were rarely studied, yielding imprecise estimates and limiting continent-wide inference. Because the review focuses on hospital isolates rather than community or general isolates, the organisms captured are more representative of hospital settings. Given the ecological nature of country-level subgrouping, we emphasize that unmeasured confounders such as hospital size and surveillance policies may influence prevalence estimates and cannot be fully accounted for in this meta-analysis. Long surveillance series may have included duplicate isolates, and colonization was under-represented since most studies focused on clinical infections. Although funnel and Egger tests showed no asymmetry, they may be underpowered under such extreme heterogeneity.
4.7. Certainty of Evidence (GRADE)
Using GRADE principles, we rated the certainty of the pooled prevalence as low to moderate. We downgraded for very serious inconsistency (I^2^ ≈ 97%) and indirectness. Imprecision affected less common genera. Risk of bias was generally low, but some studies had moderate risk due to incomplete reporting or convenience sampling. Publication bias was not detected, but tests were underpowered. Decisions based on these estimates should recognize that true prevalence may vary locally; however, the consistent detection of AmpC across decades and settings supports the conclusion that these enzymes are established in South American hospitals.
4.8. Research Priorities
Future studies should adopt prospective, specific designs with standardized diagnostic protocols and clear denominators to produce comparable prevalence estimates. Pediatric-focused research is needed to validate the lower prevalence observed in children and to assess neonatal risks. Time-series analyses could elucidate temporal trends and the impact of interventions such as pandemic-related changes. Evaluations of diagnostic algorithms and AMS bundles would provide evidence on effective strategies. Genomic epidemiology can differentiate plasmid-mediated versus chromosomal ampC, map transmission, and detect co-mechanisms with ESBLs or carbapenemases. Studies exploring colonization reservoirs and transition points (e.g., emergency departments, step-down units, outpatient clinics) will help identify entry points for intervention. Clinically, misclassification of AmpC as ESBL risks inappropriate empiric therapy and failure given frequent multidrug resistance and cephamycin hydrolysis [89]. Priorities are standardized surveillance that reports AmpC separately from ESBLs, combined phenotypic/genotypic detection, routine assessment of co-resistance, and country-level analyses to capture true geographic variation across South American hospital environments [74,89].
5. Conclusions
AmpC β-lactamase-producing organisms are entrenched in South American hospitals. Prevalence averages about one in ten isolates, but varies enormously across genera and settings, with Enterobacter spp. exhibiting the highest rates. Yet residual heterogeneity remained extreme; meta-regression showed that heterogeneity was not explained by calendar time or detection method but was influenced by taxonomic focus and population age, indicating that study design and case mix shape prevalence estimates. Implementing species-aware surveillance and stewardship programs is immediately actionable, while harmonized diagnostic methodologies and broader surveillance are needed to reduce uncertainty and monitor trends over time.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1WHO Antimicrobial Resistance 2023 Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance(accessed on 13 August 2025)
- 2Eiamphungporn W. Schaduangrat N. Malik A.A. Nantasenamat C. Tackling the Antibiotic Resistance Caused by Class A β-Lactamases through the Use of β-Lactamase Inhibitory Protein Int. J. Mol. Sci.201819222210.3390/ijms 1908222230061509 PMC 6121496 · doi ↗ · pubmed ↗
- 3Marsik F.J. Nambiar S. Review of Carbapenemases and Amp C-beta Lactamases Pediatr. Infect. Dis. J.2011301094109510.1097/INF.0b 013e 31823 c 0e 4722105420 · doi ↗ · pubmed ↗
- 4Jacoby G.A. Amp C β-Lactamases Clin. Microbiol. Rev.20092216118210.1128/CMR.00036-0819136439 PMC 2620637 · doi ↗ · pubmed ↗
- 5Ghafourian S. Sadeghifard N. Soheili S. Sekawi Z. Extended Spectrum Beta-lactamases: Definition, Classification and Epidemiology Curr. Issues Mol. Biol.201517112110.21775/cimb.017.01124821872 · doi ↗ · pubmed ↗
- 6Philippon A. Arlet G. Jacoby G.A. Plasmid-Determined Amp C-Type β-Lactamases Antimicrob. Agents Chemother.20024611110.1128/AAC.46.1.1-11.200211751104 PMC 126993 · doi ↗ · pubmed ↗
- 7Hanson N.D. Amp C-lactamases: What do we need to know for the future?J. Antimicrob. Chemother.2003522410.1093/jac/dkg 28412775673 · doi ↗ · pubmed ↗
- 8Munn Z. Moola S. Riitano D. Lisy K. The development of a critical appraisal tool for use in systematic reviews addressing questions of prevalence Int. J. Health Policy Manag.2014312312810.15171/ijhpm.2014.7125197676 PMC 4154549 · doi ↗ · pubmed ↗