A genome-wide identification and expression analysis of the class III peroxidase gene family in Mangifera indica under abiotic stresses and the MiPRX27 gene regulates oxidative stress
Jia Ran, Junliu Chen, Wang Hao, Hou Jiaxin, Min Zhu, Yeyuan Chen, Jinji Pu, He Zhang

TL;DR
This study identifies 76 peroxidase genes in mango and finds that one gene, MiPRX27, helps plants tolerate oxidative stress.
Contribution
The study provides the first genome-wide analysis of class III peroxidase genes in mango and identifies MiPRX27's role in stress tolerance.
Findings
76 MiPRX genes were identified and unevenly distributed across mango chromosomes.
MiPRX27 overexpression in Arabidopsis improved root growth under oxidative stress.
MiPRX genes show differential expression under oxidative, drought, and salt stress.
Abstract
Class III peroxidases (PRXs) are plant-specific enzymes that play vital roles in various physiological processes. However, the functional roles of mango PRXs under stress conditions remain poorly understood. In this study, we identified 76 MiPRX genes, which are unevenly distributed across the mango chromosomes. RT-qPCR analysis revealed differential expression of most MiPRX genes under oxidative, drought, and salt stress conditions, with MiPRX27 showing a particularly prominent role. Under oxidative stress, heterologous overexpression of MiPRX27 in Arabidopsis enhanced lateral root formation and accelerated root growth, suggesting that MiPRX27 contributes to reducing plant sensitivity to oxidative stress. Overall, this study provides a theoretical foundation for further exploration of MiPRX-mediated mechanisms underlying mango stress tolerance.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
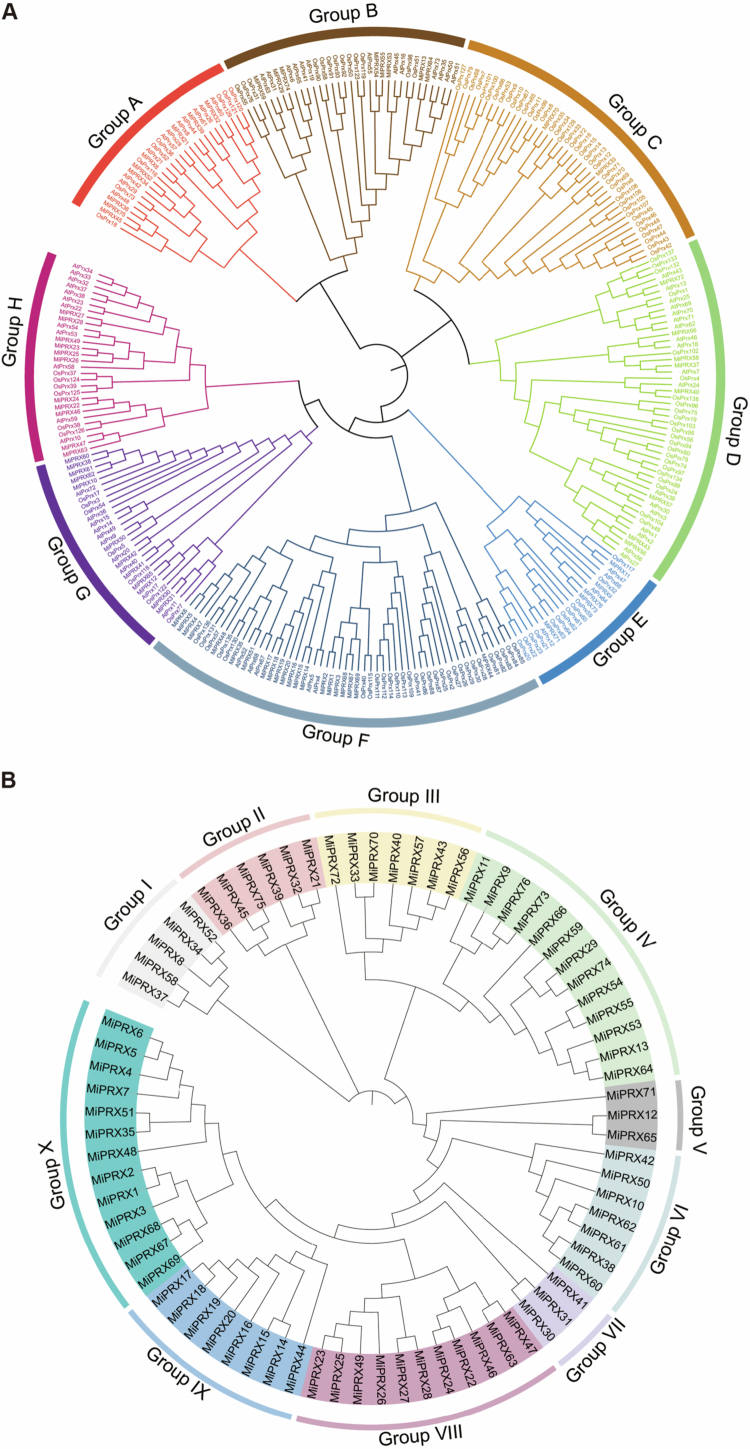 Figure 1
Figure 1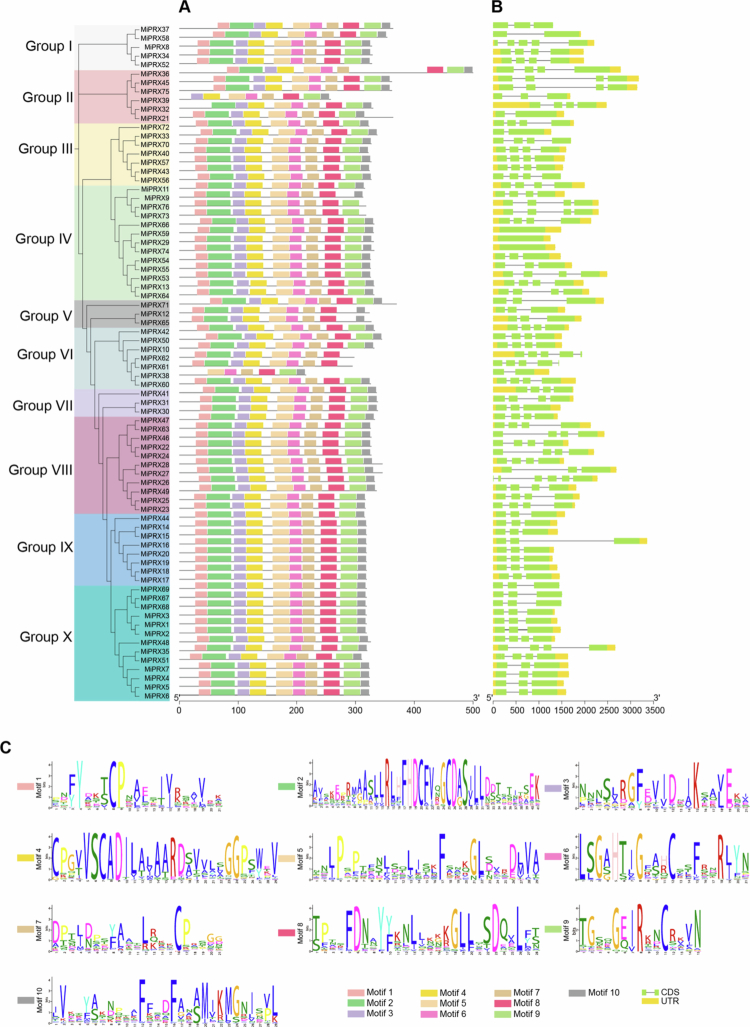 Figure 2
Figure 2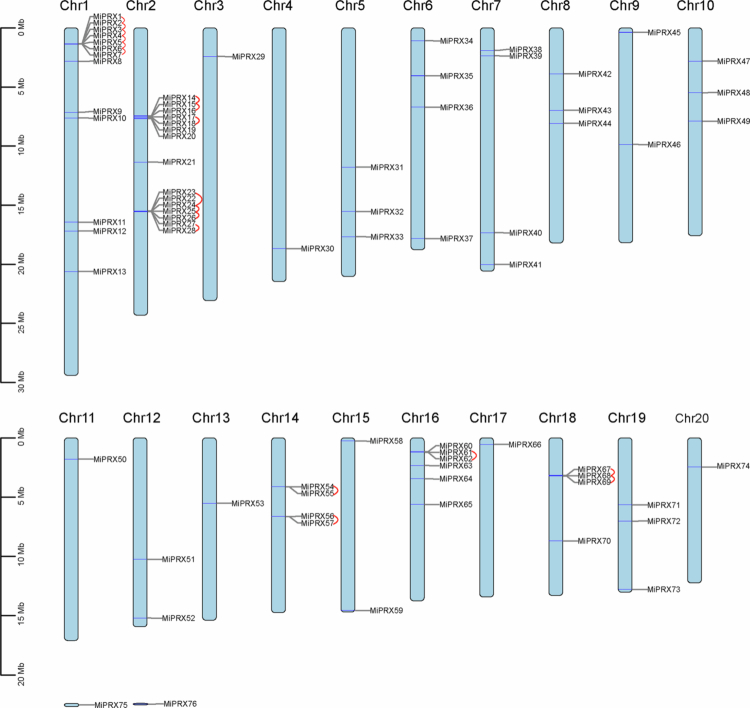 Figure 3
Figure 3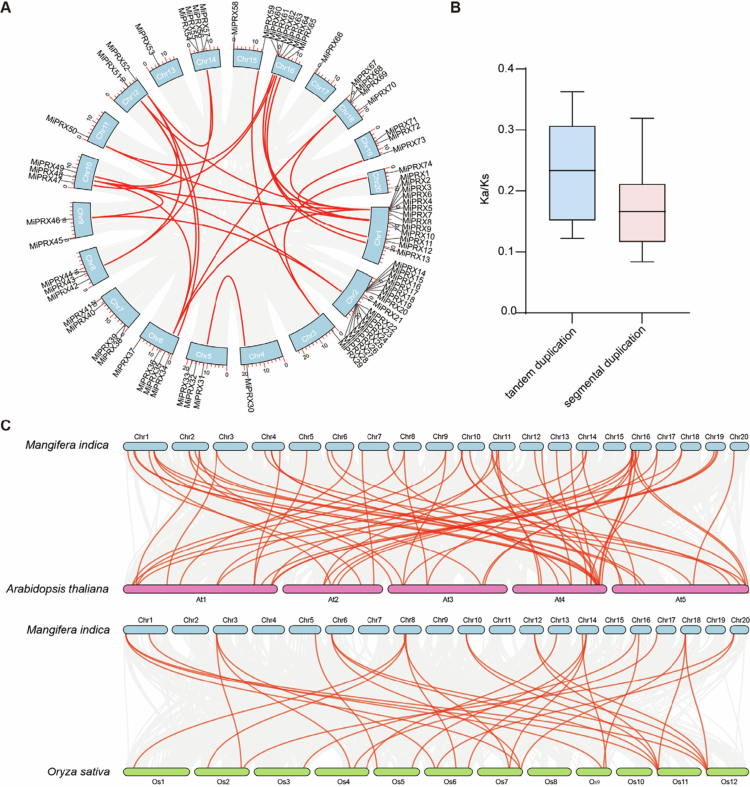 Figure 4
Figure 4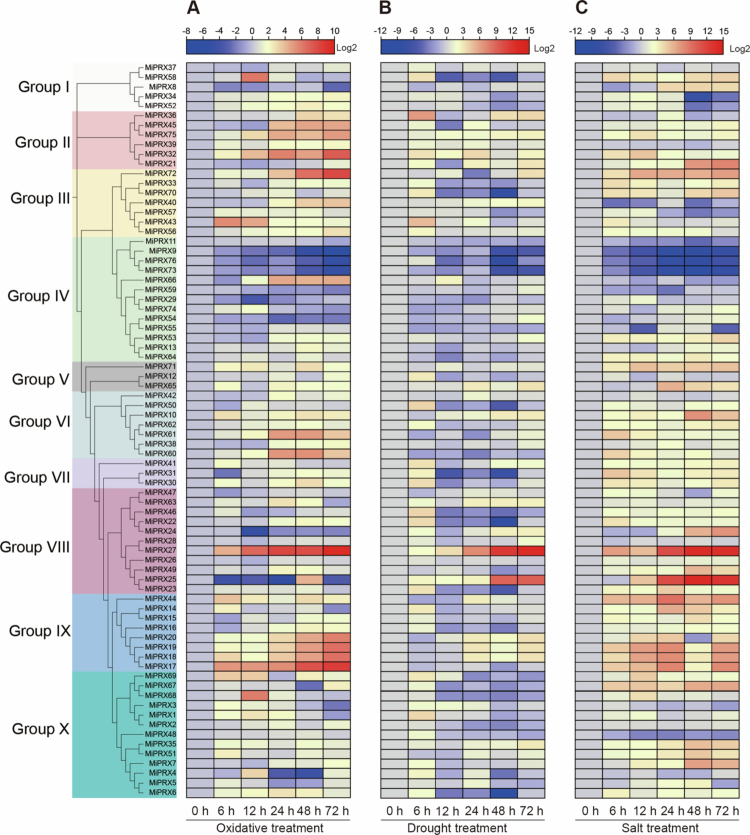 Figure 5
Figure 5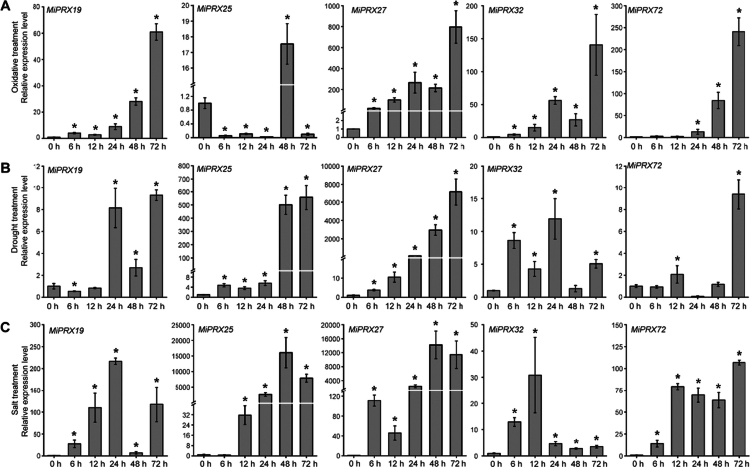 Figure 6
Figure 6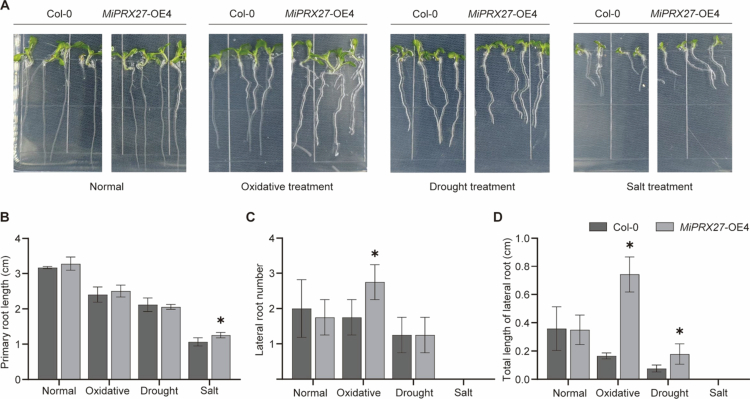 Figure 7
Figure 7- —the Major Science and Technology plan of Hainan Province
- —the Central Public-Interest Scientific Institution Basal Reserch Fund for Chinese Academy of Tropical Agricultural Sciences
- —the China Agriculture Research System of MOF and MARA
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMangiferin and Mango Extracts · Enzyme-mediated dye degradation · Fungal Biology and Applications
Introduction
Peroxidases (EC 1.11.1.X) are enzymes widely distributed across plants, animals, and microorganisms, where they catalyze the oxidation of various substrates using hydrogen peroxide as an electron acceptor.1 Based on the presence or absence of a heme prosthetic group, peroxidases are classified into heme and non-heme types.2 Heme peroxidases are further divided into two major groups: animal and non-animal peroxidases.3 In non-animal peroxidases, the heme group typically consists of protoporphyrin IX coordinated with iron (III). These enzymes are classified into three classes: class I, class II, and class III.3 Among these, Class III peroxidases (also referred to as PRX, PER, POD, or POX) are plant-specific secreted enzymes that play crucial roles in plant growth, development, signal transduction, metabolic regulation, and responses to both biotic and abiotic stresses.2^,^4^,^5
The functions of class III peroxidases in plants have been progressively elucidated. This gene family has been extensively studied in several species, including Arabidopsis,6 rice,7 tobacco,8 guava,9 cucumber,10 cotton,11 and others. Studies have shown that TaPRX-2A and GsPOD40 enhance tolerance to salt and drought stresses by increasing the activity of antioxidant enzymes, such as superoxide dismutase (SOD), peroxidase (PRX), and catalase (CAT), while reducing reactive oxygen species (ROS)-induced oxidative damage.12^,^13 Under salt stress, CRPRX1 was found to improve seed germination in tobacco.14 In response to arsenic (As) exposure, rice OsPRX38 enhances resistance by upregulating the activities of SOD, PRX, and glutathione S-transferase (GST).15 At low temperatures, Arabidopsis AtPRX62 and AtPRX69 promote root hair development.16 PRX genes are also involved in hormone regulation, influencing processes such as seed germination,17 resistance to powdery mildew,18 and other disease-related processes. PRX genes have been extensively associated with enhanced resistance to various plant pathogens.19^,^20
Mango (Mangifera indica L.) is an evergreen woody plant belonging to the Anacardiaceae family, and it stands as one of the most economically significant fruit trees in tropical and subtropical regions. It originated in South Asia and has been cultivated for over 4000 y.21 The fruit is renowned for its high nutritional value,22 with mangiferin in particular exhibiting anti-cancer and anti-diabetic properties.23-25 However, various environmental factors, such as drought and salinity, significantly hinder the growth and development of mango.26^,^27 Salt and drought stress can lead to poor vegetative growth, reduced flowering, and increased fruit drop.28 Therefore, understanding the molecular mechanisms underlying mango response to abiotic stresses is crucial for improving its resilience.
To date, limited research has been conducted on the biological functions of the PRX gene family in mango. In this study, we identified the members of the PRX gene family in the mango genome through bioinformatics analysis. Subsequently, we analyzed gene structures, conserved protein motifs, chromosomal localization, biochemical properties, and phylogenetic relationships. The expression patterns of mango PRX genes under abiotic stresses, including salt, drought, and hydrogen peroxide, were examined using RT-qPCR. Notably, the biological function of MiPRX27 in regulating oxidative stress was further investigated. This study lays the groundwork for further investigation into the roles of PRX genes in mango in response to abiotic stresses.
Materials and methods
Plant material, stress treatment
One-year-old mango plants (Mangifera indica L., cultivar 'Guifei') were used as experimental material. The plants were cultivated in a seedling nursery at the Key Laboratory of Integrated Pest Management for Tropical Crops in Haikou, Hainan Province, China, under controlled growth conditions: a temperature of 28 °C, a light intensity of 5,000 lux, 85% relative humidity, and a 16-hour light/8-hour dark photoperiod.29 Leaves were collected from healthy plants with uniform growth conditions at 0, 6, 12, 24, 48, and 72 h after root irrigation with oxidative stress (300 mmol·L⁻¹ H₂O₂), drought stress (30% PEG6000), or salt stress (300 mmol·L⁻¹ NaCl). Three biological replicates were used for each treatment.28^,^30^,^31 All samples were immediately frozen in liquid nitrogen and stored at −80 °C.
Identification, annotation, and multiple sequence alignment of MiPRX genes
To identify MiPRX gene family members, genomic data for Mangifera indica (CATAS_Mindica_2.1) were retrieved from the NCBI Genome database (https://www.ncbi.nlm.nih.gov/datasets/genome/GCF_011075055.1/). Genomic data for Arabidopsis thaliana and Oryza sativa were obtained from the TAIR10 database (https://www.arabidopsis.org/) and the Ensembl Plants databases (https://plants.ensembl.org/index.html), respectively.
A total of 138 PRX protein sequences from Oryza sativa and 73 from Arabidopsis thaliana were used as reference sequences.6^,^7 Homology-based searches were performed across the entire mango genome using BLAST. Additionally, the conserved peroxidase domain (PF00141) was utilized to construct a Hidden Markov Model (HMM) for identifying candidate PRX genes within the mango genome. The Conserved Domain Database (CDD; https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) was used to validate the presence of the PRX domain. Candidate genes identified by both BLAST and HMM searches were intersected to cross-check and remove redundant sequences, thereby confirming the members of the MiPRX gene family. Redundant sequences were removed after comparison, and the remaining non-redundant sequences were considered potential MiPRX proteins. The physicochemical properties of the MiPRX proteins, including molecular weight (MW) and isoelectric point (pI), were predicted using the ProtParam tool (https://web.expasy.org/protparam/). Subcellular localization was predicted using Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/).[32](#cit0032) A phylogenetic tree was constructed using MEGA 11 (https://www.megasoftware.net/) with the neighbor-joining method and 1000 bootstrap replicates.33
The amino acid sequences of the MiPRX gene family were aligned using ClustalW (https://www.genome.jp/tools-bin/clustalw), based on the identified MiPRX genes. The alignment results were then visualized with ESPript 3 (https://espript.ibcp.fr/ESPript/ESPript/index.php) for further analysis.
Analysis of conserved motifs, gene structure, and cis-acting elements of the MiPRX promoter
The conserved motifs of the MiPRXs were identified using the MEME online platform (https://meme-suite.org/meme/tools/meme),[34](#cit0034) with a maximum of 10 motifs specified. The domain architecture of MiPRXs was further analyzed using the Batch CD-Search tool in the Conserved Domain Database (CDD) (https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi), applying an E-value threshold of 0.01 and a maximum hit limit of 500. The chromosomal locations of MiPRX genes were determined using the GFF annotation file of the mango genome, and TBtools was used to visualize gene structures and conserved motifs.35 To explore the cis-acting elements in the promoter region of MiPRX genes, a 2000 bp sequence upstream of each MiPRX coding sequence (CDS) was extracted from the mango genome and defined as the promoter region. These sequences were submitted to the PlantCARE database (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/) for cis-element prediction.36 The number, type, and distribution of the predicted elements were visualized using TBtools.35
Chromosomal localization and duplication pattern analyzes
The chromosomal locations of MiPRX genes were obtained from the annotated mango genome. Gene duplication events were identified and visualized using TBtools, which was also employed to assign names to the MiPRXs based on their chromosomal positions.35 To explore the evolutionary relationships of Class III PRXs among M. indica, A. thaliana, and O. sativa, MCScanX was used.35
RNA extraction and RT-qPCR
Total RNA was extracted using the RNA prep Pure Polysaccharide Polyphenol Plant Total RNA Extraction Kit (TIANGEN, Beijing, China), according to the manufacturer's instructions. First-strand cDNA synthesis was performed using the FastKing gDNA Dispelling RT SuperMix (TIANGEN, Beijing, China). Quantitative real-time PCR (RT-qPCR) was conducted with the SGExcel UltraSYBR Mixture (Sangon Biotech, Shanghai, China). Gene-specific primers were designed based on the coding sequences (CDS) of MiPRXs using Primer3Plus (https://www.primer3plus.com/), with MiActin used as the reference gene (Table S1). Gene expression was quantified using a qTOWER³ real-time PCR system (Analytik Jena GmbH, Göttingen, Germany).
Construction of the expression vector and genetic transformation of Arabidopsis
The full-length coding region of MiPRX27 was amplified by PCR using Pfu DNA polymerase and gene-specific primers (Table S1), then cloned into the pEGAD overexpression vector via homologous recombination. The resulting recombinant plasmid was designated 35S::MiPRX27. Both the recombinant and empty control vectors were introduced into Agrobacterium tumefaciens strain GV3101 using the heat shock method.37
Transgenic Arabidopsis and stress treatment
Arabidopsis ecotype Col−0 was used as the wild-type control. Overexpression lines were generated via the floral dip method using Agrobacterium tumefaciens GV3101 harboring the 35S:MiPRX27 recombinant plasmid.38 Transgenic plants were selected on medium containing 20 mg/L glufosinate ammonium and further confirmed by PCR. Sterilized seeds of both transgenic and Col−0 plants were sown on 1/2MS medium supplemented with oxidative stress (0.5 mmol·L⁻¹ H₂O₂), drought stress (200 mmol·L⁻¹ mannitol), or salt stress (150 mmol·L⁻¹ NaCl), and grown for 7 d.39 Primary root length, number of lateral roots, and total lateral root length were measured using ImageJ software to assess the tolerance of transgenic plants to oxidative, drought, and salt stresses.40-42 All experiments were performed with three independent biological replicates.
Statistical analysis
All data were analyzed using a Student's t-test in IBM SPSS Statistics 26. P-values less than 0.05 were considered statistically significant and are represented by ‘*’. The relative mRNA levels for each gene were calculated using the 2^−ΔΔCT^ method.29 RT-qPCR experiments were performed at least three times under identical conditions. Results were visualized using GraphPad Prism 9 and TBtools.29^,^35 The experiment was conducted using three independent biological replicates.
Results
Genome-wide identification, phylogenetic analysis of MiPRX proteins
Using both BLAST and HMM methods, we identified 76 MiPRX proteins from the mango genome. Based on their chromosomal locations, these MiPRX family members were named MiPRX1 to MiPRX76 (Table S2). The MiPRX proteins varied significantly in length, ranging from 216 amino acids (MiPRX38) to 500 amino acids (MiPRX36), with molecular weights (Mw) ranging from 24.01 to 53.56 kDa. Additionally, the isoelectric points (pI) of the MiPRX proteins varied between 4.25 (MiPRX23) and 9.64 (MiPRX38). Predicted subcellular localization indicated that MiPRX proteins were primarily localized in the cytoplasm, with some also detected in vesicles (Table S2). A phylogenetic tree was constructed using the neighbor-joining (NJ) method to analyze the evolutionary relationships of PRX amino acid sequences from M. indica, A. thaliana, and O. sativa. The tree also facilitated the comparison of monocotyledonous and dicotyledonous plants. Eight Groups (A-H) were identified in the phylogenetic tree, as shown in Figure 1A. Group F contained the largest number of MiPRX proteins (21 members), followed by Group G (12 members). In contrast, Group C had only two members, representing 4.76% of the total in this group. To further investigate the evolutionary relationships among MiPRX members, a separate phylogenetic tree based solely on MiPRX amino acid sequences was also constructed (Figure 1B). Multiple sequence alignments of 76 MiPRX sequences were performed using ClustalW and ESPript 3 (Figure S1). Ten groups (Group I–X) were defined based on the 76 MiPRX members in mango. Notably, the distribution of MiPRX proteins across the groups varied, with group sizes ranging from 3 to 13 members per group. Group IV and Group X contained the highest number of members, with 13 in each, while Group V contained only three members, accounting for 3.9% of the total.
Analysis of conserved motifs and gene structure of MiPRXs
In this study, we explored the evolutionary conservation of MiPRX genes by analyzing their structural characteristics. A total of at least ten motifs were identified in the amino acid sequences of MiPRXs using the MEME database, as shown in Figure 2. Based on these motifs, MiPRXs were clustered into groups in the evolutionary tree, suggesting that these proteins may share functional similarities (Figure 2A). At least six conserved motifs are present in all MiPRX proteins. For example, motifs 5, 6, 7, 8, 9, and 10 are found in MiPRX38. Interestingly, motifs 5, 6, 7, and 8 are conserved across all MiPRX proteins. The amino acid composition of these conserved motifs is highly similar. Over 86.84% of all MiPRX proteins from mango contain all ten motifs.
We also performed exon-intron analysis to explore the structural diversity of MiPRX genes (Figure 2B). The results revealed that MiPRX genes contain between 0 and 5 introns. Notably, MiPRX8, MiPRX61, and MiPRX62 each contained four introns, while MiPRX29, MiPRX59, and MiPRX74 lack introns. All members of Group IX possess two introns and three exons. These findings indicate that genes within the same group tend to exhibit similar structural features.
The conserved motifs of MiPRXs were identified using the MEME tool, and the specific amino acid sequences of 10 conserved motifs were obtained (Figure 2C, Table S3). The cis-acting elements of MiPRXs were identified using PlantCARE and visualized using TBtools. Cis-acting elements can be divided into eight categories based on their function (Figure S2).
MiPRXs phylogenetic analysis. (A) Phylogenetic tree of PRX proteins from O. sativa, A. thaliana, and M. indica. Peroxidases from M. indica, A. thaliana, and O. sativa are denoted as MiPRX, AtPRX, and OsPRX, respectively. (B) Phylogenetic analysis of mango PRXs. Different colors represent different groups.
Conserved protein motifs and gene structure of MiPRXs. (A) Conserved motifs of MiPRXs. Different colored boxes represent conserved motifs, while gray lines indicate non-conserved motifs. (B) Exon-intron structures of the MiPRX genes. Green squares represent exons, yellow squares indicate untranslated regions (UTRs), and gray lines represent introns. (C) The amino acid sequences of the motifs. Larger letters indicate a higher frequency of amino acids at that position.
Chromosomal distribution of the MiPRX genes
Statistics on the distribution of the mango PRX gene family showed that 76 MiPRX genes are distributed across the chromosomes of mango. Each chromosome contains between 1 and 15 MiPRX genes, with approximately one-third of these genes located on Chr1 and Chr2 (Figure 3). On Chr3, Chr 4, Chr 11, Chr 13, and Chr 20, only one MiPRX gene is present. Additionally, MiPRX75 and MiPRX76 are located in unassembled chromosomal regions.
Chromosomal distribution and tandem duplications of MiPRXs. The MiPRX genes, from MiPRX1 to MiPRX76, were renamed based on their chromosomal positions, with two genes remaining unassigned to any chromosome. Chromosomal localization of MiPRXs was carried out using the mango genome database. Tandem duplications of two pairs are indicated by red lines.
Syntenic and selection pressure analysis of the MiPRX gene family
Gene family expansion is primarily driven by gene duplication. To investigate the expansion mechanism of MiPRXs, we performed collinearity analysis (Figure 4, Table S4). Among the 43 paralogous pairs of MiPRXs, 18 pairs exhibited tandem duplication events, while 25 pairs showed segmental duplication ([Figures 3 and 4A](#f0003 f0004)). These results indicate that both tandem and segmental duplications contribute to the expansion of MiPRXs. Additionally, the Ka/Ks ratio for both tandem and segmental duplications is less than 1, suggesting negative selection (Figure 4B, Table S4). This implies that the evolution of class III peroxidases is largely influenced by purifying selection, with tandem duplications playing a more significant role.
Syntenic relationship of MiPRX genes. (A) MiPRX genes duplication analysis. Gray lines represent duplicated gene pairs in mango, while red lines highlight the segmental duplication events of MiPRX genes. (B) Distribution of Ka/Ks values for MiPRXs duplication events. (C) PRX genes synteny analysis between O. sativa, A. thaliana, and M. indica. Chromosomes of M. indica, A. thaliana, and O. sativa are distinguished by different colors. Syntenic PRX gene pairs are highlighted by red lines, and collinear blocks are represented by gray lines.
To better understand the evolutionary relationships among PRX genes in plants, we analyzed the collinearity between M. indica and A. thaliana, as well as between M. indica and O. sativa (Figure 4C). A total of 63 homologous gene pairs were identified between M. indica and A. thaliana, and 32 pairs between M. indica and O. sativa (Figure 4C, Table S5). In these gene pairs, a single MiPRX may correspond to multiple genes in A. thaliana and O. sativa. For example, MiPRX29 was found to homologous to AT1G24110.1, AT5G40150.1, Os02t0741200-01, Os04t0465100-01, and Os06t0237600-00. These collinearity results suggest that the evolutionary relationship between M. indica and A. thaliana is closer than that between M. indica and O. sativa.
MiPRX gene expression patterns under oxidative, drought, and salt treatments
To further elucidate the potential biological functions of MiPRXs under abiotic stresses, we analyzed their expression patterns in response to oxidative, drought, and salt stresses using RT-qPCR (Figure 5). The results show that the expression levels of MiPRXs vary under different stress conditions, indicating their diverse roles in stress responses. Expression of 76 MiPRXs was evaluated under oxidative stress (Figure 5A, Table S6). Notably, MiPRX6, −10, −17, −18, −19, −20, −27, −32, −34, −35, −39, −43, −44, −52, −56, −57, −60, −61, −71, −72, and −75 exhibited high expression levels at various time points. In contrast, MiPRX7, −22, −31, −36, −38, −40, −42, −45, −36, and −53 showed increased expression after 12 h (Figure 5A). Specifically, MiPRX4 and MiPRX68 reached their peak expression at 12 h, while MiPRX25 and MiPRX43 exhibited the highest expression at 6 and 48 h, respectively. Additionally, several MiPRXs, including MiPRX9, MiPRX73, and MiPRX76, were down-regulated under stress conditions.
Following drought stress treatment, 13.1% of MiPRX members exhibited increased expression levels (Figure 5B, Table S7), with MiPRX25 and MiPRX27 showing the highest expression. In contrast, MiPRX9, −11, −73, and −76 were down-regulated under drought conditions. Notably, except for MiPRX66, which showed induced expression at 12 h, all genes in Group IV exhibited decreased expression at one or more time points. Additionally, the expression of MiPRX18 and MiPRX19 peaked at 6 h and then gradually reduced. MiPRX9, −11, −73, and −76 showed reduced transcript levels, indicating their sensitivity to drought stress.
The expression levels of Group IV members (MiPRX9, −11, −59, −66, −73, −76) were down-regulated at most time points following salt stress treatment (Figure 5 C, Table S8). MiPRX25, −27, and −72 were rapidly activated and maintained at high expression levels throughout the treatment. In contrast, MiPRX17, −18, and −19 exhibited a pattern of initial increase followed by a decrease, peaking at 36 h and reaching their lowest levels at 48 h.
The expression patterns of MiPRXs under various stress conditions were analyzed using RT-qPCR. We evaluated the MiPRXs expression profiles in response to oxidative treatment (A), drought treatment (B), and salt treatment (C). Fold changes were calculated using Log 2-based RT-qPCR, and a heat map was generated using TBtools. Relative expression was considered down-regulated if it was less than 0.5-fold of the control and up-regulated if it was 1.5-fold higher than the control. Different colors visually represent the relative expression levels.
The RT-qPCR analysis of MiPRXs under abiotic stresses revealed that MiPRX19, −25, −27, −32, and −72 respond positively to oxidative, drought, and salt treatments. Under oxidative stress, MiPRX27 expression levels increased over time (Figure 6A). Conversely, the expression levels of MiPRX25 and MiPRX27 increased following exposure to drought and salt stresses (Figure 6B–C). Under all three abiotic stress conditions, the expression levels of MiPRX27 consistently increased over time. These results suggest that MiPRX27 may play a significant role in the stress resistance mechanisms of mango.
The relative expression levels of MiPRX19, −25, −27, −32, and −72 in response to oxidative, drought, and salt treatments were assessed using RT-qPCR. Figures (A-C) show the expression levels under oxidative (A), drought (B), and salt treatment (C). The 2−∆∆CT method was used to calculate expression levels (; p-value ≤ 0.05). Three biological and technical replicates were used to calculate the mean and standard deviation (SD).*
Heterologous overexpression of MiPRX27 affects sensitivity to oxidative, drought, and salt stress in Arabidopsis thaliana
To further validate the role of MiPRX27 in mango resistance to abiotic stress, we stably overexpressed the MiPRX27 gene in Arabidopsis thaliana using Agrobacterium-mediated stable genetic transformation. Transgenic lines were screened using Sanger sequencing to identify a MiPRX27-OE4 line without nucleotide mutations. To assess the function of MiPRX27 in abiotic stress, we measured the primary root length, lateral root number, and total lateral root length of Col−0 and MiPRX27-OE overexpression lines under oxidative, drought, and salt stress treatments, respectively.
As shown in Figure 7, under normal 1/2MS culture conditions, there were no significant differences in primary root length, lateral root number, or total lateral root length between the two lines (Figure 7A). However, under oxidative stress, the lateral root number and total root length of MiPRX27-OE were significantly greater than those of Col-0 (Figure 7C). Under drought treatment, MiPRX27-OE exhibited a greater total lateral root length compared to Col-0 (Figure 7D). In response to salt treatment, the primary root length of MiPRX27-OE was significantly increased relative to Col-0 (Figure 7B).
Growth of 7-d-old Arabidopsis seedlings in Col−0, MiPRX27 overexpression lines was observed under different stresses in 1/2 MS medium supplemented with oxidative, drought, and salt treatments. Phenotypes of Arabidopsis seedlings of the Col−0 and MiPRX27-OE4 overexpression lines under different stresses (A), primary root length (B), lateral root number (C), Total length of lateral root (D) (; p-value ≤ 0.05, Student's t-test).*
Discussion
Class III peroxidases (PRXs) play a crucial role in regulating various physiological processes, including plant growth and development. They play a key role in responding to both biotic and abiotic stresses by controlling reactive oxygen species (ROS) and other oxidative molecules.1^,^3 Under abiotic stress, PRX enzymes can scavenge reactive oxygen species (ROS), thereby mitigating associated damage.42 To date, the PRX gene family has been extensively studied at the genome-wide level in Arabidopsis, rice, tobacco,8 pineapple,43 and cassava.5 However, limited research has been conducted on the PRX gene family in mango. Following the completion of the mango genome sequencing, the PRX gene family has been analyzed at the genomic level.44 In this study, we identified 76 MiPRX genes using mango genomic data, and analyzed their phylogenetic tree, collinearity, gene structures, conserved motifs, and expression patterns. This study lays the foundation for understanding the role of MiPRXs in abiotic stresses.
This study identified 76 MiPRX proteins, each containing 10 highly conserved motifs. In addition, variations in the number and type of conserved motifs were observed across the 76 MiPRX proteins. The diversity of these motifs may be implicated in the biological functions of MiPRXs. Variations in gene structures are critical for the evolution of gene families.45 Gene structural analysis revealed that the 76 MiPRX genes differ in the number of exons and introns, with clear structural similarities observed within each phylogenetic group. Additionally, some MiPRX genes lack introns, likely due to specific evolutionary mechanisms such as exon or intron gain or loss.46
The MiPRX members are unevenly distributed on 20 chromosomes, a pattern consistent with the distribution of PRX genes on chromosomes in cassava,5 birch, castor bean,47 sugarcane,48 and ginger.45 Gene family expansion is typically driven by segmental duplications, tandem duplications, and genome-wide duplications.49 Analysis of MiPRXs duplication events revealed that 58.1% of the duplicated genes resulted from segmental duplications, suggesting that segmental duplications are the primary mechanism driving MiPRXs evolution. In contrast, ShPRXs in sugarcane are predominantly expanded through tandem duplications. Ka/Ks analysis further indicates that the evolution of the MiPRX family is largely shaped by negative selection, a pattern consistent with findings in soybean,13 tobacco,8 and rice.7 Homology analysis of PRX genes revealed that mango shares a higher number of homologous genes and exhibits closer genetic relationships with Arabidopsis thaliana.
Previous studies have demonstrated that peroxidase (PRXs) genes play a crucial role in regulating both abiotic stresses and environmental stress resistance. These studies have examined the differential expression patterns of PRXs in response to salt, drought, heavy metals, oxidative stress, in species such as cucumber,10 sugarcane,48 and Arabidopsis.50 Under oxidative stress, transgenic soybean GsPRX9 exhibited enhanced antioxidant capacity, primarily by increasing the activities of PRX, SOD, and GST enzymes and reducing intracellular H_2_O_2_ levels.51 Under salt stress, PRX scavenges ROS to alleviate the detrimental effects of salinity. In tobacco, NtPRX63 upregulates the expression of antioxidant-related genes, enhancing antioxidant enzyme activity and ROS scavenging ability, thereby improving salt stress tolerance.52 Similarly, salt stress significantly induced the activity of the sweet potato IbPRX17, reducing intracellular ROS levels and mitigating salt-induced damage.53 During drought stress, the ZmPRX26 gene in maize plays a crucial role. The expression patterns of MiPRXs in response to H_2_O_2_, drought, and salt treatments showed differential transcriptional regulation in response to these stresses. Notably, the transcriptional level of MiPRX27 was significantly activated, indicating that MiPRX27 may play a key role in stress responses.
To investigate the response of MiPRX27 to abiotic stress, we introduced MiPRX27 into Arabidopsis thaliana. Seeds were cultured under oxidative stress (0.5 mmol·L⁻¹ H₂O₂), drought stress (200 mmol·L⁻¹ mannitol), and salt stress (150 mmol·L⁻¹ NaCl). Under normal 1/2 MS medium, no significant differences in taproot and lateral root growth were observed between wild-type and MiPRX27 overexpressing lines. However, under oxidative stress, the number and length of lateral roots in the overexpressing lines were significantly greater than those in the wild-type. These results suggest that MiPRX27 overexpression reduces the sensitivity of Arabidopsis lateral roots to oxidative stress (Figure 7), highlighting the specific role of MiPRX27 in regulating plant responses to oxidative stress.
Conclusion
This study provides a comprehensive genome-wide analysis of the MiPRX gene family in mango, identifying a total of 76 MiPRX members. Phylogenetic and structural analyzes revealed conserved motifs and key evolutionary patterns within the family. Additionally, MiPRX27 was found to be upregulated under oxidative, drought, and salt stress conditions, suggesting its potential role in enhancing tolerance to abiotic stresses. Overexpression of MiPRX27 in Arabidopsis was achieved using Agrobacterium-mediated floral dip transformation, and the transgenic lines were assessed for stress resistance under various conditions. Notably, under oxidative stress, MiPRX27 overexpression significantly increased the number and length of lateral roots in Arabidopsis. This study provides valuable insights into the functional roles of PRX genes and offers a rich genetic resource for developing mango varieties with improved stress resistance.
Supplementary Material
Supplementary material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kidwai M, Ahmad IZ, Chakrabarty D. Class III peroxidase: an indispensable enzyme for biotic/abiotic stress tolerance and a potent candidate for crop improvement. Plant Cell Rep. 2020;39:1381–1393. doi: 10.1007/s 00299-020-02588-y.32886139 · doi ↗ · pubmed ↗
- 2Hiraga S, Sasaki K, Ito H, Ohashi Y, Matsui H. A large family of class III plant peroxidases. Plant Cell Physiol. 2001;42:462–468. doi: 10.1093/pcp/pce 061.11382811 · doi ↗ · pubmed ↗
- 3Freitas CDT, Costa JH, Germano TA, Rocha RD, Ramos M, Bezerra LP. Class III plant peroxidases: from classification to physiological functions. Int J Biiol Macromol. 2024;263:130306. doi: 10.1016/j.ijbiomac.2024.130306.38387641 · doi ↗ · pubmed ↗
- 4Almagro L, Ros LVG, Belchi-Navarro S, Bru R, Barceló AR, Pedreño MA. Class III peroxidases in plant defence reactions. J Exp Bot. 2009;60:377–390. doi: 10.1093/jxb/ern 277.19073963 · doi ↗ · pubmed ↗
- 5Wu CL, Ding XP, Ding ZH, Tie WW, Yan Y, Wang Y, Yang H, Hu W. The class III peroxidase (POD) gene family in Cassava: identification, phylogeny, duplication, and expression. Int J Mol Sci. 2019;20:2730. doi: 10.3390/ijms 20112730.31163686 PMC 6600411 · doi ↗ · pubmed ↗
- 6Tognolli M, Penel C, Greppin H, Simon P. Analysis and expression of the class III peroxidase large gene family in Arabidopsis thaliana. Gene. 2002;288:129–138. doi: 10.1016/S 0378-1119(02)00465-1.12034502 · doi ↗ · pubmed ↗
- 7Passardi F, Longet D, Penel C, Dunand C. The class III peroxidase multigenic in land plants family in rice and its evolution. Phytochem. 2004;65:1879–1893. doi: 10.1016/j.phytochem.2004.06.023.15279994 · doi ↗ · pubmed ↗
- 8Cheng LT, Ma LX, Meng LJ, Shang HH, Cao PJ, Jin JJ. Genome-wide identification and analysis of the class III peroxidase gene family in tobacco (Nicotiana tabacum). Front Genet. 2022;13:916867. 10.3389/fgene.2022.916867.35769995 PMC 9234461 · doi ↗ · pubmed ↗