Role of the Inflammasome Pathway According to the Expression of Proteins and Genetic Polymorphisms in COVID-19 Patients
Thiago Rodrigues dos Santos, Lucas Baena Carstens, Leonardo Vinícius Barbosa, Mariana Collete, Natan de Araujo, Caroline Busatta Vaz de Paula, Marina Luise Viola Azevedo, Ana Clara de Almeida, Seigo Nagashima, Lucia de Noronha, Cleber Machado-Souza

TL;DR
This study explores how the inflammasome pathway contributes to severe COVID-19 by analyzing protein expression and genetic variations in patients who died during the second wave.
Contribution
The study identifies specific proteins and genetic polymorphisms in the inflammasome pathway associated with severe COVID-19 outcomes.
Findings
Lung tissue expressions of TLR-4, NLRP3, and IL-18 were significantly higher in second wave patients.
Genetic variants in NFKB1 and NOX4 were linked to increased protein expression in severe cases.
IL-18 showed strong immunological relevance tied to genetic variations in second wave cases.
Abstract
COVID-19 severity is frequently linked to exacerbated inflammation, with the inflammasome pathway playing a key role in activating inflammatory interleukins. This observational post-mortem study evaluated the expression of inflammasome-associated molecules in patients who died from COVID-19 during the second wave. Minimally invasive autopsies were performed on patients from the first (n = 24) and second (n = 18) waves. Lung tissue samples underwent immunohistochemical staining for ACE-2, TLR-4, NF-κB, TNF-α, NOX4, NLRP3, ASC, CASPASE-1, IL-1β, IL-18, GSDMD, and CASPASE-9. Additionally, genetic polymorphisms within inflammasome-related genes were assessed via real-time polymerase chain reaction. Lung tissue expressions of TLR-4, NLRP3, and IL-18 were significantly higher in patients from the second wave compared to those from the first, with expression levels of 26.3 versus 12.1, 13.9…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
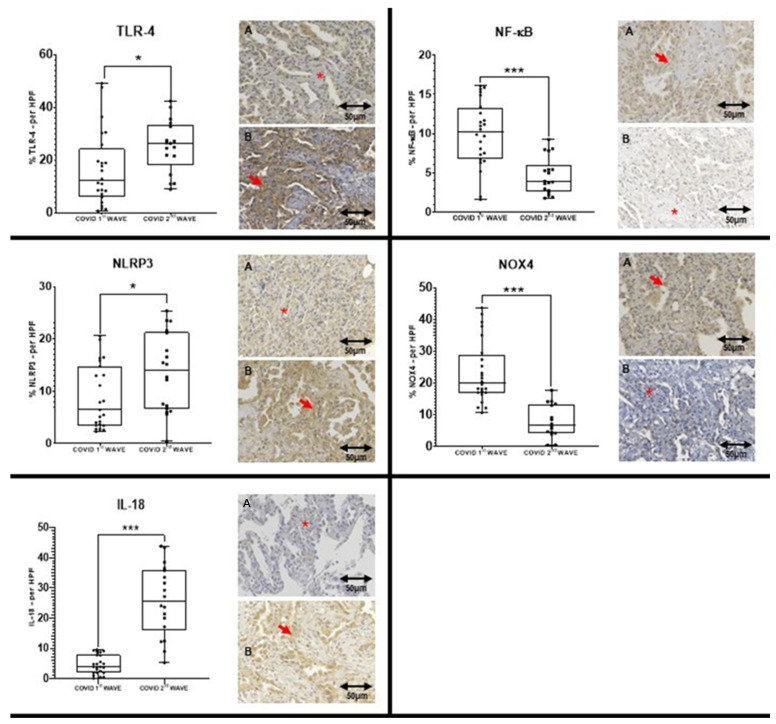 Figure 1
Figure 1- —Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)—Brasil
- —Fundação Araucária de Apoio ao Desenvolvimento Científico e Tecnológico do Estado do Paraná (FA)—Brasil
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInflammasome and immune disorders · COVID-19 Clinical Research Studies · Kawasaki Disease and Coronary Complications
1. Introduction
Several host-related risk factors have been identified as contributors to severe disease and increased mortality in coronavirus infections [1,2]. Current research indicates that the most severe COVID-19 cases typically involve clinical features such as lymphopenia, hypoalbuminemia, elevated lactate dehydrogenase, increased C-reactive protein, ferritin, and D-dimer levels. These alterations often accompany an exaggerated inflammatory response termed the “cytokine storm” [3,4,5]. Elevated cytokine concentrations contribute to the pathogenesis of severe acute respiratory syndrome (SARS) caused by SARS-CoV-2, where the virus triggers intense inflammation that can exacerbate clinical decline and increase the risk of fatality.
Central to inflammation is the activation of the inflammasome, a multi-protein intracellular complex that orchestrates inflammatory responses and induces a form of programmed cell death known as pyroptosis [6,7]. The inflammasome responds to diverse stimuli, including pathogen-associated molecular patterns (PAMPs), damage-associated molecular patterns (DAMPs), nucleic acids, and pore-forming toxins. Key molecules such as NLRP3, ASC, pro-caspase-1, pro-IL-1β, and IL-18 have established roles in inflammasome activation [8,9,10]. Extensive research has implicated the inflammasome pathway in numerous diseases, and during the COVID-19 pandemic, new and repurposed drugs targeting this pathway, including disulfiram and biologics like Anakinra^®^ and Canakinumab^®^, were evaluated for modulating the inflammatory cascade related to pyroptosis and endothelial dysfunction [11,12,13]. These molecular components represent promising candidates for further study.
Genetic variation across the human genome, particularly single-nucleotide polymorphisms (SNPs), can significantly influence gene expression and function. While many SNPs exert minimal effects, select polymorphisms may affect disease susceptibility, clinical outcomes, and response to treatment [14,15,16]. Consequently, genetic variants may serve as important components in future biomarker discovery and understanding individual risk profiles.
Biomarkers—both genetic and protein—offer measurable insights into biological states, facilitating diagnosis, prognosis, and prediction of treatment responses. The expression of inflammasome-related proteins may be modulated by specific polymorphisms, underscoring the potential of these molecules as indicators of severity and therapeutic targets in COVID-19. Given that the second wave of COVID-19 in Brazil yielded higher mortality, this study hypothesizes that differential expression and polymorphisms in inflammasome pathway genes among deceased patients could highlight molecules of biological relevance for future pandemics. Accordingly, this study aims to investigate the relationship between SNPs in inflammasome-associated genes and protein expression levels in post-mortem lung tissues from COVID-19 patients, using the second wave as a reference for outcome analysis.
2. Results
2.1. Sociodemographic and Clinical Information
Table 1 compares demographic and clinical variables between the first and second COVID-19 waves. Patients in the first wave were significantly older, with a median age of 72.5 years versus 55.5 years in the second wave (p < 0.0001). Mechanical ventilation duration was longer in the second wave group (median 14.5 days) compared to the first wave (9.5 days, p = 0.040). No significant difference was observed in hospital stay length until death (p = 0.430).
2.2. Immunohistochemistry Findings
Immunohistochemistry revealed significantly elevated expressions of TLR-4, NLRP3, and IL-18 in lung tissue from second wave patients compared to the first (median percentages: 26.3 vs. 12.1, 13.9 vs. 6.4, and 25.6 vs. 3.8, respectively). After Bonferroni correction for multiple comparisons, only IL-18 maintained statistical significance. Other markers such as ACE-2 and GSDMD showed higher expression in the second wave but without statistical significance, while NF-κB and NOX4 expression were lower in the second wave (Table 2, Figure 1).
2.3. Genotyping Findings
Table 3 summarizes genotype frequencies across COVID-19 waves. Significant differences were noted with elevated homozygosity for ACE2 rs4646188 AA (p = 0.000), rs4646156 AA (p = 0.028), and rs2048683 GG (p = 0.028) polymorphisms in the second wave. For TLR4 rs10759932 CT genotype, a higher frequency was observed in the second wave (p = 0.000). After Bonferroni correction (p < 0.001), significance persisted for ACE2 rs4646188 and TLR4 rs10759932. No significant results were found in the first and second wave of patients in grouping (dominant and recessive analysis (see Supplementary Tables S1 and S2).
Further genotype clustering analyses linked the NFKB1 rs4648090 AA genotype and NOX4 rs317155 TT + TC cluster with higher protein expression in second wave lung tissues (p = 0.022 and p = 0.008, respectively), although these associations lost significance after strict correction. IL-18-related polymorphisms (rs1946518, rs187238) showed expression trends with biological plausibility despite lacking statistical significance (Table 4).
The main analysis of this study was conducted to verify a possible association between genotypes and high tissue expression using genotype clustering models (dominant and recessive) in the first and second waves. The AA genotype for rs4648090 [G/A] of NFKB1 (p = 0.022) and the TT + TC cluster for rs317155 [T/C] of the NOX4 gene (p = 0.008) were significantly associated with the highest expression of the proteins encoded by these genes in the cohort of patients affected by the second wave of COVID-19 (Table 4); however, before the Bonferroni correction (p < 0.001), this association was lost. Considering that in the first round of analysis only IL-18 remained significant after statistical correction, the SNPs in the IL18 gene, despite not presenting statistical significance, could acquire some biological plausibility. The highest expressions of IL-18 can be associated with the GG + GT (rs1946518 G/T), and CC + CG (rs187238 C/G) grouping genotypes, respectively. No significant results were found in the first wave of patients in this analysis (see Supplementary Tables S3 and S4).
Multivariate analyses were performed, but no significance was found for any of the variables.
3. Discussion
This study aimed to further elucidate marked differences in the expression of inflammasome-related proteins and associated genetic polymorphisms between COVID-19 fatal cases from the first and second pandemic waves in Brazil. Our findings contribute to understanding the molecular underpinnings that may explain the increased severity and mortality characterizing the second wave.
The COVID 1st WAVE group exhibited a higher mean age with mostly older patients. In contrast, the COVID 2nd WAVE group predominantly comprised younger patients, which aligns with the findings reported in the extant literature [17]. The age is a major risk factor for severe COVID-19, since older patients are more vulnerable due to immune senescence and chronic low-grade inflammation [18], every ten years the risk of death rises by 1.5 times [2,19]. Despite the higher risk, the second wave of COVID-19 in Brazil was three times deadlier than the first, affecting the entire population [20]. This demonstrates that other major aspects were involved in the severity of the disease. The clinical variables not addressed in relation to the 1st and 2nd waves in this sample were not part of the analyses of this article as they have already been the subject of discussion in other articles of the group [21,22,23,24,25,26,27,28,29,30].
Immunohistochemical analysis (Table 2) revealed significantly elevated lung tissue expression of TLR-4, NLRP3, and IL-18 during the second wave, with IL-18 expression showing robust statistical significance even after stringent Bonferroni correction. This pronounced IL-18 elevation aligns with the literature recognizing IL-18 as a critical pro-inflammatory cytokine driving pulmonary inflammation and systemic cytokine storm in severe COVID-19 [31,32]. The enhanced TLR-4 expression mirrors its role as a pattern recognition receptor that can be activated by SARS-CoV-2 components and oxidized phospholipids, stimulating innate immune responses and perpetuating inflammation [33,34,35,36]. Similarly, increased NLRP3 levels reflect inflammasome assembly and activation, consistent with its established contribution to IL-1β and IL-18 maturation and subsequent pyroptotic cell death in infected tissues [8,9,10,37,38].
ACE-2 and GSDMD also demonstrated higher expression in the second wave, although without statistical significance. ACE-2 plays a dual role as the viral entry receptor and regulator of inflammatory pathways [39,40,41,42,43,44,45,46]. Elevated ACE-2 expression in post-mortem tissues may indicate enhanced viral tropism or compensatory mechanisms during later disease stages. GSDMD mediates pyroptotic membrane pore formation, further amplifying inflammasome downstream effects [47,48]. Conversely, NF-κB and NOX4 levels were paradoxically reduced in the second wave. Given NF-κB’s role in transcriptional regulation of inflammatory genes and NOX4’s involvement in reactive oxygen species generation, their reduced expression may represent negative feedback or cell exhaustion phenomena arising during sustained inflammation [8,9,49,50,51,52,53,54].
Genetic analysis (Table 3) uncovered higher frequencies of homozygous genotypes for ACE2 rs4646188 AA, rs4646156 AA, and rs2048683 GG in second wave patients, polymorphisms previously implicated in susceptibility to severe pulmonary and cardiovascular pathologies [42,43,44,45,46]. The enrichment of TLR4 rs10759932 CT genotype aligns with reports linking this variant to altered immune responses in chronic lung diseases [35,55]. Clustering analyses (Table 4) further demonstrated that NFKB1 rs4648090 AA and NOX4 rs317155 TT + TC genotypes associate with increased protein expression, underscoring the genetic influence on inflammasome pathway activity. Though statistical significance was lost after correction, these results present biologically plausible mechanisms by which host genetics exacerbate inflammasome-mediated injury.
The observed shift toward younger patients with prolonged ventilation during the second wave points to changes in clinical phenotype potentially driven by viral evolution and immune landscape remodeling [56,57]. Enhanced inflammasome activation likely exacerbates lung tissue damage through sustained cytokine release and pyroptosis, contributing to increased mortality despite demographic differences [58,59].
Our data help support a model in which inflammasome activation may contribute as a central factor in the severe pathology of COVID-19, intensified in the second wave by both increased cytokine expression and host genetic predisposition. This reinforces the promise of inflammasome components, particularly IL-18, as future biomarkers and potential therapeutic targets [11,12,13,60]. Future interventions aimed at modulating inflammasome activation have the potential to improve outcomes, especially for genetically susceptible individuals.
This study’s limitations include sample size constraints with associated power limitations, especially for genetic analyses, and the inability to temporally track longitudinal changes due to the post-mortem study design. Nonetheless, the integrated immunohistochemical and genetic approach offers valuable insights into host–pathogen interactions driving COVID-19 mortality across epidemic waves.
4. Materials and Methods
4.1. Study Population
This study was conducted following the Declaration of Helsinki and approved by the National Research Ethics Committee (CONEP; Project ID: 30188020.7.1001.0020). Families of deceased COVID-19 patients provided informed consent for minimally invasive post-mortem lung biopsies. The cohort consisted of 42 patients admitted to the intensive care unit at Hospital Marcelino Champagnat, Curitiba, Brazil. Samples included 24 patients from the first COVID-19 wave (June–August 2020) and 18 from the second wave (April–May 2021). All patients had SARS-CoV-2 confirmed by RT-PCR performed on nasopharyngeal swabs during hospitalization. Patients with negative SARS-CoV-2 tests or without consent were excluded.
4.2. Tissue Processing and Immunohistochemistry
Post-mortem lung biopsy fragments were formalin-fixed, paraffin-embedded, and assembled into tissue microarray (TMA) blocks. Sections (4 μm) were mounted on positively charged slides. Immunohistochemistry was performed using primary monoclonal antibodies against ACE-2, TLR-4, NF-κB, TNF-α, NOX4, NLRP3, ASC, CASPASE-1, IL-1β, IL-18, GSDMD, and CASPASE-9. Primary antibody incubation occurred overnight at 2–8 °C. Visualization employed horseradish peroxidase conjugated polymer detection with diaminobenzidine substrate, followed by Harris hematoxylin counterstain. Positive control reactivity confirmed assay specificity. The authors will indicate only those markers that show significantly higher tissue expression in the second wave group, as the first wave was recently reported for these samples [26].
4.3. Morphometric Analysis
Slides were digitally scanned (Axio Scan Z1, Carl Zeiss—ZEISS, Jena, Germany) producing approximately 500 high-power field images per sample. Non-representative images (e.g., bronchioles, artifacts) were excluded. This process was performed by a single researcher, ensuring the homogeneity of the selection. Thirty representative images per case were randomly selected for quantification. Image ProPlus^®^ software version 4 (Media Cybernetics, Rockville, MD, USA) semi-automatically measured immunostained area, expressed as a percentage of total tissue area, averaged per patient.
4.4. DNA Extraction and Genotyping
Genomic DNA was extracted from paraffin-embedded tissue using phenol-chloroform deparaffinization. DNA purity was assessed spectrophotometrically (OD 260/280). SNP selection targeted (Available at: https://snpinfo.niehs.nih.gov accessed on 30 July 2024) twelve inflammasome pathway genes (ACE2, TLR4, NFKB1, TNFA, NOX4, NLRP3, ASC, CASP1, IL1B, IL18, GSDMD, CASP9) based on SNPinfo database (parameters tuned for a CEU population proxy for the southern Brazilian population. The Brazilian population is heterogeneous and has overlapping genotypes due to miscegenation, but the authors used the CEU population due to its greater proximity to the population of southern Brazil where the study was carried out. Tag SNPs with functional relevance from literature were included. Genotyping was performed by real-time PCR using TaqMan^®^ probes on an Applied Biosystems 7500 system (Foster City, CA, USA). Genotypes were called automatically based on fluorescent signal ratios; homozygous genotypes produce single fluorophore signals, and heterozygotes display dual signals. Ten percent of samples were re-run to assess reproducibility.
4.5. Statistical Analysis
Categorical variables were represented as counts and percentages, continuous variables as medians with ranges. Differences between waves were assessed by Pearson’s chi-square, Fisher’s exact, or Mann–Whitney tests as appropriate. Logistic regression examined genotype-phenotype associations. Bonferroni adjustment corrected for multiple comparisons. Statistical significance was set at p < 0.05 before correction. For the multivariate analysis, the binary logistic regression model was used and included clinical variables, IHQ and SNP for each gene. A simplified Bonferroni correction was applied, based on dividing the significant p-value by the number of variables analyzed in the expression and genotypes. Analyses leveraged SPSS v20.0 (IBM, Chicago, IL, USA) and GraphPad Prism 8.0 (La Jolla, CA, USA).
5. Conclusions
With the COVID-19 pandemic now concluded, our study offers valuable insights into the molecular mechanisms underlying fatal outcomes during its course, particularly in the second epidemic wave. We demonstrated that lung tissues from patients who died in this wave exhibit increased expression of inflammasome-associated proteins, especially IL-18, TLR-4, and NLRP3, and that specific genetic polymorphisms in NFKB1 and NOX4 correlate with enhanced inflammasome activation. These results emphasize the role of host genetics and immune dysregulation in determining COVID-19 severity and mortality.
Our findings provide a theoretical foundation for developing targeted biomarkers and therapeutic strategies aimed at inflammasome components, which may have broader applications in managing hyperinflammation in future infectious or inflammatory diseases. The molecular distinctions identified between epidemic waves underline the dynamic interplay between viral evolution and host response, underscoring the importance of ongoing monitoring and personalized interventions beyond the pandemic context.
In summary, enhanced inflammasome activation driven by molecular and genetic factors highlights important aspects of fatal COVID-19 in the second wave, highlighting IL-18 and related genetic markers (rs4648090 NFKB1 and rs317155 NOX4) as molecules of key interest for prognostic and therapeutic consideration in future severe respiratory diseases.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Khreefa Z. Barbier M.T. Koksal A.R. Love G. Del Valle L. Pathogenesis and Mechanisms of SARS-Co V-2 Infection in the Intestine, Liver, and Pancreas Cells 20231226210.3390/cells 1202026236672197 PMC 9856332 · doi ↗ · pubmed ↗
- 2Kartsonaki C. Baillie J.K. Barrio N.G. Baruch J. Beane A. Blumberg L. Bozza F. Broadley T. Burrell A. Carson G. Characteristics and outcomes of an international cohort of 600000 hospitalized patients with COVID-19Int. J. Epidemiol.20235235537610.1093/ije/dyad 01236850054 PMC 10114094 · doi ↗ · pubmed ↗
- 3Baas T. Taubenberger J.K. Chong P.Y. Chui P. Katze M.G. SARS-Co V virus-host interactions and comparative etiologies of acute respiratory distress syndrome as determined by transcriptional and cytokine profiling of formalin-fixed paraffin-embedded tissues J. Interf. Cytokine Res.20062630931710.1089/jir.2006.26.30916689659 PMC 4496958 · doi ↗ · pubmed ↗
- 4Henry B.M. De Oliveira M.H.S. Benoit S. Plebani M. Lippi G. Hematologic, biochemical and immune biomarker abnormalities associated with severe illness and mortality in coronavirus disease 2019 (COVID-19): A meta-analysis Clin. Chem. Lab. Med.2020581021102810.1515/cclm-2020-036932286245 · doi ↗ · pubmed ↗
- 5Lippi G. Favaloro E.J. D-dimer is Associated with Severity of Coronavirus Disease 2019: A Pooled Analysis Thromb. Haemost.202012087687710.1055/s-0040-170965032246450 PMC 7295300 · doi ↗ · pubmed ↗
- 6Kovacs S.B. Miao E.A. Gasdermins: Effectors of Pyroptosis Trends Cell Biol.20172767368410.1016/j.tcb.2017.05.00528619472 PMC 5565696 · doi ↗ · pubmed ↗
- 7Chen M. Wang H. Chen W. Meng G. Regulation of adaptive immunity by the NLRP 3 inflammasome Int. Immunopharmacol.20111154955410.1016/j.intimp.2010.11.02521118671 · doi ↗ · pubmed ↗
- 8Murphy K. Weaver C. Janeway’s Immunobiology 9th ed.Garland Science/Taylor & Francis Group New York, NY, USA 2017