Visualization of peptidoglycan layer isolated from gliding diderm bacteria, Flavobacterium johnsoniae and Myxococcus xanthus, by quick-freeze deep-etch replica electron microscopy
Yuhei O. Tahara, Tâm Mignot, Makoto Miyata

TL;DR
This paper uses electron microscopy to study the peptidoglycan layer structure in gliding bacteria and finds differences in pore size and area.
Contribution
The study provides novel structural insights into peptidoglycan layers of gliding diderm bacteria using high-resolution imaging.
Findings
M. xanthus has larger peptidoglycan pores (51 nm) compared to E. coli and F. johnsoniae.
The pore area in M. xanthus (14.6%) is significantly higher than in other species.
The findings suggest a mechanism for force transmission across the peptidoglycan layer in gliding bacteria.
Abstract
The bacterial peptidoglycan layer plays an important role in protecting the bacteria from turgor pressure, viruses, and predators. However, it also acts as a barrier in transmitting forces generated on the cell membrane to adhesion proteins on the surface during gliding locomotion. In this study, peptidoglycan layers were isolated from two species of gliding diderm, i.e., gram-negative bacteria, and their structures were visualized by quick-freeze deep-etch replica electron microscopy. The horizontal bonding of peptidoglycan did not differ obviously among the three species. However, the diameter of pores in the peptidoglycan layer of M. xanthus and the area of surface pores were 51 nm and 14.6%, respectively, which were significantly larger than those of E. coli (32 nm and 5.8%) and F. johnsoniae (29 nm and 7.0%). Based on this, we discussed the mechanism by which diderm bacteria…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
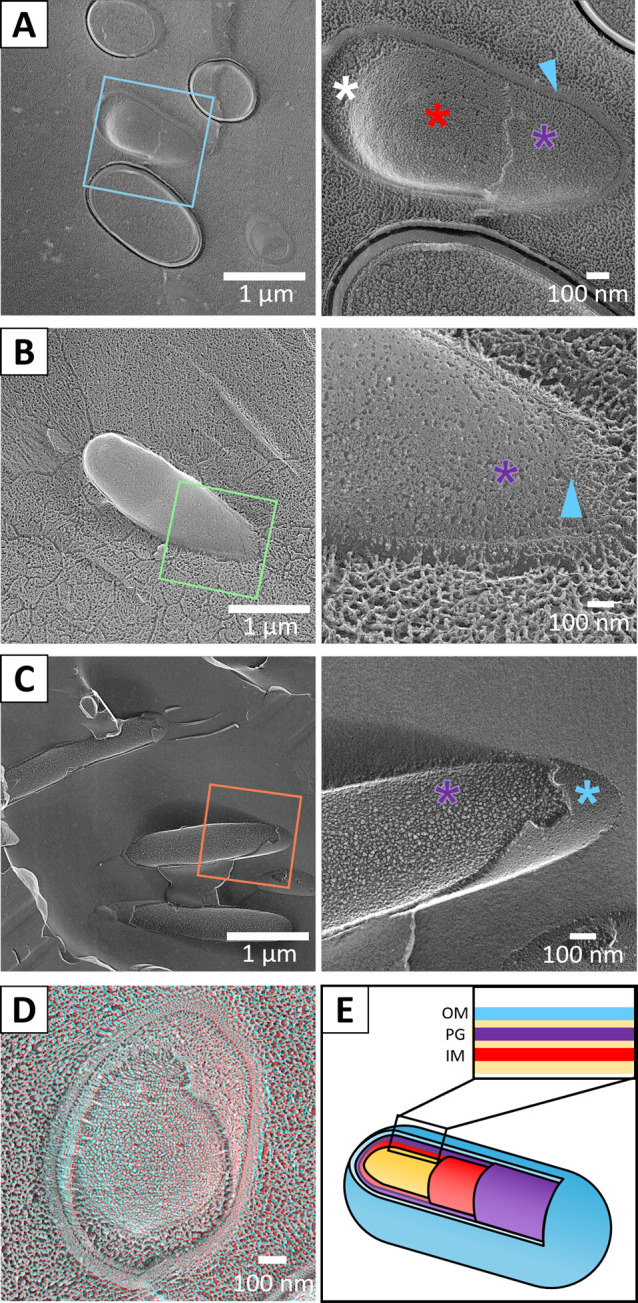 Figure 1
Figure 1 Figure 2
Figure 2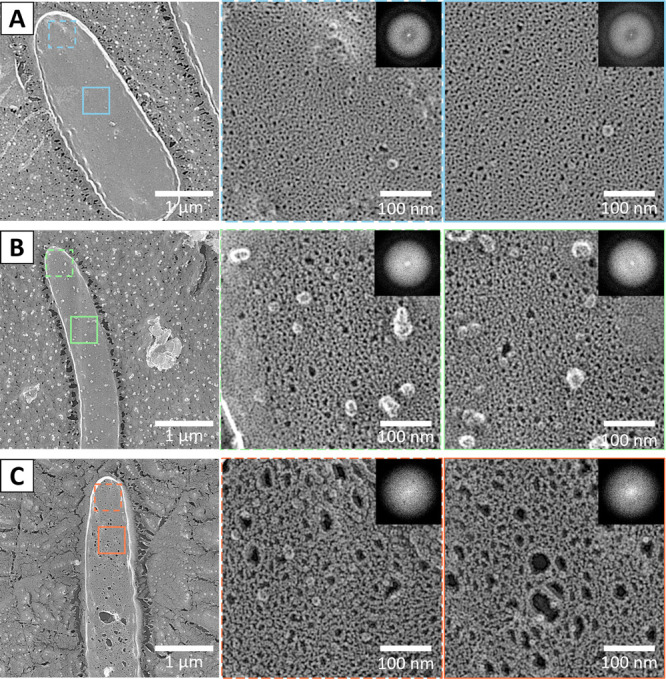 Figure 3
Figure 3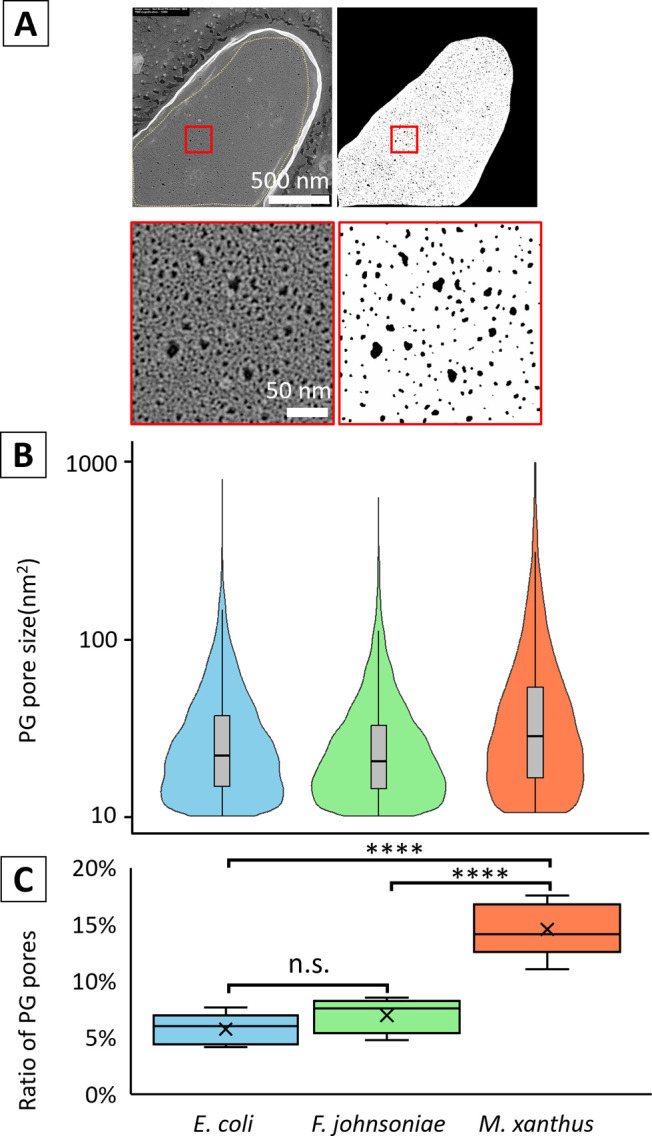 Figure 4
Figure 4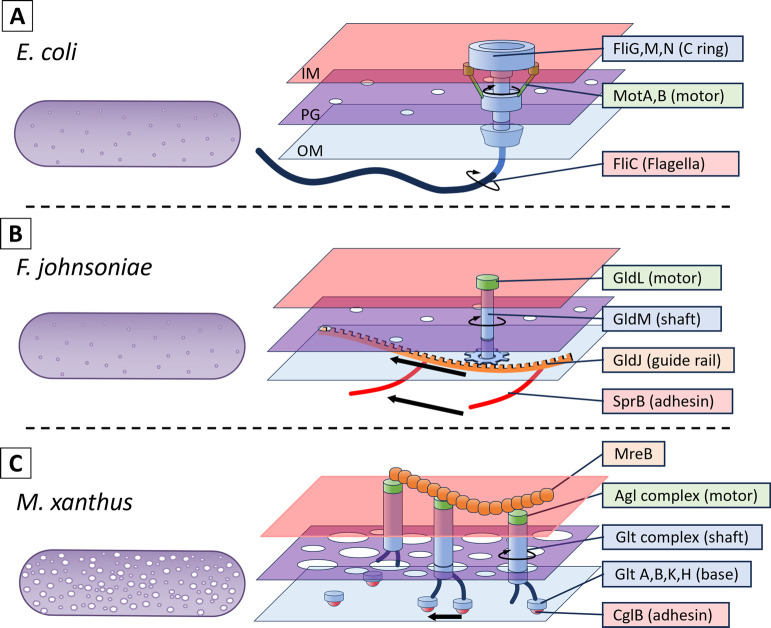 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Community Ecology and Physiology · Genomics and Phylogenetic Studies
Significance
Peptidoglycan is a rigid meshwork that protects bacterial cells from turgor stress, virus, and predators. Bacteria move to get nutrients and escape from predators. The force for motility is generated in the cell membrane and transmitted to the surface across the peptidoglycan layer. In this study, the peptidoglycan structures of two representative gliding Gram-negative bacteria were visualized to understand the transmission mechanism.
Introduction
Peptidoglycan (PG) is a common peripheral structure in bacteria that contributes to shape maintenance and turgor resistance of the cell body [1]. It is a filament composed of two amino sugar units, N-acetylglucosamine (GlcNAc) and N-acetylmuramic acid (MurNAc), crosslinked laterally by peptides to form a meshwork. As PG is a bag-like structure that covers the entire cell, its reorganization is necessary during cell growth and division [2]. The PG mesh can allow macromolecules such as proteins to pass through, however complexes larger than 5 nm are blocked [3].
Many bacteria move to acquire nutrients or new genes or to escape predators or waste products and most bacterial motilities are caused by flagellar rotation or type IV pili (T4P) retraction [4,5]. The proximal ends of flagella and pili are connected to force-generating units, which are embedded in the cell membrane. The force generated by the units in the membrane must be transmitted across the PG and, in the case of Gram-negative bacteria (which are also called “diderm”), further across the outer membrane to the cell surface. As these force-generating units partially penetrate the peptidoglycan and cell membrane, the forces generated inside the cell are directly transmitted to the outside [4,6,7]. However, the gliding motility of Bacteroidota, Gram-negative bacteria, and the T4P-independent gliding motility, “adventurous motility” of Myxococcus require that the force generated in the cell membrane to be transmitted across the PG layer to a substrate such as glass or agar [8–10]. To discuss how forces are transmitted across the PG in these systems, information about the structure of the PG is essential. The information is: Are the PG layers of the board horizontally bonded? How hard is the PG layer? We have previously developed a method to isolate PG layers and reveal their detailed structures, and have visualized PGs of Escherichia coli and Bacillus subtilis, where PGs are isolated by treating cells with SDS and Proteinase K, and then observed by quick-freeze replica electron microscopy (EM) [11–14]. In the quick-freeze replica electron microscopy, samples are frozen within milliseconds using liquid helium. The sample is fractured with a knife, water evaporated, replicated with platinum, and then the replica is observed [15]. This method produces a high contrast image with a resolution that is much better than scanning electron microscopy (SEM). Other advantages of this method are low distortion due to mechanical force and high contrast, unlike AFM and cryo-electron microscopy [2,16]. In this study, we isolated and visualized PGs from nonflagellated E. coli, F. johnsoniae, and M. xanthus, and analyzed and compared their structures in order to understand their motility mechanisms.
Materials and methods
Strains and culture conditions
E. coli (DH5α) was cultivated in Luria-Bertani (LB) medium at 37°C with shaking at 180 rpm. F. johnsoniae (ATCC17061) and M. xanthus (DZ2) were grown in Casitone Yeast Extract (CYE) medium at 30°C with shaking at 150 rpm shaking [17,18]. Cells were harvested at an optical density of around 0.9 at 600 nm.
PG isolation
PG was isolated as previously described. Briefly, 1 mL of culture was collected by centrifugation at an OD_600_ around 0.9. Cells were suspended with 0.5 mL PBS (75 mM sodium phosphate (pH 7.3), 68 mM NaCl). Sodium dodecyl sulphate (SDS) was added to be 0.5% and incubated at 96°C for 3 h. PG was collected by centrifugation at 20,000×g for 20 min, and suspended in 1 mL PBS. This procedure was repeated twice. To the suspension, 10 μL of 10 mg/mL chymotrypsin was added and incubated at 37°C for 2 h. The PG was collected by the centrifugation, suspended in 1 mL water and this process was repeated twice. The final suspension was made with 10 μL water.
Optical microscopy
Cultured cells and purified peptidoglycan of each bacterium were observed in the medium and after purification, respectively. Optical microscopy was performed using a BX50 microscope (Olympus, Tokyo, Japan) equipped with a UPlanSApo 100×1.4 numerical aperture (NA) Ph3 and complementary metal-oxide-semiconductor (CMOS) camera, DMK33UX174 (The Imaging Source Asia Co., Ltd. Taipei, Taiwan) [11,19]. The cell length was measured using Image J ver1.52a, and data analysis was performed using R statistical software (version 4.3.2).
Electron microscopy
Negative staining electron microscopy was performed as previously described [12]. Briefly, samples were placed on EM grids for 1 min, stained with 2% phosphotungstic acid for 1 min, and then dried to remove excess stain.
For quick-freeze deep-etch replica electron microscopy (QFDE-EM) and freeze-fracture electron microscopy, specimens were washed with water and concentrated by centrifugation. A copper block was cooled with liquid helium using a CryoPress, and the samples were slammed it and quickly frozen. The samples were set in a chamber at –180°C of a JFDV freeze-etcher (JEOL, Akishima, Japan). After raising the temperature of the samples to –120°C, their surfaces were freeze-fractured with a knife. For QFDE-EM, samples were etched at –104°C for 15 min and coated by platinum and carbon. For freeze-fracture, they were platinum and carbon coated without etching. The replicas were then floated on original concentration hydrofluoric acid, rinsed with water, washed with a commercial bleach solution containing sodium hypochlorite, rinsed with water, and finally placed on 400-mesh Cu grids. TEM samples were observed with a transmission electron microscope (TEM) using a JEM1010 EM (JEOL) equipped with a FastScan-F214(T) charge coupled device (CCD) camera (TVIPS, Gauting, Germany). The images were measured using Image J ver1.52a, and analysis were performed using R statistical software (version 4.3.2). For width and thickness averaging, we measured positions of typical dimensions from three to four cells or PG sacs.
Results
Visualization of peripheral structures including peptidoglycan layer
To obtain an overview of the peripheral structures, nonflagellated E. coli, F. johnsoniae, and M. xanthus were observed by quick-freeze replica electron microscopy (Fig. 1). For E. coli, two types of images were observed: one in which the outer membrane and periplasmic layer were removed by fracturing, and the other in which the cytoplasm and other cross sections were exposed (Fig. 1A, D, E). We identified each layer, based on the thickness of each layer observed in the cross section and the order of the layers from the outside. The thickness of the periplasmic space was 28±1.8 nm (n=11), thicker at the cell poles, consistent with a previous report [21]. The surface of the PG was porous, which was observed in the area where the outer membrane and periplasmic space were removed. The cell membrane underlying the PG showed general undulation, large holes in some places, and small particles 7.5±0.2 nm (n=30) in diameter [22]. The holes should be structures left after membrane proteins have been pulled out by the fracture, and the particles should be membrane proteins. The fractured images of F. johnsoniae showed that the outer membrane was detached in many cells, exposing the PG. The periplasmic thickness was 26.9±0.7 nm (n=30), in agreement with a previous report (Fig. 1B) [23]. Fine pores similar to those of E. coli were observed on the PG surface below the outer membrane. The fractured images of M. xanthus showed that the outer membrane was detached, exposing the PG (Fig. 1C), as observed in F. johnsoniae. The periplasmic thickness was 28.2±1.0 nm (n=24), in agreement with a previous report [24]. No significant difference in periplasmic thickness was found between the three species (ANOVA p=0.562). The inner and outer membranes as well as the PG layers were too thin to measure their thickness from QFDE-EM. The particles observed on PG layers are likely protein molecules [22].
Isolation of peptidoglycan layers
We isolated PG layers from three bacterial species (Fig. 2). Light microscopy and negative staining EM images showed a clear loss of contrast in the isolated PGs in all three species, indicating that the treatment removed the membrane and cytoplasm. No significant damage was observed in the sac structures visualized by negative staining EM, indicating that sacs composed of PG layers could be isolated in an almost intact state. The results for E. coli were in agreement with a previous report [25]. The widths of cells and isolated PG sacs measured by negative staining EM were 819.0±16.8 nm (n=18) and 1367.6±46.0 nm (n=19), 506.1±16.2 nm (n=13) and 648.0±21.3 nm (n=22), 426±6.4 nm (n=30) and 801.0±58.0 nm (n=12) for E. coli, F. johnsoniae, and M. xanthus, respectively (Fig. 2E). These numbers were in good agreement with those obtained by optical microscopy (Fig. 2E). The width of the cell and PG sac could be explained by the relationship that the contents of the PG sac are removed from and it is flat, because two times of PG sac width agreed with three times of cell diameter, suggesting that the mesh-like PG sac is not significantly stretched when it covers the cell.
Visualization of isolated PG by QFDE-EM
We observed and analyzed PG sacs isolated from each of the three bacterial species using QFDE-EM (Fig. 3). Most of the images obtained by this method also showed a sac shape, and no significant damage was found. The holes were more clearly visualized than by QFDE-EM of the cells or by negative staining of the isolated sac. The structure of E. coli was not significantly different from that of another strain previously published by our group [12–14,26,27]. The holes were present throughout the PG layer sac, with no difference in size between the pole and the center. No directionality with respect to the cell axis was observed in the shape or orientation of the pores, which was also supported by the FFT (Fig. 3A).
No significant damage was observed in the PG sac of F. johnsoniae, and the shape of the pore was clearly observed (Fig. 3B). No orientation of the pore shape or alignment with respect to the cell axis was observed. The PG layer of M. xanthus also retained the sac shape, and the pore shape was clearly observed as in the other two species (Fig. 3C). Therefore, we concluded that the PG layers of two gliding species do not differ obviously regarding lateral bonding from that of E. coli. The Fourier transform of the image did not show any spots or distortions indicating periodicity in the shape of the holes. However, in some places the pores were significantly larger than others, indicating partial damage. Overall, the pores appeared larger than those of the other two species.
Quantification of pore sizes in isolated PG sac
We quantified pore size to objectively characterize the three PG layers. We cut out areas of uniform distribution of holes from the PG sac images, binarized them, and judged the areas of high contrast between 10 nm^2^ and 1000 nm^2^ as ‘holes’ (Fig. 4A). We selected five cells from each species and summed these numbers. The mean area of the holes was 31.8±0.3 nm^2^ for E. coli (n=8666); 29.1±0.3 nm^2^ (n=8500) for F. johnsoniae, 51.2±0.8 nm^2^ (n=8912) for M. xanthus. Welch’s ANOVA, followed by Games-Howell post-hoc tests, was employed to analyze the differences in mean values among the three groups, accounting for the observed heterogeneity of variances. A statistically significant difference among the groups’ mean values was confirmed (p<0.001). Detailed analysis, including the calculation of effect sizes (Cohen’s d), revealed that although the difference between the E. coli and F. johnsoniae groups was statistically significant, its effect size was approximately 0.09, which is considered negligible, suggesting limited practical relevance. In contrast, the M. xanthus group exhibited significantly higher mean values compared to both the E. coli and F. johnsoniae groups. The effect sizes for these differences, while classified as small, were deemed practically meaningful, i.e. larger sized holes were more common in M. xanthus than in the other two species. We compared the percentage of area occupied by the holes in the area (Fig. 4C). The mean values were 5.8%±0.6% for E. coli (n=5), 7.0%±0.7% for F. johnsoniae (n=5), and 14.6%±1% for M. xanthus (n=5). No significant difference between E. coli and F. johnsoniae (Tukey HSD test: p=0.569), while M. xanthus was significantly different from the other two species (p<0.001).
Discussion
The network structure of the F. johnsoniae PG sac was not significantly different from that of E. coli (Figs. 3 and 4), which is consistent with the fact that F. johnsoniae cells appear rigid under the light microscope, as do E. coli cells. Force generation for F. johnsoniae gliding is thought to be caused by proton flow through the inner membrane complex of GldL and GldM [8,28,29]. This complex is structurally similar to the stator of the flagellar motor. F. johnsoniae is observed to rotate in a precise circle around a position to which it adheres and clings to the glass surface, as observed as “tethered” in the bacterial flagellar motor [29–32], In addition, the adhesion protein SprB is anchored to the conveyor belt structure under the OM, and the conveyor belt structure is thought to move in a direction approximately along the cell axis by the proton gradient [23,33]. Based on these observations, a scenario is inferred in which the rotational motion generated by static GldL motors drives the conveyor belt (Fig. 5) [8,23,28]. If we accept this scenario, then there is no inconsistency with the PG structure of F. johnsoniae being common to E. coli, since the only motion transmitted from the cell membrane across the PG layer to the OM surface is the rotation of GldM within a motor complex like a shaft.
In M. xanthus, the area of each pore of the PG sac was larger than in E. coli, as was the percentage of the total pore area on the surface (Figs. 3 and 5). This may be one reason why M. xanthus cells are softer and have a much lower bending modulus than other bacteria [34]. We found a PG sac of M. xanthus with damage that was rarely observed in the other two species. This may also indicate that the PG sac of M. xanthus is softer and more fragile due to its porosity. M. xanthus has two different gliding motility mechanisms (Fig. 5) [35]. One is pili motility, which is observed in many bacteria such as Pseudomonas, Neisseria, and Synechococcus, and is also called “social motility” in M. xanthus. The other is “adventurous motility” characteristic of M. xanthus. This motility is also thought to be driven by a protein closely related to the stator of the flagellar motor, AglR, which generates force through a proton motive force [36–38]. However, unlike flagella and F. johnsoniae, this protein remains membrane-embedded and moves along an unknown track in a nearly axial direction. The fact that the adhesion part moves with the motor means that the lateral movement is transmitted across the PG layer into the extracellular space [9,17,35,39]. If the PG is a rigid sheet [12,16,26,27], forces generated by the motor must be transmitted across the PG to the outside. How this occurs remains to be discovered, but a recent structural model predicts that flexible periplasmic domains of the motor-associated periplasmic Glt proteins link the IM and OM, connecting dynamically to an OM complex containing an adhesin (CglB) [17,40]. The PG holes observed in this study could accommodate these interactions and explain the transfer mechanism.
Conflict of interest
The authors declare that they have no conflict of interest.
Author contributions
YOT did experiments and analyses. TM provided strain, methods and other information of Myxococcus. YOT and MM wrote a draft and discussed the story. All completed the manuscript.
Data availability
The evidence data generated and/or analyzed during the current study are available from the corresponding author on reasonable request. A preliminary version of this work, DOI: https://doi.org/10.1101/2025.01.30.635643, was deposited in the bioRxiv on January 31, 2025.
Acknowledgements
We thank to Daisuke Nakane at the University of Electro-Communications for helpful discussions and Junko Shiomi at Osaka Metropolitan University for technical assistance. This study was supported by Grants-in-aid for scientific research (A) (JP17H01544), a JST CREST grant (JPMJCR19S5) to MM.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Garde, S., Chodisetti, P. K., Reddy, M. Peptidoglycan: Structure, synthesis, and regulation. Eco Sal Plus 9, e ESP-0010-2020 (2021). https://doi.org/10.1128/ecosalplus.ESP-0010-202010.1128/ecosalplus.ESP-0010-2020 PMC 1116857333470191 · doi ↗ · pubmed ↗
- 2Turner, R. D., Mesnage, S., Hobbs, J. K., Foster, S. J. Molecular imaging of glycan chains couples cell-wall polysaccharide architecture to bacterial cell morphology. Nat. Commun. 9, 1263 (2018). https://doi.org/10.1038/s 41467-018-03551-y 29593214 10.1038/s 41467-018-03551-y PMC 5871751 · doi ↗ · pubmed ↗
- 3Demchick, P., Koch, A. L. The permeability of the wall fabric of Escherichia coli and Bacillus subtilis. J. Bacteriol. 178, 768–773 (1996). https://doi.org/10.1128/jb.178.3.768-773.19968550511 10.1128/jb.178.3.768-773.1996 PMC 177723 · doi ↗ · pubmed ↗
- 4Wadhwa, N., Berg, H. C. Bacterial motility: machinery and mechanisms. Nat. Rev. Microbiol. 20, 161–173 (2022). https://doi.org/10.1038/s 41579-021-00626-434548639 10.1038/s 41579-021-00626-4 · doi ↗ · pubmed ↗
- 5Miyata, M., Robinson, R. C., Uyeda, T. Q. P., Fukumori, Y., Fukushima, S. I., Haruta, S., et al. Tree of motility—A proposed history of motility systems in the tree of life. Genes Cells 25, 6–21 (2020). https://doi.org/10.1111/gtc.1273731957229 10.1111/gtc.12737 PMC 7004002 · doi ↗ · pubmed ↗
- 6Minamino, T., Kinoshita, M. Structure, assembly, and function of flagella responsible for bacterial locomotion. Eco Sal Plus 11, eesp 00112023 (2023). https://doi.org/10.1128/ecosalplus.esp-0011-202337260402 10.1128/ecosalplus.esp-0011-2023 PMC 10729930 · doi ↗ · pubmed ↗
- 7Chang, Y. W., Rettberg, L. A., Treuner-Lange, A., Iwasa, J., Sogaard-Andersen, L., Jensen, G. J. Architecture of the type I Va pilus machine. Science 351, aad 2001 (2016). https://doi.org/10.1126/science.aad 200126965631 10.1126/science.aad 2001 PMC 5929464 · doi ↗ · pubmed ↗
- 8Trivedi, A., Gosai, J., Nakane, D., Shrivastava, A. Design principles of the rotary Type 9 secretion system. Front. Microbiol. 13, 845563 (2022). https://doi.org/10.3389/fmicb.2022.84556335620107 10.3389/fmicb.2022.845563 PMC 9127263 · doi ↗ · pubmed ↗