Purification, Structural Characteristics, Bioactive Properties, and Applications of Naematelia aurantialba Polysaccharides: A Comprehensive Review
Ri-Na Wu, Yun-Yang Zhu, Run-Hui Ma, Zhi-Jing Ni, Xiao-Juan Deng, Kiran Thakur, Zhao-Jun Wei

TL;DR
This paper reviews the properties and applications of polysaccharides from the golden ear fungus, highlighting their health benefits and potential in food, medicine, and cosmetics.
Contribution
The paper provides a comprehensive review of the extraction, structure, bioactivity, and industrial applications of Naematelia aurantialba polysaccharides.
Findings
N. aurantialba polysaccharides exhibit antioxidant, hypoglycemic, and anti-inflammatory effects.
The review highlights the structural characteristics and functional mechanisms of NAPs.
Current limitations and future research directions for NAPs are discussed.
Abstract
Jin’er (Naematelia aurantialba), commonly known as golden ear, is a traditional edible fungus that has long been recognized for its medicinal and culinary properties in China. Recently, it has been registered as a new cosmetic ingredient, drawing significant attention across various fields, including medicine, food, and cosmetics, due to its array of nutritional and medicinal benefits. N. aurantialba is rich in bioactive compounds, such as polysaccharides, dietary fiber, polyphenols, and active peptides. Among these, N. aurantialba polysaccharides (NAPs) are the primary active components, exhibiting a range of biological properties, including antioxidant, hypoglycemic, immunomodulatory, intestinal flora modulatory, anti-tumor, and anti-inflammatory effects. This comprehensive review summarizes the latest advancements in the extraction, purification, structural characteristics,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
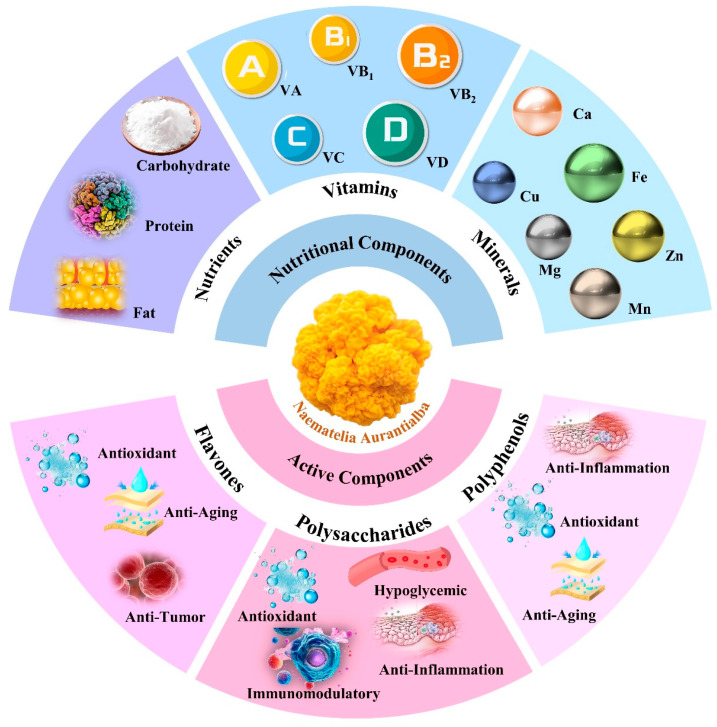 Figure 1
Figure 1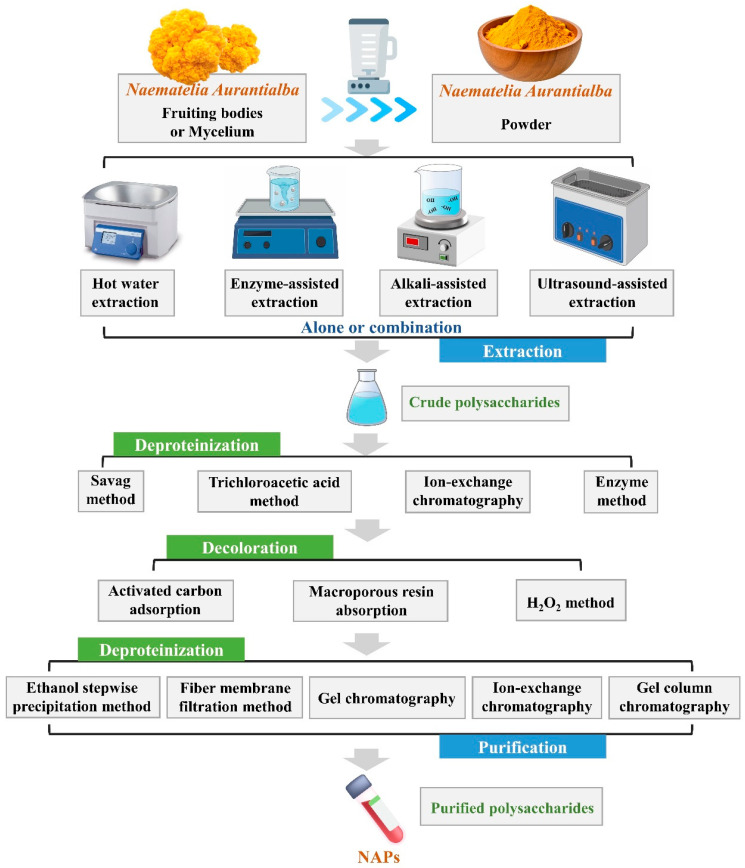 Figure 2
Figure 2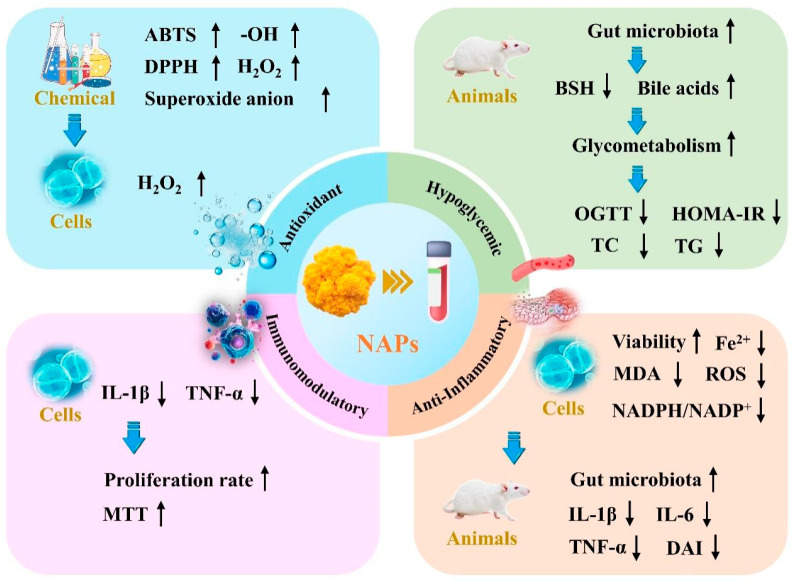 Figure 3
Figure 3- —National Natural Science Foundation of China
- —Ningxia Hui Autonomous Region Science and Technology Innovation Team Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPolysaccharides and Plant Cell Walls · Polysaccharides Composition and Applications · Seaweed-derived Bioactive Compounds
1. Introduction
Jin’er (Naematelia aurantialba (Bandoni & M. Zang) Millanes & Wedin), abbreviated as N. aurantialba, is a rare and precious edible and medicinal fungus native to China. It is commonly known as golden ear, brain ear, or golden wood ear. This fungus belongs to the order Tremellales, family Naemateliaceae, and genus Naematelia Fries [1,2,3]. Previously initially named Tremella aurantialba, following advancements in scientific research and species classification, it has been reclassified into the genus Naematelia, leading to its current designation as N. aurantialba [1,4,5]. The wild strain of N. aurantialba is predominantly found in the alpine oak forest belt, where it exhibits parasitic or partially symbiotic relationships with species such as Gramma mucronulatum and Gramma flatus. It is distributed across regions including Xizang, Yunnan, Sichuan, and Gansu provinces in China. The formation of the fruiting body of N. aurantialba is influenced by the ratio of N. aurantialba to Stereum hirsutum, complicating its artificial cultivation [6,7]. However, with the advancement of artificial breeding techniques, established technologies for the artificial cultivation of its fruiting bodies have been developed in regions such as Ningxia, Yunnan, and Zhejiang in China [1,8]. Various studies have indicated that N. aurantialba is a quality source of essential nutrients, such as proteins, amino acids, polysaccharides, fats, minerals, and vitamins. It is also abundant in functional nutrients such as polysaccharides, polyphenols, and flavonoids, which are commonly found in fungi [9,10,11,12]. The nutritional and bioactive components of N. aurantialba are shown in Figure 1. According to the text “Chinese Medicinal Fungi”, N. aurantialba is warm with a hint of cold in nature and sweet in taste. It is used to treat conditions such as lung heat, cough, asthma, hypertension, and other ailments [4,11]. Given its unique flavor, rich nutritional value, and excellent health functions, N. aurantialba is regarded as an important raw material in the food industry. It is frequently used as an active component in functional foods, including vegetable products, fruit processing, and dietary supplements [5,9,11,13,14]. In February 2025, N. aurantialba was officially registered as a new cosmetic ingredient in China, with many of its chemical components approved for use as cosmetic additives.
Polysaccharides derived from edible fungi have garnered significant scientific interest in recent decades due to their remarkable biological activities [15,16,17,18]. Systematic research on these polysaccharides offers a valuable opportunity to identify and develop innovative therapeutic agents that enhance human health [19,20,21]. Notably, N. aurantialba contains exceptionally high levels of polysaccharides, far exceeding those found in Tremella fuciformis and even higher than those in Dendrobium officinale. Given its dual status as both an edible and medicinal fungus, N. aurantialba polysaccharides (NAPs) have become a focal point of extensive scientific research [4,22]. Recent pharmacological studies have demonstrated that NAPs exhibit a wide range of biological activities, encompassing antioxidative, hypoglycemic, anti-tumor, antihypertensive, anti-inflammatory, and immunoregulatory effects. [1,13,14]. NAPs represent a promising area of fundamental research with promising biomedical applications in food science, natural products, pharmaceuticals, and pharmacology [10,23]. At present, many scholars are engaged in investigating the extraction, separation, and purification methods of NAPs and exploring the intrinsic connection between the chemical structure and physiological functions of NAPs [4,23,24].
In recent years, improvements in living standards and medical care have led to a growing emphasis on obtaining nutrients and health components from diet [25,26,27]. N. aurantialba is abundant in natural nutrients and bioactive components, particularly its polysaccharides, which are beneficial to human health and comply with the standards for health foods and nutritional supplements [5,7,11]. However, current research on the purification technology, structural characteristics, biological activities, mechanisms of action, and potential applications of NAPs remains insufficient [4,23,28]. Therefore, this review aims to summarize the research progress of NAPs over the years, covering aspects such as their extraction and purification techniques, structural characteristics, biological properties, and mechanism of action. It also addresses the current research status and gaps in the field of edible fungus polysaccharides, promoting their development and utilization in medicine and functional foods.
2. Extraction and Purification of NAPs
Polysaccharides are critical components of N. aurantialba, and their extraction and purification form the foundation for subsequent activity research. Prior to extraction, plants typically undergo pretreatment processes such as washing, drying, grinding, sieving, and crushing. The above steps are essential for removing colored contaminants, monosaccharides, oligosaccharides, and other small molecular substances [29,30]. Studies indicated that the extraction method can significantly influence the properties, structure, and functional activity of polysaccharides. Consequently, the selection of extraction methods must consider various factors, such as biological sources, target requirements, and potential interfering substances [31,32]. The extraction and purification technologies of NAPs are shown in Figure 2.
2.1. Extraction and Preparation of NAPs
The methods for obtaining NAPs can be broadly categorized into three approaches: extraction from natural or artificially cultivated N. aurantialba fruiting bodies; extraction from fermentation-cultured mycelium or fermentation broth; and extraction from fermentation-cultured N. aurantialba spore fermentation broth [33,34,35]. Table 1 presents the main extraction techniques of NAPs along with their advantages and disadvantages.
2.1.1. Extraction from Fruiting Bodies
Extraction from natural or artificially cultivated N. aurantialba fruiting bodies is the predominant method for obtaining NAPs. Over the years, researchers have optimized and innovated various extraction techniques and processes [29,31,43]. These techniques primarily include hot water extraction (HWE), enzyme-assisted extraction (EAE), ultrasound-assisted extraction (UAE), and alkali-assisted extraction (AAE). These methods are applicable not only to polysaccharide extraction from fruiting bodies but also to mycelial sources [43,44,45].
Hot water extraction (HWE) is the most traditional and commonly used method for polysaccharide extraction. It is based on the principle that most polysaccharides have high solubility in hot water and are more stable. HWE has the advantages of simple operation steps, minimal equipment requirements, and preservation of polysaccharide bioactivity, making it the preferred method of polysaccharide extraction for most edible mushrooms [31,44]. Furthermore, hot water leaching is frequently combined with other extraction methods, such as enzymatic hydrolysis, acid-base hydrolysis, physical treatments, and chemical derivatization, to yield stable and highly biologically active polysaccharides [43]. Yuan et al. extracted TAP-3 from high-quality N. aurantialba fruiting bodies by pretreating the corms and then extracting them with hot water at 95 °C for 3 h, repeating the process three times, and achieving a final yield of 50.60% [36]. However, HWE also suffers from low extraction efficiency, long extraction times, and high operating temperatures [32]. For example, Du et al. used 100 °C hot water to extract NAPs from N. aurantialba fruiting bodies, resulting in an extraction rate of only 3.84% [37].
The enzyme-assisted extraction (EAE) method utilizes biological enzymes to disrupt the cellulose and cell wall structures of edible fungi, facilitating the dissolution of polysaccharide molecules. This method offers high extraction efficiency, low energy consumption, and mild reaction conditions [46]. Compared to other methods, EAE can effectively increase polysaccharide yield without destroying the polysaccharide structure or bioactivity. However, it is not suitable for industrial production due to enzyme inactivation and high costs [45]. For example, Sun et al. employed cellulase, pectinase, and papain to extract non-starch polysaccharides (NAPs), resulting in the acquisition of TAPS-E fractions. Additionally, Sun et al. used cellulase for NAP extraction, achieving a final extraction rate of 24.95% for NAP-3 [40].
Ultrasound-assisted extraction (UAE) employs the cavitation effect of ultrasound to enhance solvent penetration, disrupt cell walls, and accelerate the release of intracellular components. This method offers several advantages, including short extraction time and extraction rates [46,47]. Several studies have shown that the structure of polysaccharides extracted by UAE can be changed, and the biological activity of polysaccharides extracted without UAE not only has a higher extraction rate, but also has a stronger functional activity [48,49]. For example, Feng et al. found that ultrasound treatment changed the polysaccharide molecular weight and monosaccharide composition ratio of Cucumis sativus polysaccharide (SSU), thus improving its antioxidant and antitumor ability compared to untreated hot water extracted Cucumis sativus polysaccharide (SSW) [50]. Huang et al. employed ultrasound extraction to obtain natural active products (NAPs) from the fruiting bodies of the N. aurantialba, achieving an extraction rate of 46.36% with high purity [41]. Similarly, Du et al. integrated hot water extraction with ultrasonic extraction methods to isolate TA 2-1 fractions with anti-inflammatory properties [42].
The alkali-assisted extraction (AAE) method is particularly effective for certain acidic polysaccharides or high-molecular-weight polysaccharides, as these polysaccharides have higher solubility in dilute alkaline solutions compared to hot water, significantly increasing the yield of polysaccharides [51,52]. For example, Deng et al. employed this method to isolate the crude polysaccharide CMCP component from the mycelium of the N. aurantialba fungus [34]. Zhao et al. extracted acidic polysaccharide components from chicken leg mushrooms using NaOH aqueous solution and further analyzed their structural characteristics and antioxidant activity [53]. Similarly, Yu et al. used EAE, UAE, AAE, and acidic aqueous extract to obtain four polysaccharide fractions (E-NAP, U-NAP, Al-NAP, and Ac-NAP) from the fruiting bodies of N. aurantialba, each exhibiting different functional properties [28].
Given the scarcity of natural or artificially cultivated N. aurantialba fruiting bodies, research on the extraction of NAPs has been limited [37,41]. The application of more efficient and intelligent emerging technologies, such as microwave-assisted extraction, subcritical water extraction, three-phase extraction, pulsed electric field-assisted extraction, nanoparticle milling extraction, and high-pressure homogenization extraction, may significantly improve polysaccharide yield and enhance the utilization of NAPs.
2.1.2. Mycelium Submerged Fermentation Pathway
Traditional methods for extracting polysaccharides from edible fungi face significant limitations, including long cultivation periods and challenges in maintaining operational consistency. These limitations negatively impact the efficiency of polysaccharide production and the uniformity of their nutritional and bioactive properties, which are crucial for their application as functional food ingredients [34,54]. In contrast, mycelium submerged fermentation is a more efficient method for producing NAPs due to its short cycle time, controllability, traceability, and large-scale production. Moreover, it can significantly enhance the yield, quality, and consistency of polysaccharides by allowing precise control over fermentation parameters, including pH, nutrient replenishment, oxygen levels, fermentation temperature, and fermentation duration [55]. Multiple studies have investigated the optimization of fermentation and extraction conditions for producing NAPs through submerged fermentation of mycelia by modifying the medium formulation and fermentation conditions [56]. Deng et al. cultured the N. aurantialba strain on potato dextrose agar (PDA) slant medium for strain acquisition, followed by transferring it to PDA plate medium for rejuvenation. The strain was then transferred to a liquid medium composed of 20% potato, 2% glucose, and 2% agar for fermentation at 28 °C. The mycelial fermentable polysaccharides were successfully obtained using alkaline water extraction [34]. Similarly, Zhang et al. utilized the T. aurantialba JSU-05 strain in a 100 mL fermentation setup and successfully obtained a purified polysaccharide, which they named TMP [39].
2.1.3. Spore Fermentation Pathway
Edible fungi from the order Tremellomycetes can produce yeast-like strameno spores, which proliferate by budding in the absence of commensal fungi. This process is easier to regulate compared to traditional mycelial fermentation, making it a viable strategy for large-scale polysaccharide production [57,58]. This method has been successful for the preparation of ginkgo polysaccharides. For example, Zhu et al. produced Tremella fuciformis polysaccharides (TFPs) from T. fuciformis spores in a 5 L stirred-tank bioreactor under pH-controlled mode, achieving a yield of 4.48 g/L. They further increased the yield of TFPS to 5.80 g/L using a three-stage pH-controlled process [59]. In another study, Sun et al. isolated and purified the N. aurantialba spore strain NX-20 and utilized it in fermentation to produce NAPs. The resulting product had the same monosaccharide composition as TAPS obtained by substrate extraction. The NAPS were produced by NX-20 in a 7.5 L fermenter at 25 °C, using tofu wastewater instead of defatted soybean meal as the raw material. The effects of fermentation temperature, duration, initial pH, and inoculum amount on the NAPs yield were systematically evaluated, ultimately achieving a maximum yield of 15.02 ± 0.40 g/L [35]. Subsequent studies have focused on the structural characterization and functional evaluation of the acquired NAPs [58,60]. Spore fermentation is considered a promising technology for NAPs production, offering significant cost savings.
2.2. Purification of NAPs
The products obtained through the aforementioned methods are crude polysaccharides mixed with amino acids, proteins, pigments, lipids, inorganic salts, and monosaccharides. These impurities can interfere with the structural analysis and functional activity evaluation of NAPs [61,62,63]. To ensure accurate structural analysis and explore the pharmacological activity of NAPs, the crude polysaccharides must be further separated and purified. Prior to the purification process, the crude polysaccharides must undergo pretreatment. Decolorization methods primarily include activated carbon adsorption, hydrogen peroxide oxidation, and ion exchange. Protein removal techniques encompass the Sevag method, trichloroacetic acid method, and protein digestion method [49,64]. The purification techniques for NAPs mainly include ethanol precipitation, fiber membrane filtration, ion exchange chromatography, and gel column chromatography. Among the various purification methods, graded precipitation of ethanol is the most commonly used technique. In recent years, several novel technologies have been integrated with the ethanol precipitation method to enhance the separation and purification of polysaccharides from N. aurantialba [4,37]. Du et al. used an ultrafiltration membrane to complete decolorization and protein removal in one step, then obtained two components, TAPA1 and TAPB1, through ion exchange chromatography and gel permeation chromatography [33,38]. Yuan et al. obtained crude NAPs from N. aurantialba fruiting bodies using hot water extraction, followed by dialysis and lyophilization. The TAP-3 fraction was further purified using a DEAE sepharose fast flow column [36]. Sun et al. isolated and purified crude polysaccharides extracted from the conidial fermentation broth using semi-permeable membrane dialysis and the Seveg method, resulting in the acquisition of NAPs [35,58,60]. Peng et al. precipitated polysaccharides using 80% ethanol, followed by further purification through ion exchange chromatography and gel chromatography to isolate the Ta 2-1 fractions [42]. Similarly, Sun et al. extracted polysaccharides from N. aurantialba fruiting bodies using ethanol precipitation, followed by dialysis and ion-exchange chromatography to obtain the NAP-3 fractions [40], which were further applied in antihyperglycemic activity studies [65].
3. Structural Characterizations of NAPs
Polysaccharides are macromolecular carbohydrates composed of multiple monosaccharides linked by glycosidic bonds. Their structural characterization can be divided into primary and advanced structures. The primary structure refers to the composition of the main chain and branched chains, while the advanced structure involves the conformation of the main chain and non-covalent interactions between polysaccharides [66,67]. As a consequence of the limitations in extraction costs and analytical techniques, the structural characterization of NAPs primarily focuses on the primary structure. This includes the analysis of the total sugar content, molecular weight and distribution, monosaccharide composition and molar ratios, and type of glycosidic bonds [29,30]. The primary techniques involved include nuclear magnetic resonance spectroscopy (NMR), high-performance liquid chromatography (HPLC), gas chromatography-mass spectrometry (GC-MS), and infrared spectroscopy (IR). As shown in Table 2, the structural features of NAPs, such as monosaccharide composition, molecular weight, chemical properties, and other structures, have been summarized [49,68].
3.1. Relative Molecular Weight
Molecular weight (M_W_) is a fundamental parameter for characterizing the physicochemical properties of natural plant polysaccharides. A significant correlation has been observed between their pharmacological activities and average molecular weights [71]. Current methods for determining the molecular weight of NAPs include high-performance liquid chromatography (HPLC), high-performance gel chromatography (HPGPC), high-performance gel filtration chromatography (HPGFC), and polyacrylamide gel electrophoresis (PAGE) [18,58]. To date, researchers have successfully extracted and purified NAPs with varying molecular weights. For instance, Fei et al. isolated a low molecular weight heteropolysaccharide (TABP) with a molecular weight of only 5408 Da from the endophytic bacterium Bacillus sp. TAB [70]. However, Sun et al. obtained higher molecular weight NAPs-25 and NAPs-30 from the fermentation broth of N. aurantialba NX-20 strain, with molecular weights of 2948.0 and 4647.0 kDa, respectively [69]. Interestingly, the same research team used the same strains for fermentation and successfully isolated NAPS-A, NAPS-B, TAPS-E, and TAPS-F with molecular weights of 2924.0, 1763.0, 1130.4, and 2924.6 kDa, respectively [35,58]. Additionally, Du et al. isolated two fractions from the fruiting bodies of N. aurantialba, TAPA1 and TAPB1, respectively, with molecular weights of 1350 and 760 kDa [33,38]. These results demonstrate that even when the same source strains are employed for fermentation or extraction of polysaccharides, significant differences in the obtained products can still exist. This variation in M_W_ may be attributed to the diverse sources of NAPs, as well as differences in extraction and purification methods.
3.2. Monosaccharide Composition
Natural plant polysaccharides are typically composed of monosaccharides with varying chemical structures linked by glycosidic bonds, resulting in a wide diversity of polysaccharide structures [72]. Consequently, a detailed analysis of monosaccharide composition is crucial to reveal the structural diversity and pharmacological activities of these polysaccharides [73]. Current methodologies for monosaccharide composition determination include high-performance liquid chromatography (HPLC), Fourier transform infrared spectroscopy (FT-IR), capillary electrophoresis (CE), and gas chromatography-mass spectrometry (GC-MS) [68,74]. Comprehensive studies have shown that NAPs are composed of various monosaccharides, including Galactose (Gal), Mannose (Man), Epichitosamine (Man-N), Glucose (Glc), Fructose (Fru), Glucosamine hydrochloride (GlcN), Glucuronic acid (GlcA), Rhamnose (Rha), Galacturonic acid (GalA), Arabinose (Ara), and Xylose (Xyl) [75]. For example, Sun et al. used the spore fermentation method to produce NAPs as an alternative to traditional fruiting body extraction. They isolated and purified four polysaccharide components from the NX-20 strain of N. aurantialba: NAPS-A, NAPS-B, TAPS-E, and TAPS-F. Among them, NAPS-A and NAPS-B are composed of mannose (Man), xylose (Xyl), glucuronic acid (GlcA), glucose (Glc), and galactose (Gal), while TAPS-E and TAPS-F consist of five monosaccharides: Man-N, Man, GlcA, Glc, and Xyl, with varying proportions [58]. Earlier, Du et al. identified only three monosaccharides, namely Man, Xyl, and GlcA, from the NAPs of the N. aurantialba fruiting bodies, specifically TAPA1 (Man:Xyl:GlcA = 5:4:1) and TAPB1 (Man:Xyl:GlcA = 3.1:2.9:1.2) [38]. In a subsequent study, Huang et al. employed ultrasonication to extract polysaccharides from the fruiting bodies, which were designated as NAPs. The monosaccharide composition of these NAPs included Man (59.04%), Xyl (23.89%), GlcA (14.07%), GalA (2.12%), and Glc (0.76%). Additionally, some other NAPs exhibited unique monosaccharide compositions, such as Fru and GlcN found in TA 2-1 and TABP, respectively [41].
3.3. Structural Characteristics
The chemical structures of polysaccharides, such as the length of the sugar chain, linkage types, degree of branching, and types of monosaccharide residues, collectively determine their biological activities [29,46,47,48]. Consequently, an in-depth investigation of the chemical structure of NAPs is essential for elucidating their physiological functions. Current methods for studying the chemical structure of NAPs include high-performance gel permeation chromatography (HPGPC), high-performance liquid chromatography (HPLC), gas chromatography-mass spectrometry (GC-MS), Fourier-transform infrared spectroscopy (FT-IR), nuclear magnetic resonance (NMR), atomic force microscopy (AFM), transmission electron microscopy (TEM), and scanning electron microscopy (SEM), and so on [43,49]. The methods not only determine the type and configuration of glycosidic bonds in NAPs, but also facilitate the accurate analysis of substituent types, repeating units, and the linkage order within polysaccharide chains [63]. NMR is a common technology employed for the structural analysis of polysaccharides, as it can identify the types of glycosidic bonds, the specific monosaccharides present, and the sequence of glycosidic linkages through chemical shifts. In contrast, FT-IR is typically used to ascertain the types of polysaccharides and some aspects of their configurations via characteristic absorption peaks. Complementing these techniques, LC-MS/MS and MALDI-TOF/MS provide direct sequence evidence by analyzing oligosaccharide fragmentation patterns, enabling the determination of glycosidic linkage sequences and branching points with high sensitivity, which may play a crucial role in the future structural analysis of NAPs [48,51]. Additionally, SEM and AFM are commonly engaged for analyzing the surface topography of polysaccharides [72,74]. Significant progress has been made in the structural characterization of NAPs. Du et al. structurally characterized two NAPs separately, identifying four residues in TAPA1 as →3)-α-D-Manp-(1→3)-α-D- Manp-(1→3)-α-D-Manp-(1→, 4-β-D-Manp-(1→3)-β-D-Xylp-(1→4)-β-D-ClcAp-(1→2)-α-D-Manp-(1→, α-D-Manp-(1→4)-β-D-Xylp-(1→2)-α-D-Manp-(1→,β-D-Xylp-(1→2)-β-D-Xylp-(1→4) -α-D-Manp-(1→; Meanwhile three residues were detected in TAPB1 as →3)-α-D -Manp-(1→3)-α-D-Manp-(1→3)-α-D-Manp-(1→,β-D-ClcAp-(1→3)-β-D-Xylp-(1→2)-α-D-Manp-(1→, β-D-Xylp-(1→2)-β-D-Xylp-(1→4)-α-D-Manp-(1→ [38]. With advancements in technology and a deeper understanding of polysaccharide structures, researchers have successfully identified not only the types of sugar residues but also their respective proportions. For instance, in the study by Sun et al., the proportions of various residues in NAPS-25 were reported as follows: 2,3,4-Me3-Xylp:2,4-Me2-Xylp:3,4-Me2-Xylp:2,3,4,6- Me4-Manp:2,3,6-Me3-GlcAp:2,4,6-Me3-Manp:6-Me-Manp = 10.27:7.07:13.58:31.00:2.45:20.05:15.58. In contrast, the ratios of several residues in NAPS-30 were 2,3,4-Me3-Xylp:2,4-Me2-Xylp:3,4-Me2-Xylp:2,3,4,6-Me4-Manp:2,3,6-Me3-GlcAp:2,4,6-Me3-Manp:6-Me-Manp = 8.69:8.17:13.95:26.76:2.85:20.62:18.96 [69]. In a subsequent study, the structure of NAPS-A was elucidated through methylation analysis and NMR, revealing sugar residues such as →3)-α-D-Manp, →2,3,4)-α-D-Manp-(1→, →3)-α-D-Manp-(1→, β-D-Manp-(1→, →2)-β-D-Xylp-(1→, β-D-Xylp-(1→, and →3)-β-D-Xylp-(1→. SEM and TEM were used to analyze the microstructure of NAPS-A, while XRD analysis showed that the polysaccharide exhibited bun-shaped non-crystalline structures [58].
The activity-structure relationship of NAPs remains underexplored. Sun et al. found that NAPS-A (4647 kDa) preferentially binds with water molecules in solution, reducing free water and ice crystal formation, thereby enhancing antifreeze quality and making it a valuable additive for frozen foods [58]. Conversely, lower molecular weight TABP (5408 Da) effectively promotes microbial growth as a prebiotic [70]. Du et al. proposed that sulphonation and acetylation of TAPA1 enhance its immunostimulatory and antioxidant activities, respectively [38]. Additionally, Yuan et al. demonstrated that the immunomodulatory activity of TAP-3 correlates with its chain length, with the low molecular weight dTAP-3a fragment showing superior activity [36].
4. Biological Activity of NAPs
As a well-known fungus recognized for its dual role as both a food source and medicinal agent, its traditional pharmacological effects are extensively documented in classical Chinese medicine texts. For instance, the “Record of Famous Doctors” (5th–6th Century AD, Tao Hongjing, Late Liang to Early Tang Dynasty) notes that N. aurantialba benefits brain health and helps dispel cold effects. In addition, the “Compendium of Materia Medica” (Li Shizhen, 1578 AD, Ming Dynasty) from the Ming dynasty records its use in treating various diseases, highlighting its efficacy in moistening the lungs, relieving cough, preserving liver function, and tonifying the kidneys [1,5]. With the increasing attention to the physiological efficacy of traditional medicinal and food products, the modern chemical composition of N. aurantialba has been thoroughly analyzed, confirming its richness in dietary fiber, polyphenols, active peptides, polysaccharides, and other functional substances. Among these, polysaccharides are identified as the most significant active components in N. aurantialba, demonstrating various beneficial effects, including antioxidant, hypoglycemic, immune regulation, intestinal microbiota modulation, anti-tumor, and anti-inflammatory properties [23,36,37]. Figure 3 illustrates the main functional activities and related mechanisms of NAPs, while Table 3 summarizes the key research progress on their physiological activities.
4.1. Antioxidant Activity
Oxidative stress is a disorder associated with an imbalance in redox mechanisms, typically resulting from an excess of reactive oxygen species (ROS) or reactive nitrogen species (RNS) in the body, which can cause cellular damage through direct or indirect means [80,81]. NAPs exhibit potent antioxidant activity and can protect cells and tissues from damage by effectively scavenging free radicals and alleviating oxidative stress. Both in vivo and in vitro studies have investigated the antioxidant activity of NAPs [37,69,76]. In the study conducted by Du et al., TAPA1 and TAPB1, derived from fruiting bodies, demonstrated extremely strong superoxide anion and H_2_O_2_ scavenging capabilities in vitro. In a subsequent study, a PC12 cell model of oxidative injury induced by H_2_O_2_ was treated with a 500 µg/mL solution of TAPA1, resulting in a cell survival rate increase from 56.10% to 157.63% [23,33,76]. Similarly, Huang et al. reported that NAPs obtained from substrates effectively scavenged ABTS, DPPH, and superoxide anion radical [41]. In an in vivo study, NAP-3 reduced malondialdehyde (MDA) levels in the serum of hyperlipidemic mice by regulating gene expression while increasing catalase (CAT) and glutathione (GSH) levels, thereby alleviating oxidative stress in these mice [40]. It is noteworthy that NAPs obtained from tansy spore fermentation have similar effects. Sun et al. also demonstrated that NAPS-A and NAPS-B exhibited strong scavenging activities against ABTS, DPPH, OH, and superoxide anion radicals, while NAPS-25 and NAPS-30 effectively scavenged DPPH and superoxide anion radicals [58,69]. Furthermore, Zhang et al. reported that TMP extracted from mycelium significantly increased total antioxidant capacity (T-AOC), catalase (CAT), superoxide dismutase (SOD), and glutathione (GC) levels, while reducing malondialdehyde (MDA) content in mouse tissues, effectively alleviating oxidative stress in type 2 diabetic rats [39].
4.2. Hypoglycemic Activity
Type 2 diabetes mellitus (T2D) is characterized by a progressive and uneven loss of insulin secretion from pancreatic β-cells, often following insulin resistance (IR). This condition is a component of the metabolic syndrome and accounts for approximately 96% of diabetes mellitus cases [82,83,84]. Polysaccharides derived from various edible fungi have been shown to effectively improve hyperglycemia and diabetes through glucose metabolism regulation and enhanced insulin sensitivity and resistance [85,86]. For example, Sun et al. demonstrated that NAP-3 improved HepG2 cell damage by increasing glucose consumption and reducing ROS generation. In a T2D mouse model, the combination of NAP-3 and metformin showed significant hypoglycemic activity, effectively reducing body weight, serum insulin level, glucose tolerance, and insulin tolerance [40]. Their follow-up study further revealed that NAP-3 enhanced metformin’s efficacy on lipid and glucose metabolism in T2D mice through a gut microbiome-dependent mechanism. This enhancement amplified the effect of metformin on glucose metabolism in T2D mice via the gut microbial-bile acid-nuclear receptor-GLP-1 interaction axis, effectively ameliorating diabetes [65]. In another study, TAP effectively reduced plasma glucose levels by increasing insulin levels in mice, with further studies demonstrating TAP improved diabetes by increasing hepatic glucose metabolism through elevated activities of glucokinase, hexokinase and glucose 6-phosphate dehydrogenase in vivo [77,78,79]. Therefore, NAPs represent a promising therapeutic option for diabetes treatment.
4.3. Immunomodulatory Activity
Immunomodulation is crucial for maintaining the balance of the immune system, ensuring effective defense against pathogens while preventing host tissue damage [87]. NAPs, as natural edible fungus polysaccharides, have significant immunomodulatory functions [5]. Yuan et al. reported that TAP-3 significantly enhanced macrophage secretion of nitric oxide (NO), interleukin-1 beta (IL-1β), and tumor necrosis factor-alpha (TNF-α), functioning as an immunostimulatory agent [36]. Du et al. compared the immunostimulatory activities of crude, semi-purified, and purified polysaccharides extracted from N. aurantialba using mouse spleen lymphocytes. The results confirmed that the purified polysaccharide TAPA1 exhibited the strongest immunomodulatory activity, with concentrations of 50, 200, and 500 μg/mL increasing mouse spleen lymphocytes proliferation rates to 339.67%, 484.10%, and 593.98%, respectively [37,38].
4.4. Other Activity
In addition to the above activities, NAPs possess other pharmacological activities, such as anti-inflammatory, anti-tumor, regulation of gut microbiota, hypolipidemic activity, and life-extension properties. Anti-tumor activity is usually associated with immune modulation, and NAPs have demonstrated some anti-tumor effects [36,38,87]. Lee et al. reported that crude polysaccharides extracted from the fruiting bodies of N. aurantialba exhibited no significant toxicity to RAW 309 CR.1 and Sarcoma 180 cells but extended the lifespan of Sarcoma 180-inoculated homozygous mice by 11.1% to 66.7% through immune response modulation [88]. Oxidative stress is closely linked to inflammatory responses, and NAPs with strong antioxidant activity play a crucial role in preventing certain inflammatory conditions [80,81]. Peng et al. demonstrated that the small molecule TA 2-1 significantly reduced ulcerative colitis symptoms in a mouse model by regulating intestinal flora and inhibiting iron apoptosis in epithelial cells [42]. Disturbances in gut microbiota can induce various metabolic diseases, such as hyperlipidemia, diabetes, inflammatory responses, and cancer [89,90]. Sun et al. and Peng et al. proved that NAP-3 and TA 2-1 alleviated T2D and ulcerative colitis in mice by modulating intestinal flora [42,65]. Fei et al. reported that TABP significantly promoted the growth of Lacticaseibacillus paracasei and Lacticaseibacillus rhamnosus strains (growth rates exceeding 90%) [70]. Zhang et al. illustrated that TMP effectively regulated lipid metabolism, reducing cholesterol, phospholipids, and triglycerides in the plasma of diabetic rats [39]. Overall, NAPs exhibit various biological activities, positioning them as a promising category of active ingredients derived from edible mushroom sources with broad applications.
5. Application of NAPs
In recent years, the increasing emphasis on health has driven a growing interest in natural products. Nutrients derived from plants or edible fungi, which are known for their beneficial effects, hold significant potential in fields such as functional foods, clinical medicine, and cosmetics [25,27,91,92]. N. aurantialba, with its superior functional characteristics, has gained recognition for its significant value in production and application, leading to wide-spread utilization across various fields [4,23]. China has a long history of consuming N. aurantialba, which is regarded as a functional raw material for both culinary and medicinal purposes. Compared to other regions, its applications in China are more diverse, and it has been widely accepted as a food ingredient [1,5,93]. For example, Cheng et al. investigated the impact of N. aurantialba powder on the gelatinization characteristics of starch and subsequently prepared starch jelly [94]. Feng et al. reported the preparation of an N. aurantialba bread, which significantly improved the sensory acceptability of the bread and effectively inhibited glucose release. This finding plays an important role in the development and application of starch-based products [13]. Zhang et al. fermented cigar filler leaves using Tremella aurantialba SCT-F3 (CGMCC No. 23831), thereby enhancing the sensory quality of cigar filler leaves [95]. Similarly, NAPs have been utilized in the development of a variety of functional foods and cosmetics. Sun et al. reported that the macromolecular polysaccharide NAPS-A exhibited significant antioxidant and antifreeze properties, providing a chemical and biological basis for the development of novel functional food additives and antifreeze agents [58]. Owing to its excellent water retention properties, N. aurantialba is commonly used in the development of various cosmetic products. As the functional value of NAPs is gradually realized and new preparation processes emerge that significantly reduce their production cost of NAPs. NAPs are expected to be further utilized in the development of more products, with broad application prospects and high economic value.
6. Conclusions and Future Perspectives
Natural polysaccharides, including NAPs, are emerging as promising candidates for functional food supplements and pharmaceutical development due to their abundant sources, high safety profile, minimal side effects, and diverse bioactivities. This review comprehensively examines the extraction, purification, structural characterization, bioactivity and application of NAPs in food, cosmetics and other industries, highlighting their practical value and research potential. NAPs exhibit a wide range of functional activities, including antioxidant, hypoglycemic, immunomodulatory, intestinal flora regulation, antitumor, and anti-inflammatory effects. These bioactivities are influenced by various factors, including raw material sources, extraction methods, purification techniques, molecular weight, structural characteristics, and chemical modifications, which present both opportunities and challenges for their application in disease management and industrial production.
Despite progress in NAPs research and the registration of N. aurantialba as a new cosmetic ingredient, several challenges remain. These include challenges in its extraction and purification, structural analysis, mechanism of action, clinical validation and industrial application, which need to be further explored in depth. Most studies on NAPs are confined to laboratory settings, with a significant gap remaining between experimental findings and practical industrial applications. Optimizing industrial production technologies is thus a priority. The efficiency of polysaccharide extraction and purification is a critical bottleneck. The biological activity of NAPs is closely related to their structural characteristics, and different extraction methods yield polysaccharides with varying structures. Therefore, optimizing extraction and purification procedures based on specific requirements is essential. Structural analysis of NAPs remains a significant hurdle. While current research has focused on monosaccharide composition and major polysaccharide residues, in-depth analysis of the backbone structure and specific structural features is lacking. This gap constrains the investigation of structure-function relationships in NAPs. The mechanisms underlying NAPs’ biological effects, such as hypoglycemic and hypolipidemic actions, are not fully understood. Moreover, while in vitro and animal studies are prevalent, clinical research remains limited.
Given the substantial industrial and clinical potential of NAPs, future research should focus on several key areas: (1) innovative preparation processes, including developing novel production methods such as strain screening and cultivation to reduce industrial costs; (2) integrated mechanistic studies, combining in vitro, animal, and clinical experiments to elucidate the mechanisms of NAPs’ biological activities; (3) nano delivery systems, exploring new technologies to enhance NAPs’ bioavailability; and (4) artificial intelligence integration, using AI to optimize interactions between NAPs and other food components, thereby reducing application costs and maximizing functional value. In summary, addressing these challenges and harnessing emerging technologies will be essential for advancing the research and applications of NAPs in functional foods, pharmaceuticals, and other industries.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wang Z. Huang K. Pu K.L. Li L. Jiang W.X. Wu J. Kawagishi H. Li M.L. Qi J.Z. Naematelia aurantialba: A comprehensive review of its medicinal, nutritional, and cultivation aspects Food Med. Med. Homol.20252942007210.26599/FMH.2025.9420072 · doi ↗
- 2Elkhateeb W.A. Daba G.M. Mycotherapy of the good and the tasty medicinal mushrooms Lentinus, Pleurotus, and Tremella Pharm. Pharmacol. Res.20214010610.31579/2693-7247/29 · doi ↗
- 3Zhu B.H. Jing S. Nian S.W. Xi Y.F. Xu H.D. Mao Y.M. Qian D. Kou L.P. Moisture state and volatile flavor behavior characterization of Naematelia aurantialba during postharvest in modified atmosphere packaging storage after treated with ultraviolet radiation C Postharvest Biol. Technol.202522711361110.1016/j.postharvbio.2025.113611 · doi ↗
- 4Yan Y.H. Wang M.T. Chen N. Wang X. Fu C.H. Li Y.M. Gan X.R. Lv P. Zhang Y. Isolation, structures, bioactivities, application and future prospective for polysaccharides from Tremella aurantialba: A review Front. Immunol.202213109121010.3389/fimmu.2022.109121036569950 PMC 9773546 · doi ↗ · pubmed ↗
- 5Yan Y.H. Wang M.T. Gan X.R. Wang X. Fu C.H. Li Y.M. Chen N. Lv P. Zhang Y. Evaluation of pharmacological activities and active components in Tremella aurantialba by instrumental and virtual analyses Front. Nutr.20229108358110.3389/fnut.2022.108358136570135 PMC 9767953 · doi ↗ · pubmed ↗
- 6Geng X.R. Yang D.X. Zhang Q.Y. Chang M.C. Xu L.J. Cheng Y.F. Wang H.X. Meng J.L. Good hydrolysis activity on raffinose family oligosaccharides by a novel α-galactosidase from Tremella aurantialba Int. J. Biol. Macromol.20201501249125710.1016/j.ijbiomac.2019.10.13631739012 · doi ↗ · pubmed ↗
- 7Sun T. Zhang Y.X. Jiang H. Yang K. Wang S.Y. Wang R. Li S. Lei P. Xu H. Qiu Y. Whole genome sequencing and annotation of Naematelia aurantialba (Basidiomycota, Edible-Medicinal Fungi)J. Fungi 20218610.3390/jof 8010006 PMC 877797235049946 · doi ↗ · pubmed ↗
- 8Lan J.Q. Zhang Y.K. Cai Y.L. Shi X.F. Zhang K.X. Huang J.C. Yang C.M. He X.H. Yu F.Q. Liu W. Spatial ratio of two fungal genotypes content of Naematelia aurantialba and Stereum hirsutum in nutritional growth substrate and fruiting bodies reveals their potential parasitic life cycle characteristics J. Agr. Food. Res.20252210210110.1016/j.jafr.2025.102101 · doi ↗