Genome-Wide Identification and Analysis of the Fatty Acid Export Family Revealed the Role of GmFAX8 in Improving Soybean Oil Accumulation
Yan Zhang, Yina Zhu, Xiuli Rui, Yuan Li, Jie Wang, Yuhang Zhan, Yongguang Li, Xue Zhao, Yingpeng Han, Xunchao Zhao

TL;DR
This study identifies and analyzes fatty acid export genes in soybeans, finding that GmFAX8 boosts oil accumulation.
Contribution
The study identifies eight GmFAX genes in soybean and demonstrates that GmFAX8 enhances fatty acid accumulation.
Findings
Eight GmFAX genes were identified in soybean, grouped into four subfamilies.
GmFAX8 expression is high in leaves and stems, and overexpression increases fatty acid content in hairy roots.
GmFAX genes contain a conserved Tmemb_14 domain, suggesting functional importance.
Abstract
Fatty acid transporters (FAXs) play an important role in fatty acid synthesis by facilitating transport fatty acids from the plastid to the endoplasmic reticulum. This process is essential for providing precursor substances necessary for triglycerides (TAGs). Although FAX genes have been identified in variety of plant species, the identification and molecular functions of the GmFAX gene members in soybean are still unclear. In this study, soybean FAX genes were identified through the utilization of the Phytozome (v13) and NCBI online websites. Subsequently, phylogenetic trees, expression patterns, gene structures, and qRT-PCR were analyzed. A total of eight GmFAX members were identified at the whole genome level, and further phylogenetic analysis revealed that these members can be categorized into four subfamilies. In addition, all members of GmFAX contain a highly conserved domain…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
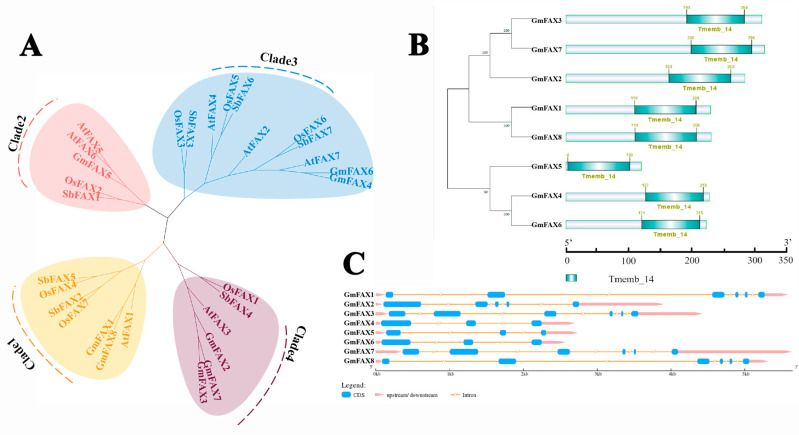 Figure 1
Figure 1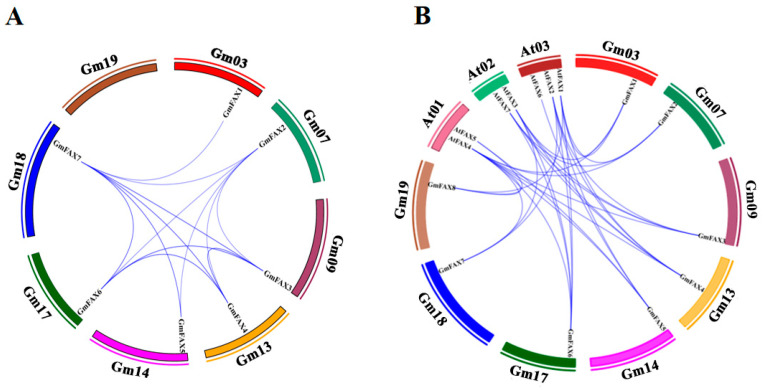 Figure 2
Figure 2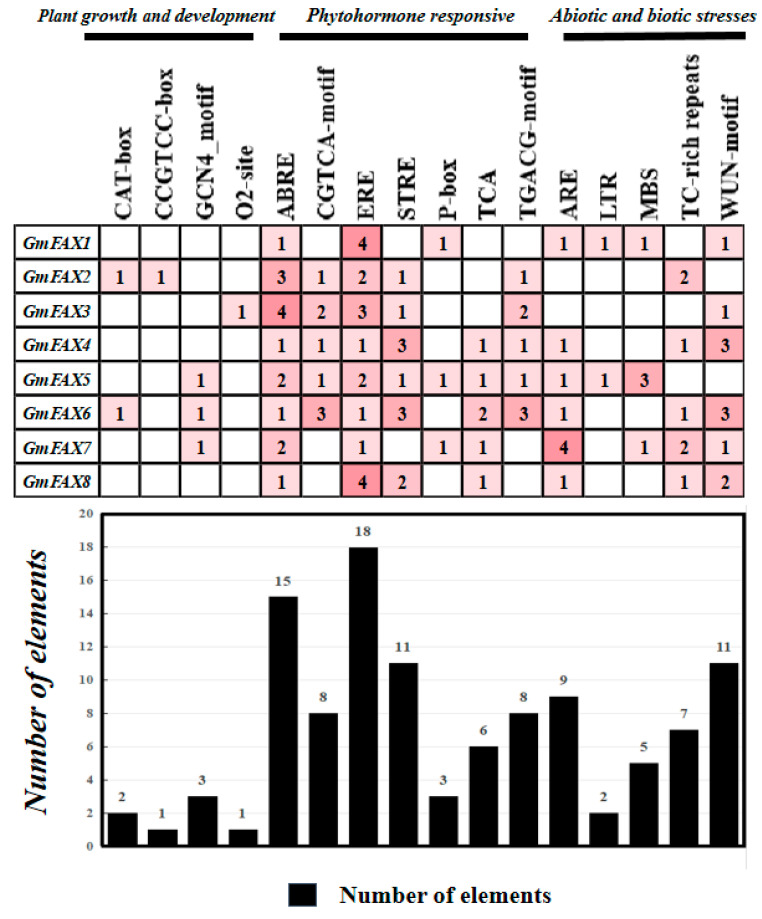 Figure 3
Figure 3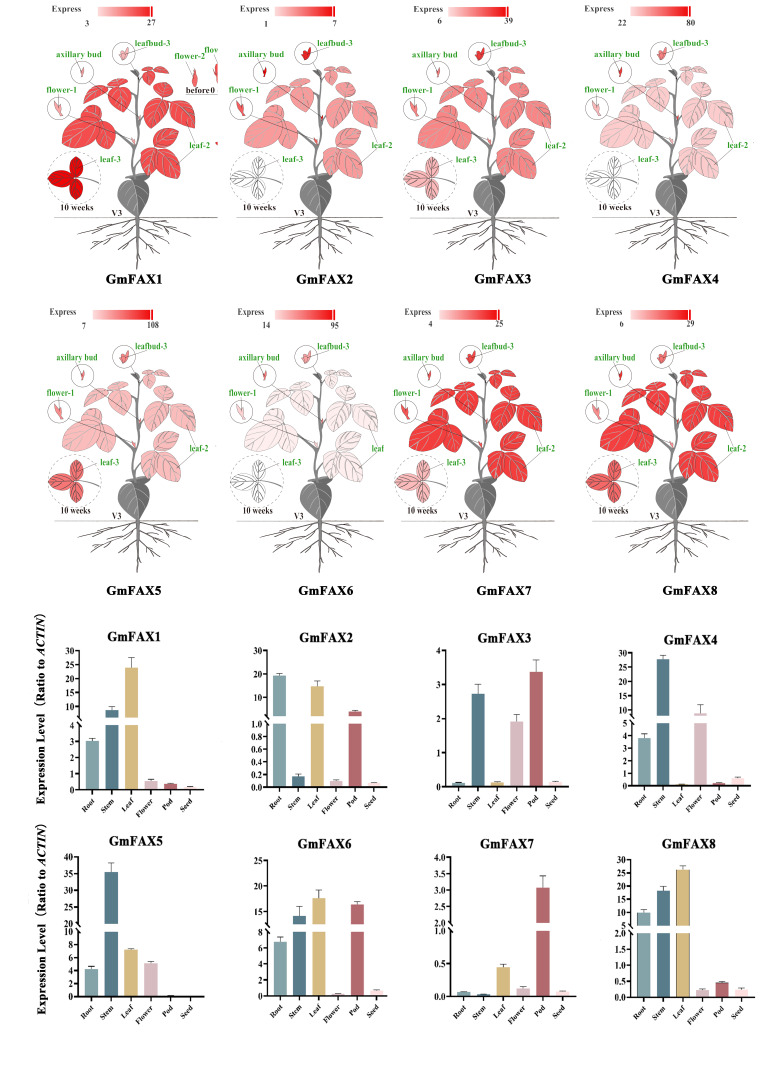 Figure 4
Figure 4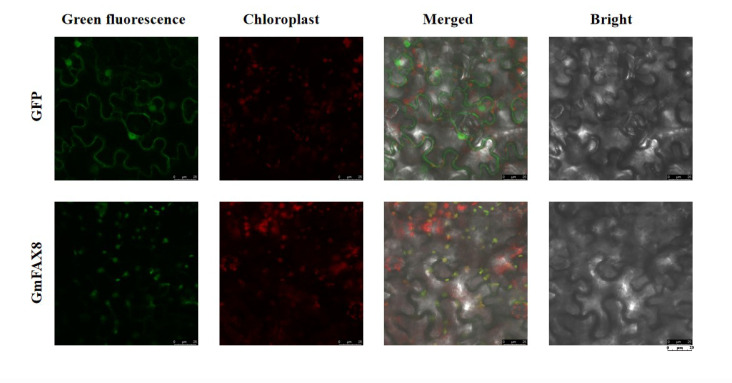 Figure 5
Figure 5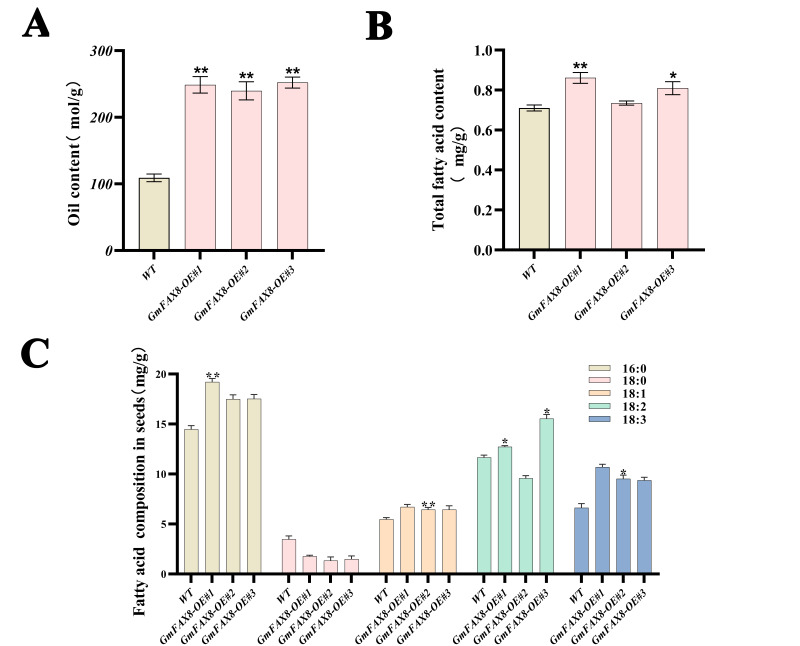 Figure 6
Figure 6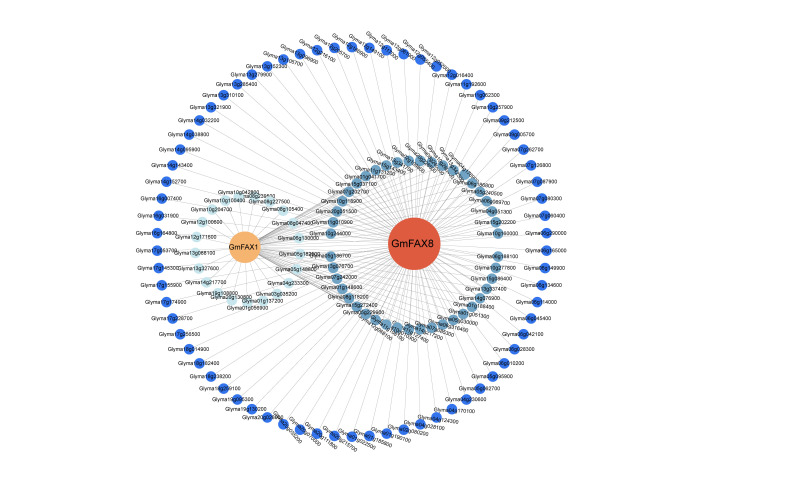 Figure 7
Figure 7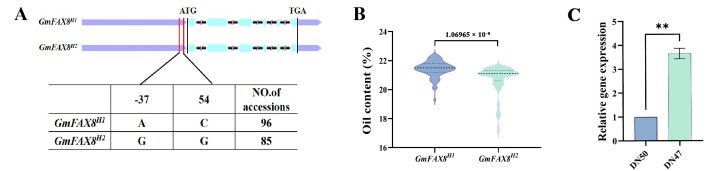 Figure 8
Figure 8- —The Chinese National Natural Science Foundation
- —Heilongjiang Provincial ‘Outstanding Young Teachers’ Basic Research Support Program
- —The Talent Introduction Project of Northeast Agricultural University
- —The National Key Research and Development Project of China
- —The Youth Leading Talent Project, The Ministry of Science and Technology in China
- —The national project
- —Young leading talents of Northeast Agricultural University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLipid metabolism and biosynthesis · Photosynthetic Processes and Mechanisms · Ubiquitin and proteasome pathways
1. Introduction
Fatty acids (FAs) are fundamental components of living organisms and are widely distributed in plants [1,2]. Fatty acids play a crucial role in the synthesis of triacylglycerols (TAGs), and serve as the primary component in oil biosynthesis [3,4]. The FA synthesis pathway represents one of the most essential metabolic pathways in organisms, characterized by its relative complexity [5]. The synthesis of fatty acids predominantly relies on acetyl-CoA as a substrate, which is derived from pyruvate produced through the glycolysis pathway from glucose [6]. Acetyl-CoA is condensed with malonyl-CoA to generate precursors for fatty acid synthesis. These precursors undergo a series of reduction, dehydration, and further reduction processes, ultimately resulting in the synthesis of long-chain fatty acids [5]. Then fatty acids are transported to their respective tissue sites via various transporters [7].
FAX serves as the initial transporter protein in the FA shuttle, and plays an important role in lipid synthesis [8]. The fatty acid transporter (FAX) gene is crucial for the regulation of lipid metabolism in plants, and it is widely present in animal and plant cells [8,9]. FAX genes play a crucial role in the transport of fatty acids from plastids to the endoplasmic reticulum, thereby supplying precursors necessary for triglyceride synthesis [10]. Previous studies have shown that FAX genes are crucial in the regulation of plant growth and development, as well as in the processes of oil synthesis [11]. So far, the FAX genes of many plant species have been cloned and characterized, including Arabidopsis, rapeseed, and green algae. In Arabidopsis, the loss of FAX1 function disrupts intracellular fatty acid/lipid homeostasis, further damaging the formation of pollen walls and pollen viability, leading to male sterility [12]. Similarly, double mutants of FAX2 and FAX4 exhibit reduced FA export compared to the wild type, resulting in a significant decrease in TAG content in the seeds [13]. In green algae, CrFAX1 and CrFAX5 participate in TAG synthesis by functioning in chloroplasts and endoplasmic reticulum membranes, respectively. CrFAX1 and CrFAX5 synergistically shuttle FA from chloroplasts to endoplasmic reticulum for TAG synthesis [14]. The overexpression of the FAX1-1 gene can significantly increase seed oil content in Brassica napus, and it was further found that the levels of PC and diacylglycerol (DAG) were significantly increased [15].
Soybean (Glycine max) is a major global oil crop, and increasing its seed oil content is a key objective for genetic improvement. Therefore, identifying key genes regulating soybean oil accumulation is crucial for developing high-oil varieties. Although FAX genes have been characterized in many plant species, the FAX gene family in soybean remains uncharacterized, and its role in regulating oil accumulation is unclear. In this study, eight FAX genes were identified in soybean, and their phylogenetic, gene structure, collinearity, and promoter elements were analyzed. In addition, it has also been preliminarily confirmed that the GmFAX8 gene plays a role in enhancing oil accumulation in soybeans.
2. Results
2.1. Identification of GmFAX Genes in Soybean
To identify the FAX gene family members in soybean, the protein sequence of Arabidopsis FAXs was used as queries to perform a BLASTP search against the soybean genome in the Phytozome (v13) database. A total of eight GmFAX genes were identified and named FAX–FAX8 based on their chromosomal positions. The full-length CDS sequence of GmFAX1–GmFAX8 ranged from 360 bp to 951 bp. The predicted isoelectric point of GmFAX genes ranged from 5.68 to 9.8, and their molecular weight ranged from 12.57 to 34.57 kDa. In order to further understand the phylogenetic relationship of the soybean FAX gene family, the FAX proteins of Arabidopsis, Glycine max, Oryza sativa, and Sorghum bicolor were obtained from the Phytozome (v13), and the phylogenetic tree was generated through MAGE version 11.0 software. The phylogenetic tree revealed that the FAX genes are divided into four distinct subfamilies (I–IV). Subfamily I includes GmFAX1 and GmFAX8; subfamily II contains GmFAX5; subfamily III comprises GmFAX4 and GmFAX6; and subfamily IV consists of GmFAX2, GmFAX3, and GmFAX7. To explore the gene structure of FAX genes, the gene structure of GmFAX genes was generated through GSDS online website. As shown in Figure 1, members of subfamily I typically possess six exons, whereas genes in subfamilies II and III contain only three exons. Conserved domain analysis performed with the SMART database revealed that all GmFAX proteins contain the Tmemb_14 domain, suggesting that their functions are evolutionarily conserved (Figure 1).
2.2. Collinearity Analysis of GmFAXs
To further characterize duplication events within the soybean genome, a synteny analysis of GmFAX genes was performed. The eight GmFAX genes were located on eight different chromosomes (Chr03, 07, 09, 13, 14, 17, 18, and 19). The synteny analysis identified 20 duplicated gene pairs among these GmFAX genes (Figure 2). A total of 20 pairs of syntenic gene pairs were identified in soybean. Furthermore, to trace the evolutionary history of these genes, we conducted a comparative syntenic analysis between soybean and Arabidopsis thaliana. As shown in Figure 2, there are duplicate events between GmFAXs and AtFAXs, including 30 pairs of reciprocal relationships between G. max and A. thaliana.
2.3. Analysis of Cis-Acting Elements of GmFAX Promoter
To further explore the cis-acting elements of the GmFAX genes promoter, the 2 kb genomic sequence upstream of the translational start site (ATG) of each GmFAX gene was analyzed. As shown in Figure 3, ABRE (abscisic acid-responsive element) and ERE elements were found in all GmFAXs. CAT-box (involved in meristem expression) elements were found in GmFAX2 and GmFAX6. The CCGTCC box element was only found in GmFAX2. Notably, an LTR (low temperature responsiveness) element was identified in GmFAX1 and GmFAX5, while an MBS (drought inducibility) element was found in GmFAX1, GmFAX5, and GmFAX7. The O2 site element was only found in GmFAX3 (Table S4). The prevalence of these stress- and hormone-related cis-elements strongly suggests that GmFAX genes play important roles in mediating plant growth, development, and responses to environmental stimuli.
2.4. Expression Pattern of Soybean FAXs in Different Developmental Stages
In order to determine the expression pattern of soybean FAX genes at different developmental stages, the high-throughput sequencing data of the National Bioinformatics Center (https://ngdc.cncb.ac.cn/soyomics/transcriptome/) (accessed on 27 March 2025) were retrieved and analyzed. As shown in Figure 4, the expression of GmFAX genes was investigated in different tissues. GmFAX4 was highly expressed in leaves, seeds, and root nodules, while GmFAX5 and GmFAX8 showed broad expression patterns across multiple tissues. At the same time, it was found that the expression levels of GmFAX5 and GmFAX8 genes were the highest in the flowers. In contrast, GmFAX2 displayed universally low expression levels across all examined tissues. Subsequently, through qRT-PCR, we found that the expression level of GmFAX2 was relatively low in different tissues, while the expression levels of FAX1 and FAX8 genes were relatively high in the roots, stems, and leaves. The expression levels of FAX4 and FAX5 were relatively high in the stems (Figure 4).
2.5. Subcellular Localization of GmFAX8
The GmFAX8 gene was amplified from the DN50 cDNA, and then it was constructed into the pCAMBIA3300 vector containing green fluorescent protein (GFP) and transformed into GV3101 (pSoup-p19) competent cells. The recombinant 35S::GmFAX8-GFP plasmid, along with the positive control (35S::GFP) was transferred into tender Nicotiana tabacum using the Agrobacterium mediated method. As shown in Figure 5, the tobacco leaves injected with the pCAMBIA3300-GmFAX8-GFP expression vector showed only visible green fluorescence at the chloroplasts when observed under a laser confocal microscope. No nuclear or cell membrane expression was found. The empty pCAMBIA3300-GFP could be observed to have green fluorescence at the cell membrane, cytoplasm, and nucleus, indicating that the GmFAX8 protein is mainly localized in the chloroplasts.
2.6. Overexpression of GmFAX8 Increased Total Fatty Acid Content
To investigate the role of GmFAX8 in total fatty acid accumulation, its coding sequence was cloned into the pYES3 vector, and the recombinant vector was transformed into Saccharomyces cerevisiae strain INVSc1. The oil content in yeast cells expressing the pYES3-GmFAX8 construct was significantly higher than in those harboring the empty pYES3 vector, which indicated that the GmFAX8 gene promotes the accumulation of oil content. Consistent with the yeast results, the total fatty acid content in transgenic GmFAX8 hairy roots was also significantly higher than in control lines (Figure 6), as determined by gas chromatography. Subsequently, when analyzing the contents of each component of fatty acids, we found that the contents of palmitic acid, oleic acid, linoleic acid, and linolenic acid were significantly higher than those of the control group. However, the content of stearic acid was significantly lower than that of the control group.
2.7. Co-Expression Analysis of Transcription Factors and GmFAXs in Soybean
In order to further clarify the regulatory mechanism of upstream transcription factors and GmFAX family genes at the transcription level, we constructed a co-expression network with transcription factors and GmFAX genes. As shown in Figure 7, a total of 179 co-expression modules were constructed, encompassing GmFAX1, GmFAX8, and 136 transcription factors. GmFAX8 showed a significant positive correlation with Glyma13g341700, Glyma11g131200, Glyma10g160000, Glyma06g042100, Glyma15g086400, and Glyma18g014900 (r > 0.90, p < 0.05). GmFAX1 showed a significant positive correlation with Glyma10g160000 (r > 0.97, p < 0.05). It is noteworthy that both GmFAX8 and GmFAX1 showed a significant positive correlation with the Glyma10g160000 gene (r > 0.90, p < 0.05). The above results suggest that the GmFAX8 gene may interact with multiple transcription factors to further regulate the accumulation of lipids.
2.8. Haplotype Analysis of the GmFAX8 Gene
In order to investigate the relationship between the GmFAX8 gene and the oil content of soybeans, we conducted a gene-based association analysis using the GLM model. In 181 soybean samples, two SNPs significantly related to the oil content were identified, namely rs39595759 and 39,595,813 (−log10 (P) ≥ 2.5) [10], which were located in the 5′ UTR and upstream regions of the GmFAX8 gene, respectively (Figure 8A). The result analysis revealed that the oil content of haplotype 1 was significantly higher than that of haplotype 2 (Figure 8B). Further analysis revealed that the expression level of the GmFAX8 gene in high-oil materials was significantly higher than that in low-oil materials (Figure 8C).
3. Discussion
FAX genes play an important role in the FA pathway and are mainly involved in the transmembrane transport of fatty acids [16]. Previous studies have shown that FAXs play an important role in the accumulation of seed oil across various species [8,16]. Although FAX genes have been analyzed in various plant species, including Arabidopsis [13], Brassica napus [17], Camelina sativa [18], and Green microalga [14], their identification and molecular functions in soybean remain unclear. It has been reported that FAX plays a crucial role in the transport of fatty acids, but the transport of fatty acids in soybeans still needs further exploration. In this study, eight GmFAX genes were identified in the soybean genome. Phylogenetic analysis can clearly prove the evolutionary relationship between soybean FAX and other species. The results showed that the eight GmFAXs were categorized into four distinct clusters according to their different evolutionary relationships and functional characteristics. All GmFAXs contained Tmemb_14 conserved domains. These results indicate that GmFAX genes were relatively conserved. Previous studies have identified seven AtFAX genes in Arabidopsis thaliana, named AtFAX1–7, which were divided into seven sub-clusters and performed different functions [8]. For instance, AtFAX1 (subfamily I) is localized to the chloroplast inner membrane of chloroplast and mediates the output of fatty acids from chloroplast [15]. Subfamilies II and IV, AtFAX2 and AtFAX4, are able to transport fatty acids into the endoplasmic reticulum, where they can be further converted into triacylglycerols [13]. Currently, the function of the FAX genes has been found in various species, including Arabidopsis, rapeseed, and green algae [16,17]. It was found that GmFAX genes are expressed in different tissues through the online website of the National Biological Information Center. The expression levels of GmFAX4, GmFAX5, and GmFAX6 were higher in different tissues, while the expression levels of GmFAX2 were lower in different tissues. Subsequently, the expression level of GmFAXs in different tissues was analyzed via qRT-PCR. It was found that the expression level of GmFAXs in stems and leaves was relatively high. Previous studies have found that AtFAX2 and AtFAX4 are highly expressed in the roots, stems, and leaves of Arabidopsis thaliana [13]. In rapeseed, FAX1 also has a relatively high expression level at different stages of tissue development. [17]. Phylogenetic tree and domain analysis indicated that FAX1 and FAX8 had high homology. Further co-expression network analysis revealed that transcription factors were positively correlated with FAX1 and FAX8, respectively. The above results indicate that the FAX1 gene may have a positive effect on lipid accumulation. To further elucidate the molecular function of GmFAX8 in lipid accumulation, transgenic soybean hairy roots were generated. The results showed that the total fatty acid content in GmFAX8-overexpressing hairy roots was significantly higher than in the control lines. Previous studies have found that the overexpression of BnaA09-FAX1 in Brassica napus can significantly increase the content of PC and DAG [17]. In Chlamydomonas reinhardtii, the co-expression of FAX1 and FAX2 significantly increased the content of triacylglycerol (TAG) [19,20,21]. In rapeseed, the overexpression of BnaFAX1-1 significantly increased the content of oleic acid (C18:1) in seeds [15]. Previous studies have found that the bHLH7 gene in Arabidopsis thaliana can significantly increase the oil content of seeds [22]. In this study, through the co-expression network, bHLH (Glyma13g341700) was identified to have a significant positive correlation with GmFAX8, which suggests that this gene plays an important role in regulating lipid accumulation. Through candidate gene association analysis, this study identified two haplotypes. Among them, haplotype 1 had a significantly higher oil content than haplotype 2. The above results can provide a basis for molecular-assisted selection breeding.
In conclusion, a total of eight soybean FAX genes were classified into four distinct clusters. These GmFAX genes exhibited varying expression levels at different developmental stages, and it was preliminarily demonstrated that the GmFAX8 gene can regulate the accumulation of soybean oil.
4. Materials and Methods
4.1. Identification of the FAX Gene Family
In order to screen the members of the FAX gene family in soybean, the reported protein sequences of Arabidopsis FAX family members were retrieved from the TAIR and used as alignment sequences to perform BLASTP sequence alignment with the soybean genome database on the online website of Phytozome (v13) (https://phytozome-next.jgi.doe.gov/). Candidate genes were selected based on an E-value < 10^−10^ and sequence similarity >90% [23]. The presence of the characteristic FAX domain in these candidates was further verified using the SMART and Pfam databases [24,25]. The candidate genes of the soybean FAX gene family were determined, and their domain distribution map was drawn using IBS 1.0 [26]. The physicochemical properties of soybean GmFAX family members, including molecular formula, protein molecular weight, and isoelectric point, were predicted through the bioinformatics resource website ExPASy (http://www.expasy.org) [27].
4.2. Phylogeny, Genetic Structure, and Collinearity Analysis of the FAX Gene Family
The FAX protein sequences of soybean (Glycine max), Arabidopsis thaliana (A. thaliana), rice (O. sativa), and sorghum (Sorghum bicolor) were used to construct a phylogenetic tree based on the neighbor joining method and bootstrap test set at 1000 replicates using MEGA11 [28]. The intron/exon position structure of the FAX genes was obtained through the GSDS (https://gsds.gao-lab.org/Gsds_help.php) (accessed on 17 March 2025) online website [29]. To investigate the evolutionary relationships between the FAX genes of soybean and Arabidopsis, a collinearity analysis was performed using TBtools software [30].
4.3. Promoter Analysis of GmFAXs
In order to explore the key cis-acting elements in the upstream promoter region of GmFAX genes, the 2 kb upstream promoter sequence of GmFAX genes were obtained from Phytozome v13 database (https://phytozome-next.jgi.doe.gov/), and submitted to the PlantCARE database to analyze the category, quantity, and location of the cis-acting elements [23].
4.4. Expression Analysis of GmFAX During Soybean Development
The expression levels of the GmFAX gene in soybean at different developmental stages were obtained through the National Bioinformatics Center database (https://ngdc.cncb.ac.cn/soyomics/index)(accessed on 25 March 2025). To further confirm the expression levels of the GmFAX gene during different developmental stages, soybean seeds (DN50) were used as materials, and three plants with consistent growth were selected to collect samples at different developmental stages, including roots, stems, leaves, flowers, pods, and seeds. The collected samples were rapidly frozen in liquid nitrogen, and then RNA was extracted from each sample using Trizol reagent (Takara, Beijing, China). Quantitative real-time qRT-PCR was performed using the CFX Connect TM real-time system (Bio-RAD, Hercules, CA, USA) and SYBR Select Master Mix qRT-PCR (SYBR Green, Toyobo, Osaka, Japan). GmActin4 (GenBank accession number AF049106) was used as the internal reference. The expression level of GmFAX genes was calculated by 2^−ΔΔCt^ [31]. Each reaction was set up with three biological replicates and three technical replicates.
4.5. Subcellular Localization Analysis of GmFAX8
Using the cDNA of soybean DN50 as the template, the CDS region of GmFAX8 gene was amplified and cloned into the pCAMBIA3300 vector, which harbored a green fluorescent protein (GFP) tag to generate the pCAMBIA3300-GmFAX8::GFP construct to generate pCAMBIA3300-GmFAX8::GFP. The recombinant plasmid pCAMBIA3300-GmFAX8::GFP and the empty plasmid pCAMBIA3300 were transformed into Agrobacterium GV3101 (pSoup-p19), and the young leaves of tobacco were transiently infected by Agrobacterium. The empty vector pCAMBIA3300 (containing green fluorescent GFP label) was used as the control. The expression of the pCAMBIA3300-GmFAX8::GFP fusion protein was observed by a laser confocal microscope (laser confocal microscope, SP8 LIGHTNING, Wetzlar, Germany).
4.6. Overexpression of GmFAX8 in Soybean Hairy Roots
The above-mentioned recombinant plasmid (pCAMBIA3300-GmFAX8::GFP) and empty pCAMBIA3300 plasmid were transferred into Agrobacterium rhizogenes K599, respectively. Soybean (DN50) was introduced using the Agrobacterium rhizogenes-mediated cotyledonary node method. The growth of hairy roots was observed by explant preparation, infection, co-culture, and rooting. The culture dish was taken out at the right time, and the residual culture medium was rinsed with distilled water for PCR detection to obtain transgenic hairy roots [32].
4.7. Determination of Total Fatty Acid Content Using the Gas Phase Method
In this experiment, gas chromatography (GC) was employed to measure the fatty acid content in the hairy roots of soybeans. We plotted a standard curve using five major fatty acids as internal standards, with the concentration unit of the internal standards being milligrams per milliliter. The x-axis of the standard curve represented the concentration (milligrams per milliliter), and the y-axis represented the peak area. After drying the above-mentioned transgenic hair-like roots, 0.1 g was weighed, ground, and added to 3 mL of n-hexane. It was extracted in a 50-degree water bath for 1 h (mixed up and down once every 5 min). After extraction, 3 mL of sodium hydroxide methanol solution (0.5 mol/L) was added, and the vortex was shaken for 30 min. It was left to stand at room temperature for 60 min. The supernatant was filtered into a 1.5 mL brown bottle. The fatty acid content was then determined using an Agilent gas chromatograph. Meteorological chromatographic conditions were as follows: quartz capillary column; the carrier gas was helium; the initial temperature of the column was set at 180 °C and kept for 1.5 min; the temperature was raised to 225 °C at 10 °C/min and kept for 2 min; the injection port temperature was set at 250 °C, and the column flow rate was 67.5 mL/min; and the split ratio was set as 20:1. All data were subjected to three biological replicates.
4.8. Determination of Triglyceride Content in Yeast
The amplified GmFAX8 gene was connected to the vector pYES3 to obtain the recombinant vector pYES3-GmFAX8. The recombinant plasmids pYES3-GmFAX8 and pYES3 were transferred into Saccharomyces cerevisiae INVSc1. The yeast was grown in a solid SD-Trp medium (containing 2% galactose) and cultured at 30 °C for 48 h. The content of TG and protein in the recombinant vector and empty vector (pYES3) was determined by the enzyme assay kit for the content of triglycerides (TGs) in tissue cells (Applygen Technologies Inc., Beijing, China) and the BCA protein quantitative Kit (Applygen Technologies Inc., Beijing, China).
4.9. Co-Expression Network Analysis of GmFAX Genes and Transcription Factors
To investigate the co-expression relationships between transcription factors and the target genes, we utilized transcriptome data from 10 soybean varieties (5 high-oil and 5 low-oil) at the R6 developmental stage [33]. Differentially expressed genes (DEGs) were first identified with a threshold of |Log2FC| > 0.6. Transcription factors and their potential target genes among these DEGs were then selected for network analysis. The co-expression network relationships were constructed using the R package version 4.4.1 (|r ≥ 0.75|, p < 0.01).
4.10. Haplotype Analysis of the GmFAX8 Gene
The whole genome of 181 different varieties of soybean varieties was re-sequenced at 30× coverage. With MAF > 0.1 as the threshold for filtering, 1,355,928 SNP markers were obtained. The general linear model (GLM) of TASSEL 5.0 software was used to analyze the sequence variations of the GmFAX8 gene based on the phenotypic values of oil content. SNPs with a threshold of −log10 (P) ≥ 2.5 were defined as significantly associated loci. The single-nucleotide polymorphisms of the gene promoter, 5′ untranslated region, exon, and 3′ untranslated region of GmFAX8 were studied.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Troncoso-Ponce M.A. Nikovics K. Marchive C. Lepiniec L. Baud S. New Insights on the Organization and Regulation of the Fatty Acid Biosynthetic Network in the Model Higher Plant Arabidopsis thaliana Biochimie 20161203810.1016/j.biochi.2015.05.01326025475 · doi ↗ · pubmed ↗
- 2Xu C. Shanklin J. Triacylglycerol Metabolism, Function, and Accumulation in Plant Vegetative Tissues Annu. Rev. Plant Biol.20166717920610.1146/annurev-arplant-043015-11164126845499 · doi ↗ · pubmed ↗
- 3Sagun J.V. Yadav U.P. Alonso A.P. Progress in Understanding and Improving Oil Content and Quality in Seeds Front. Plant Sci.202314111689410.3389/fpls.2023.111689436778708 PMC 9909563 · doi ↗ · pubmed ↗
- 4Bates P.D. Stymne S. Ohlrogge J. Biochemical Pathways in Seed Oil Synthesis Curr. Opin. Plant Biol.20131635836410.1016/j.pbi.2013.02.01523529069 · doi ↗ · pubmed ↗
- 5Rawsthorne S. Carbon Flux and Fatty Acid Synthesis in Plants Progress. Lipid Res.20024118219610.1016/S 0163-7827(01)00023-611755683 · doi ↗ · pubmed ↗
- 6Chen R. Yamaoka Y. Feng Y. Chi Z. Xue S. Kong F. Co-Expression of Lipid Transporters Simultaneously Enhances Oil and Starch Accumulation in the Green Microalga Chlamydomonas Reinhardtii under Nitrogen Starvation Metabolites 20231311510.3390/metabo 1301011536677040 PMC 9866645 · doi ↗ · pubmed ↗
- 7Li N. Xu C. Li-Beisson Y. Philippar K. Fatty Acid and Lipid Transport in Plant Cells Trends Plant Sci.20162114515810.1016/j.tplants.2015.10.01126616197 · doi ↗ · pubmed ↗
- 8Li N. Gügel I.L. Giavalisco P. Zeisler V. Schreiber L. Soll J. Philippar K. FAX 1, a Novel Membrane Protein Mediating Plastid Fatty Acid Export P Lo S Biol.201513 e 100205310.1371/journal.pbio.100205325646734 PMC 4344464 · doi ↗ · pubmed ↗