New alleles of D-2-hydroxyglutarate dehydrogenase enable studies of oncometabolite function in Drosophila melanogaster
Madhulika Rai, Prince Okah, Shefali A Shefali, Alexander J Fitt, Michael Z Shen, Mandkhai Molomjamts, Robert Pepin, Travis Nemkov, Angelo D'Alessandro, Jason M Tennessen

TL;DR
Researchers created new fruit fly mutants to study how the oncometabolite D-2HG affects development and metabolism.
Contribution
The paper introduces two novel D2hgdh loss-of-function alleles in Drosophila for studying D-2HG function.
Findings
The D2hgdh mutants show elevated D-2HG levels and distinct developmental and metabolomic phenotypes.
The new alleles provide a model to study context-dependent effects of D-2HG in human disease and normal physiology.
Abstract
D-2-hydroxyglutarate (D-2HG) is a potent oncometabolite capable of disrupting chromatin architecture, altering metabolism, and promoting cellular dedifferentiation. As a result, ectopic D-2HG accumulation induces neurometabolic disorders and promotes progression of multiple cancers. However, the disease-associated effects of ectopic D-2HG accumulation are dependent on genetic context. Specifically, neomorphic mutations in the mammalian genes Isocitrate dehydrogenase 1 (IDH1) and IDH2 result in the production of enzymes that inappropriately generate D-2HG from α-ketoglutarate (αKG). Within this genetic background, D-2HG acts as an oncometabolite and is associated with multiple cancers, including several diffuse gliomas. In contrast, loss-of-function mutations in the gene D-2-hydroxyglutarate dehydrogenase (D2hgdh) render cells unable to degrade D-2HG, resulting in excessive buildup of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
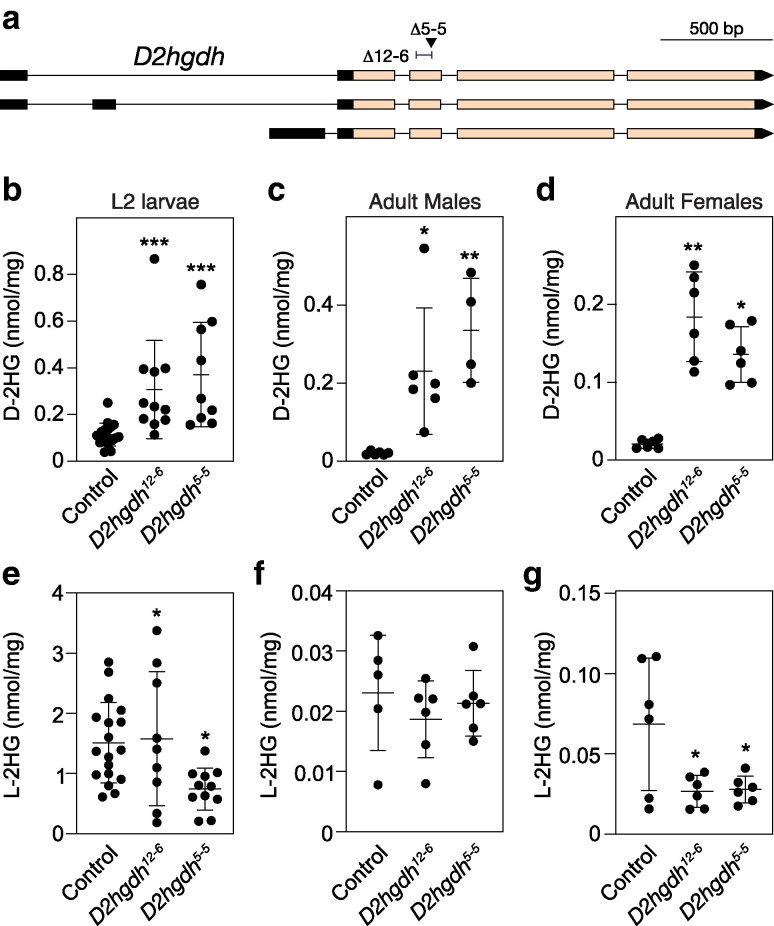 Fig. 1
Fig. 1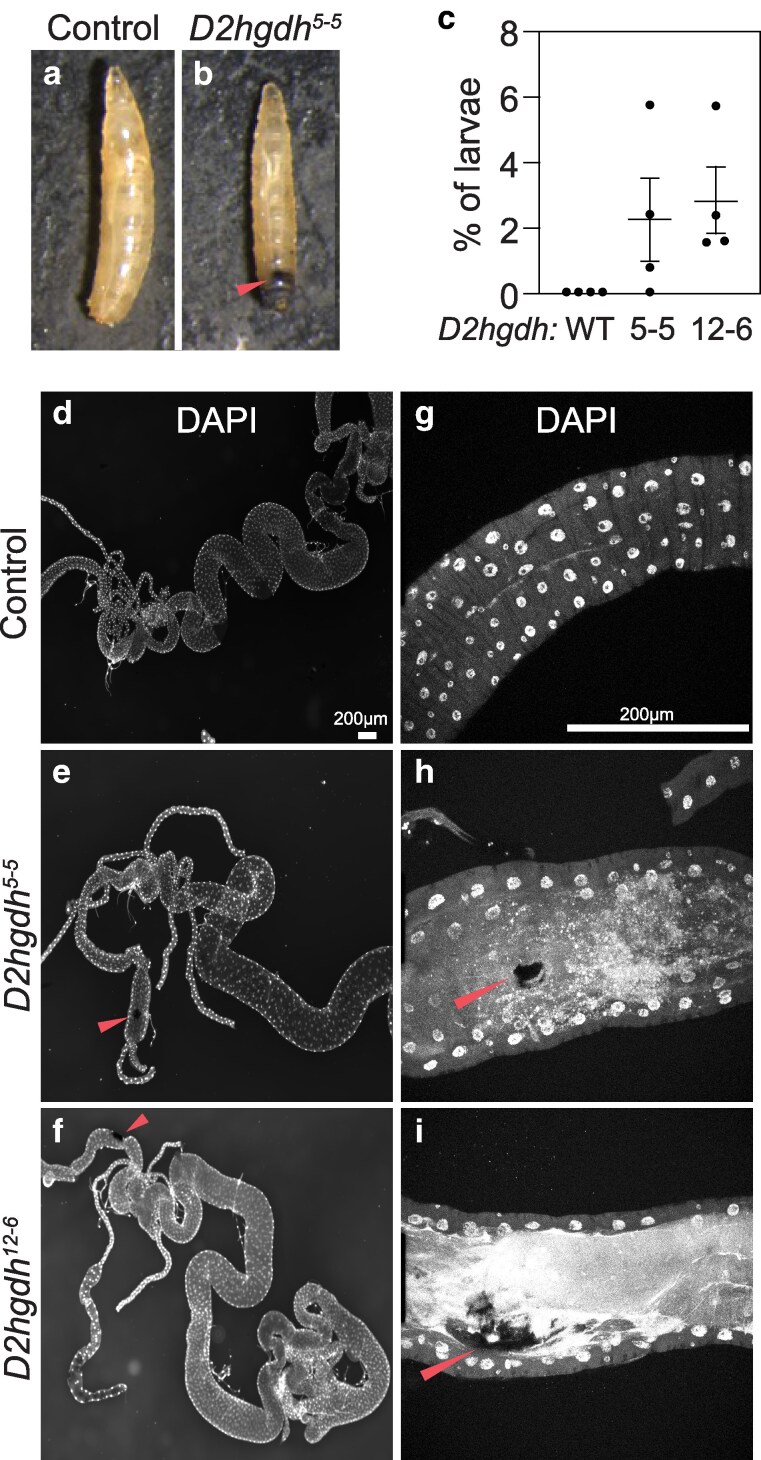 Fig. 2
Fig. 2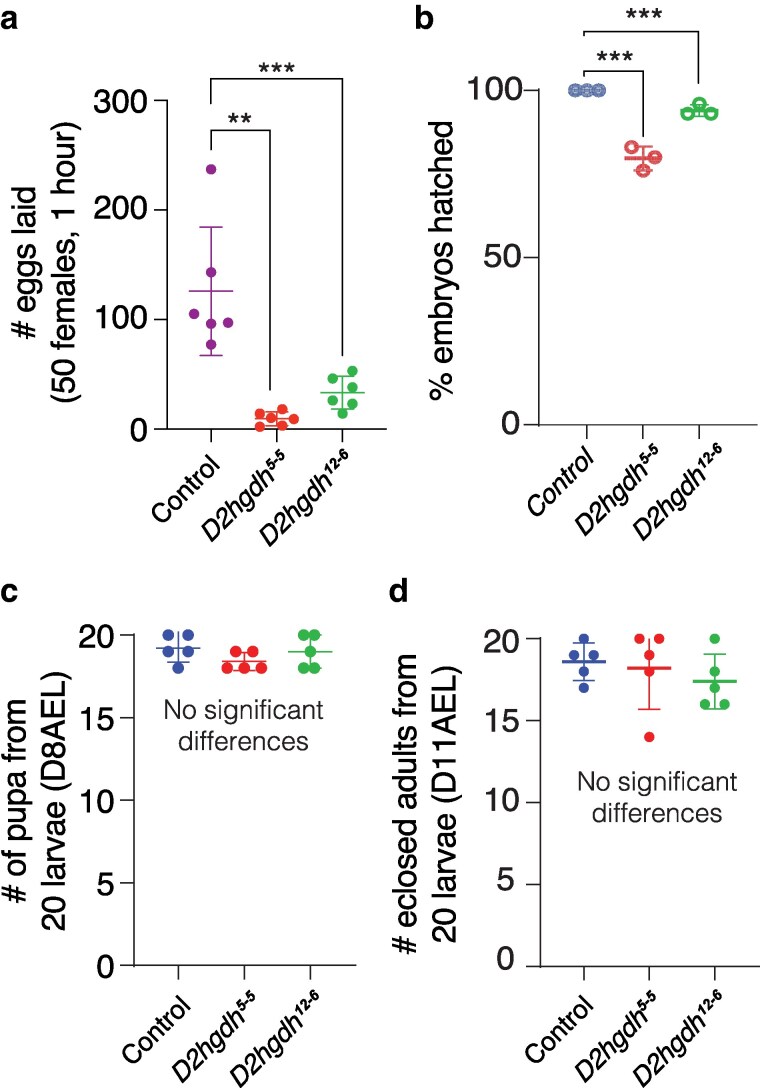 Fig. 3
Fig. 3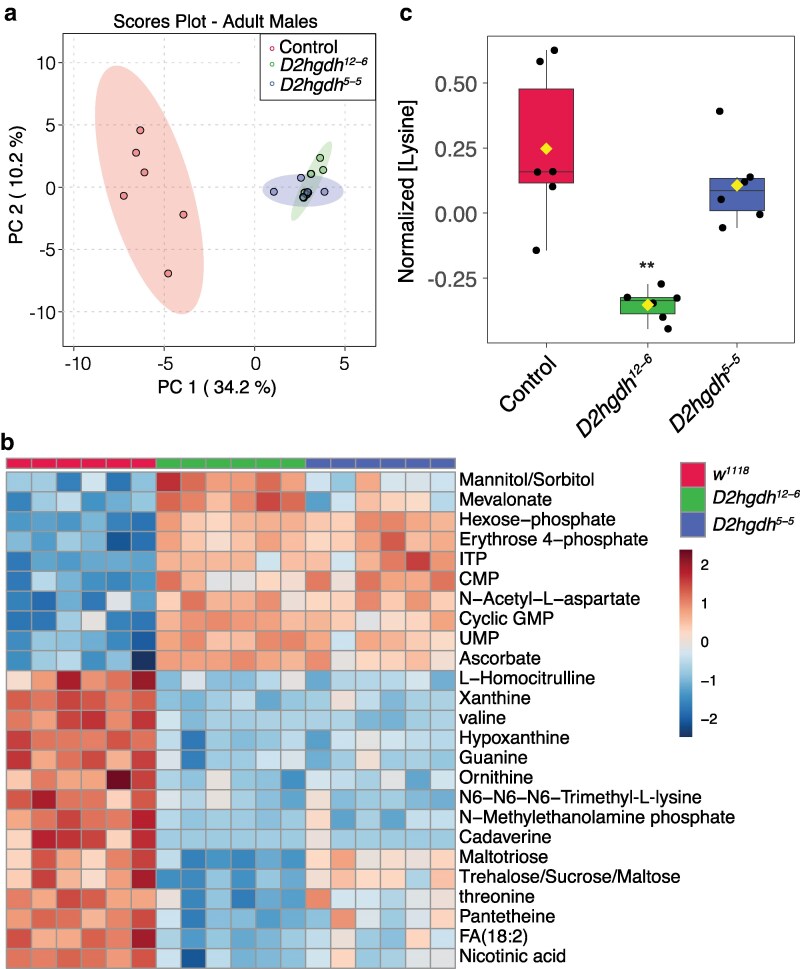 Fig. 4
Fig. 4- —Indiana University Cox Scholars Program
- —American Heart Association10.13039/100000968
- —National Institute of General Medical Sciences10.13039/100000057
- —National Institutes of Health10.13039/100000002
- —MIRA10.13039/100020434
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAmino Acid Enzymes and Metabolism · Epigenetics and DNA Methylation · Cancer, Hypoxia, and Metabolism
Introduction
D-2-hydroxyglutarate (D-2HG) is a hydroxy acid produced from the citric acid cycle intermediate alpha-ketoglutarate (αKG) (Ye et al. 2018; Du and Hu 2021). Under physiological conditions, animals can synthesize D-2HG by 2 enzymatic mechanisms: (1) the enzyme hydroxyacid-oxoacid transhydrogenase (HOT) interconverts αKG and γ-hydroxybutyrate into succinic semialdehyde and D-2HG (Struys et al. 2005b) and (2) the noncanonical activity of the enzyme d-3-phosphoglycerate dehydrogenase reduces αKG to form D-2HG (Fan et al. 2015). However, despite being a side-product of normal cellular metabolism, intracellular D-2HG concentration remain relatively low due, in part, to the activity of the mitochondrial enzyme D-2-hydroxyglutarate dehydrogenase (D2HGDH), which converts D-2HG into αKG in an FAD dependent manner (Achouri et al. 2004).
The D2hgdh-imposed limitation on D-2HG pool size is essential for normal cellular function, as D-2HG is a potent signaling molecule capable of inhibiting a wide-range of αKG-dependent dioxygenases, including the Jmj class of histone lysine demethylases, the Tet family of enzymes, and prolyl hydroxylases (Figueroa et al. 2010; Chowdhury et al. 2011). As a result, increased D-2HG accumulation can induce dramatic changes in chromatin architecture, gene expression, and cellular metabolism, and ultimately lead to a variety of disease phenotypes (Ye et al. 2018; Du and Hu 2021). For example, loss-of-function mutations in the human D2hgdh gene result in an inborn error of metabolism known as D-2-hydroxyglutaric aciduria, which is characterized by elevated urinary D-2HG levels, developmental delays, seizures, and brain abnormalities (Struys et al. 2005a; Struys 2006). Similarly, conditional deletion of D2hgdh in the mouse brain results in increased D-2HG accumulation and leads to impaired activation of radial-glia-like neural stem cells, reduced histone acetylation, and impairment of hippocampus-dependent learning (Liu et al. 2023). Together, these findings highlight the essential role of D2hgdh in limiting pathological D-2HG accumulation.
Because D-2HG interferes with αKG-dependent processes, this molecule is also capable of acting as an oncometabolite and is well-documented to promote the growth and development of a wide range of tumors (Ye et al. 2018; Wang et al. 2020; Chou et al. 2021; Du and Hu 2021). The oncogenic activity of this metabolite, however, is sensitive to the genetic mechanism responsible for inducing aberrant D-2HG accumulation. In nearly all documented studies of D-2HG-positive human tumors, the underlying causes are neomorphic gain-of-function mutations that affect either arginine 132 of IDH1 or arginine 172 of IDH2 (Dang et al. 2009; Losman and Kaelin 2013). These mutations alter the catalytic activity of IDH1/IDH2 in a manner that induces inappropriate D-2HG production (Losman and Kaelin 2013). Such mutations are found in a wide variety of human tumors and are particularly common in lower-grade gliomas, with 70–80% of patients with grade II and III astrocytomas and oligodendrogliomas harboring IDH1 mutations (Yan et al. 2009). Similar findings have emerged from mice, where expression of IDH1 (R132H) in astrocytes induces D-2HG production and promotes glioma development in sensitized backgrounds (Philip et al. 2018). In fact, the widespread occurrence of IDH1/IDH2 mutations in gliomas and acute myeloid leukemia has led to the successful development of FDA approved IDH inhibitors (Gatto et al. 2021; Tian et al. 2022), which are now used in combination therapies to treat IDH1/IDH2-positive tumors.
While there is overwhelming evidence that D-2HG acts as an oncometabolite in cells that express mutant IDH1/IDH2, there is limited evidence indicating that changes in D2hgdh expression or activity drive glioma progression, with only a handful of studies hinting this possibility (Lin et al. 2015; Eckel-Passow et al. 2020). The paucity of evidence linking D2hgdh mutations with glioma occurrence is striking considering the widespread prevalence of IDH1/IDH2 mutations in these tumors (Yan et al. 2009). Such observations raise the question as to why D-2HG only acts as an oncometabolite in specific genetic backgrounds and highlights the need to further study endogenous D-2HG metabolism in both healthy tissues and disease models. Towards this goal, we are using the fruit fly Drosophila melanogaster as a model to understand how loss of D2hgdh activity affects metabolism, growth, and development.
Similar to humans and mice, flies ubiquitously expressing an oncogenic Idh mutant allele (UAS-Idh-R195H) accumulate elevated D-2HG levels. Moreover, UAS-Idh-R195H expression in blood cells using hml-Gal4 induces melanotic mass formation and excess hemocyte production (Reitman et al. 2015). This finding demonstrates that expression of a neomorphic Idh variant in flies partially phenocopies the disease associated Idh1/2 phenotypes observed in other organisms. Furthermore, overexpression of D2hgdh from a UAS-D2hgdh transgene reduces D-2HG accumulation in flies expressing UAS-Idh-R195H, indicating that Drosophila D2hgdh also functions to degrade D-2HG (Reitman et al. 2015). However, these earlier Drosophila studies primarily focused on the oncogenic effects of mutant Idh expression and largely overlooked the endogenous biological roles of D-2HG and D2hgdh. We would also note that while several studies focusing on L-2HG have also quantified D-2HG levels (Li et al. 2017, 2018; Mahmoudzadeh et al. 2020), these investigations did not examine the role of D-2HG in development, physiology, or metabolism.
To facilitate additional studies of D-2HG metabolism in Drosophila, we used a CRISPR/Cas9 approach to generate loss-of-function mutations in D2hgdh. Here, we demonstrate that the resulting mutant lines are viable and display elevated D-2HG levels. Moreover, D2hgdh mutants exhibit decreased fecundity and viability, as well as occasional formation of melanotic masses. Finally, metabolomic analysis of adult male D2hgdh mutants reveal significant changes in essential amino acids levels. Overall, our findings establish Drosophila as a powerful genetic model for studying endogenous D-2HG metabolism and suggest that future studies of the D2hgdh mutants will advance our understanding of how loss of D2hgdh activity results in disease phenotypes.
Materials and methods
Drosophila melanogaster husbandry
Fly stocks were maintained at 25°C on Bloomington Drosophila Stock Center (BDSC) food. Larvae were raised on molasses agar plates with yeast paste spread over the surface as previously described (Li and Tennessen 2017). Since all mutations were generated in a w^1118^ background, we used w^1118^ as a control for all experiments. For all adult analysis, males and virgin females were collected within 8 hours of eclosion, the sexes separated into vials containing BDSC food, and animals were aged for 5–7 days. Flies were then anesthetized using carbon dioxide and collected immediately for subsequent analysis. Flybase was used as an essential reference tool throughout this study (Gramates et al. 2022; Öztürk-Çolak et al. 2024).
Generation of D2hgdh mutations
D2hgdh mutations were generated using CRISPR/Cas9 (Gratz et al. 2013; Sebo et al. 2014). An oligo encoding a guide RNA sequence that targeted exon 3 of the D2hgdh locus (5′ CTTCGCTTAAGCCCGGAAGCACGG 3′) was inserted into the BbsI site of pU6-BbSI-gRNA (Addgene). The gRNA construct was injected into RRID:BDSC_51324 (w[1118]; PBac{y[ + mDint2] = vas-Cas9}VK00027). Two mutations isolated from independently injected embryos were identified by PCR amplification of the region surrounding exon 2 of the D2hgdh locus using oligos 5′-TCCTCCACGATGAGATTCCAAC-3′ and 5′-TGATCGAAGTTCTCCAGGATGC-3′. The resulting PCR product was sequenced using oligos 5′-CCTATCACCACCACCACCATC-3′ and 5′-AAGGACAATCTCGTCGCAGATC-3′. This approach identified 2 D2hgdh deletions: D2hghd^12-6^ consists of a 76 bp deletion (5′-CCACTCTGACCGACAAGGATGTGGCGCATTTCGAGCAGCTCCTGGGCAAGAACTTCGTGCTCACTGAGGACCTGGA-3′). D2hghd^5–5^ contains a 5 bp deletion of 5′-GACCT-3′ within the sequence 5′-GAACTTCGTGCTCACTGAGGACCTGG-3′ (see Fig. 1a).
*D2hgdh mutations induce elevated D-2HG accumulation. a) A schematic diagram illustrating the D2hgdh locus and the mutations D2hgdh12-6 and D2hgdh5-5. The arrow under Δ5–5 points to the location of the D2hgdh5-5 7 bp deletion. The bracket next to Δ12–6 illustrates to the location of the D2hgdh12-6 76 bp deletion. b–g) The amount of D-2HG and L-2HG were simultaneously quantified in both control and D2hgdh mutants during the b, e) L2 larval stage as well as in c, f) adult males and d, g) adult females. Data presented as a scatter plot with the lines representing the mean and standard deviation. P-values calculated using a Wilcoxon/Kruskal–Wallis tests followed by a Dunn's test to compare mutant sample with control. *P < 0.05. **P < 0.01. **P < 0.001. A minimum of 6 biological replicates were analyzed for each genotype. Each replicate contained either 25 mid-L2 larvae or 20 adults (5–7 days post-eclosion). Experiments were repeated 3 times.
Gas chromatography-mass spectrometry analysis
D- and L-2HG were quantified using a protocol specifically designed for analyzing these 2 metabolites (Li and Tennessen 2019). Briefly, either 25 mid-L2 larvae or 20 adults (5–7 days post-eclosion) were placed in a pre-tared 2 mL screw cap tube containing 1.4 mm ceramic beads (Catalog No. 15-340-153; Fisher Scientific), the mass was determined with an analytical balance, and the tube was immediately dropped into liquid nitrogen and subsequently stored at −80°C until processing. Eight hundred microliters of ice cold extraction buffer (90% MeOH with 8 mg of 2,2,3-d3-R,S-2-hydroxyglutarate) was then added to each sample (kept in −20°C enzyme caddies) and tubes were then immediately placed in a Omni Bead Ruptor 24 and homogenized for 30 seconds at 6.4 m/s. Samples were subsequently incubated at −20°C for 1 hour. After the incubation, samples were centrifuged at 20,000 × g for 5 min at 4°C. Six hundred microliters of the supernatant was then transferred into a 1.5 mL microfuge tube and dried overnight using a vacuum centrifuge.
Dried samples were resuspended in 50 μL of R-2-Butanol and 5 μL of HCL and incubated at 90°C for 3 hours with shaking at 300 rpm using an Eppendorf ThermoMixer F1.5. After cooling, 200 μL of water and 500 μL of hexane were added to each tube. The organic phase (hexane) was then transferred to a new tube and dried for 30 minutes using a vacuum centrifuge. The dried samples were resuspended in 60 μL of acetic anhydride and 60 μL of pyridine and incubated at 80°C for 1 hour with shaking at 300 rpm. Samples were then dried for 3 hours using a vacuum centrifuge and resuspended in 60 μL of hexane. Gas chromatography-mass spectrometry (GC-MS) analyses were performed using an Agilent GC6890-5973i MS equipped with a Gerstel MPS autosampler and a 30 m Phenomex ZB5-5 MSi column.
Ultra high-pressure liquid chromatography—mass spectrometry-based metabolomics
D2hgdh mutants and the control strain were raised on BDSC food at 25°C and groups of 20 newly enclosed adult males were transferred to a fresh vial and aged for 5 days, at which point they were placed in a 1.5 mL microfuge tube and drop frozen in liquid nitrogen. Samples were then individually transferred to a pretared 1.4 mm bead tube, the mass was measured using a Mettler Toledo XS105 balance, and the sample was returned to a liquid nitrogen bath prior to being stored at −80°C. Samples were subsequently homogenized in 0.8 mL of prechilled (−20°C) 90% methanol containing 2 μg/mL succinic-d4 acid, for 30 seconds at 6.45 m/s using a bead mill homogenizer located in a 4°C temperature control room. The homogenized samples were incubated at −20°C for 1 hour and then centrifuged at 20,000 × g for 5 min at 4°C. The resulting supernatant was sent for metabolomics analysis at the University of Colorado Anschutz Medical Campus.
Ultra high-pressure liquid chromatography—mass spectrometry metabolomics analyses were performed as previously described (Nemkov et al. 2019). Briefly, the analytical platform employs a Vanquish UHPLC system (Thermo Fisher Scientific, San Jose, CA, USA) coupled online to a Q Exactive mass spectrometer (Thermo Fisher Scientific, San Jose, CA, USA). The (semi)polar extracts were resolved over a Kinetex C18 column, 2.1 × 150 mm, 1.7 µm particle size (Phenomenex, Torrance, CA, USA) equipped with a guard column (SecurityGuard^TM^ Ultracartridge—UHPLC C18 for 2.1 mm ID Columns—AJO-8782—Phenomenex, Torrance, CA, USA) using an aqueous phase (A) of water and 0.1% formic acid and a mobile phase (B) of acetonitrile and 0.1% formic acid for positive ion polarity mode, and an aqueous phase (A) of water:acetonitrile (95:5) with 1 mM ammonium acetate and a mobile phase (B) of acetonitrile:water (95:5) with 1 mM ammonium acetate for negative ion polarity mode. The Q Exactive mass spectrometer (Thermo Fisher Scientific, San Jose, CA, USA) was operated independently in positive or negative ion mode, scanning in Full MS mode (2 μscans) from 60 to 900 m/z at 70,000 resolution, with 4 kV spray voltage, 45 sheath gas, 15 auxiliary gas. Calibration was performed prior to analysis using the Pierce^TM^ Positive and Negative Ion Calibration Solutions (Thermo Fisher Scientific).
Statistical analysis of metabolomics data
Both metabolomics datasets were analyzed using Metaboanalyst 5.0 (Pang et al. 2021). Samples were normalized to mass and the data was preprocessed using log normalization and Pareto scaling. Preprocessed data can be found in Supplementary Table 1. The Quantitative Enrichment analysis tool was used to conduct the enrichment analysis in Supplementary Figs. 1 and 2.
Characterization of developmental phenotypes
Twenty-five males and 50 virgin females for the control and mutant genotypes were placed in an egg laying chamber and placed in a 25°C incubator. For fecundity assays, flies were mated for 24 hours prior to start of the assay, after which egg-laying plates were placed in the chamber for 1-hour intervals and the number of eggs were counted by visual observation in using a standard dissection microscope. Embryonic viability was measured by counting the number of eggs on each egg-laying plate that hatched 30 hours after egg-laying. Pupal viability was measured by transferring 20 newly hatched L1 larvae to a food vial and subsequently counting the number of pupae until 8 days after egg-laying.
Statistical analysis
All data analyses were conducted using Graphpad Prism 9. P-values were calculated using a Wilcoxon/Kruskal–Wallis tests followed by a Dunn's test to compare mutant sample with control.
Results
D2hgdh mutants accumulate excess D-2HG
As a first step towards determining how D2hgdh activity regulates D-2HG levels during the Drosophila life-cycle, we used CRISPR/Cas9 to generate 2 mutations in the D2hgdh gene. As described in the methods, our approach isolated 2 D2hgdh mutant alleles consisting of a 5 bp deletion (D2hgdh^5–5^) and a 76 bp deletion (D2hghd^12–6^), both of which disrupt the conserved catalytic region of the enzyme (Fig. 1a; see Methods for a description of the deleted regions). The resulting mutants are homozygous/hemizygous viable, thus allowing us to determine if D2hgdh mutations alter steady-state D-2HG levels at multiple points during the life-cycle. Consistent with the conserved role of D2hgdh in catabolizing D-2HG, both D2hgdh^5-5^ and D2hghd^12-6^ mutant males, females, and L2 larvae displayed significantly higher D-2HG levels as compared with controls (Fig. 1b–d). In contrast, L-2HG levels were either unchanged or decreased in D2hgdh mutants, indicating that D2hgdh activity is specific to D-2HG (Fig. 1e–g). These results demonstrate that D2hgdh encodes the enzyme responsible for limiting D-2HG accumulation in the fly.
D2hgdh mutants develop melanotic masses and display developmental defects
D2hgdh mutations in humans lead to developmental delays and as well as hypermelanization of the abdominal region—a condition termed angiokeratoma, which is characterized by benign skin lesions (Preston et al. 2019). Consistent with these human disease phenotypes, we observe similar phenomenon in D2hgdh mutants—both D2hgdh^5-5^ and D2hghd^12-6^ mutant larvae exhibit melanotic masses within their abdominal cavity (Fig. 2a–c). While these lesions are relatively rare (present in ∼2% of larvae; Fig. 2c), the masses are easily observed and occur at similar rates in both strains. Moreover, a closer examination of the lesions using confocal microscopy reveal that these masses were often associated with regions of the hindgut that lacked nuclei (Fig. 2d–i). While the significance of these lesions remains unclear, we note that a similar observation was reported in larvae expressing the UAS-Idh-R195H transgene in hemocytes using hml-Gal4 (Reitman et al. 2015), suggesting that these masses stem from elevated D-2HG levels during blood cell development.
D2hgdh mutant exhibit melanotic mass formation. Unlike a) w1118 controls, b) D2hgdh mutant develop melanotic masses in their abdominal region. c) The melanotic mass phenotype occurs in ∼2% of larvae (5 biological replicates of each genotype with n = 30 larvae each). Data presented as a scatter plot with the lines representing the mean and standard deviation. d–i) Larval intestine from w1118 controls and D2hgdh mutants with melanotic masses were dissected and stained with DAPI. e, f) These lesions are most prevalent in the mutant hindgut and h, i) are associated with regions that lack of DAPI stained nuclei. h, i) Red arrows point to melanotic masses in confocal images. The scale bar representing 200 μM in d) applies to e, f). The scale bar representing 200 μM in g) applies to h, i). All the experiments were repeated 3 times.
In addition to the melanotic mass phenotype, D2hgdh^5-5^ and D2hghd^12-6^ mutant females exhibit reduced fecundity, and embryos laid by mutants displayed reduced viability (Fig. 3a, b). However, neither D2hgdh mutation affected larval or pupal viability (Fig. 3c, d). These phenotypes differ from those reported in previous studies using UAS-Idh-R195H, where expression using Tubulin-Gal4 induced an embryonic lethality, and expression driven by Ubiquitin-Gal4 and Actin5C-Gal4 resulted in pupal lethality (Reitman et al. 2015). Thus, developmental phenotypes displayed D2hgdh mutants are milder than those induced by UAS-Idh-R195H overexpression.
*D2hgdh mutants exhibit several developmental phenotypes. When compared with the w1118 control strain, both D2hgdh12–6 and D2hgdh5–5 mutants exhibit significant decreases in a) female fecundity and b) embryonic viability (assessed at 30 hours after egg-laying). In contrast, both c) pupation rate at 8 days after egg-laying (D8AEL) and d) eclosion rate were similar across all 3 strains. Data are presented in scatter plots with mean ± standard deviation. P-values calculated using a Wilcoxon/Kruskal–Wallis tests followed by a Dunn's test to compare mutant sample with control. *P < 0.05. **P < 0.01. **P < 0.001. Note that all P-values compare the indicated mutant samples with the control. For developmental analysis, n = 30 eggs and n = 20 larvae were used. All the experiments were repeated 3 times.
Metabolomics analysis of D2hgdh mutants
To better understand how loss of D2hgdh activity alters Drosophila metabolism and physiology, we used a metabolomics approach to compare adult male D2hgdh^5-5^ and D2hghd^12-6^ mutants with w^1118^ controls. Principle component analysis of the resulting datasets revealed that the metabolomic profiles of both mutant strains significantly differ from the control strain but not from each other, demonstrating that loss of D2hgdh activity appreciably alters the metabolome of adult male flies (Fig. 4a). Moreover, both mutant strains exhibit similar changes in metabolite abundance (Fig. 4b), suggesting that the observed metabolic phenotypes are not the result of genetic background effects.
*Targeted metabolomic analysis of adult D2hgdh mutant males. a) Adult D2hgdh mutant males and w1118 control males were analyzed using a targeted metabolomic approach. a) Analysis of the metabolomics data using principal component (PC) analysis. Note that the mutant data group together and are entirely separated from the controls. b) A heatmap illustrating the top 25 most significantly altered compounds in D2hgdh mutants as compared with controls. c) The relative abundance of lysine in adult D2hgdh mutant males and a w1118 control strain are presented using a box and whiskers plot. Note that lysine levels are either downregulated or unchanged in mutant strains as compared with the control. Figure panels were generated using Metaboanalyst 5.0, as described in the methods. P-values calculated using an ANOVA test followed by a Fisher's test to compare mutant samples with control. *P < 0.01. A total of 6 biological replicates were used for each genotype. Each replicate had 20 adults.
KEGG enrichment analysis of both datasets highlighted changes in several metabolic processes, including the pentose phosphate pathway, glutathione, nucleotide, and amino acid metabolism (Supplementary Figs. 1 and 2). Consistent with these analyses, both mutant strains exhibit elevated levels of erythrose-4-phosphate, an intermediate in the pentose phosphate pathway, as well as the nucleotides ITP, CTP, UMP, and guanine (Fig. 4b). Moreover, we observed decreased levels of the purine degradation products xanthine and hypoxanthine, perhaps suggesting that D-2HG influences purine metabolism. We also observed a highly significant decrease in cadaverine—a breakdown product of lysine commonly found in bacteria. Similarly, N6-N6-N6-trimethyl-L-lysine levels were also significantly lower in mutant males when compared with adults.
Finally, 2 metabolites, 2HG and lysine, failed to show the expected changes in D2hgdh mutants. First, 2HG levels were not significantly altered in our analysis (Supplementary Table 1). We would note, however, that the semi-targeted analysis used for this study does not differentiate between D-2HG and L-2HG and thus the observed 2HG values represents the relative abundance of both molecules. As noted above, D2hgdh mutant male flies tend to harbor normal levels of L-2HG (see Fig. 1), which would partially mask any increase in D-2HG. Second, we also failed to observe an increase in lysine levels, which differs from a recent metabolomic studies of C. elegans, where dhgd-1 mutants displayed increased lysine accumulation (Ponomarova et al. 2023). In fact, we observed the opposite phenomenon, as lysine was slightly downregulated in one mutant strain as compared with controls (Fig. 4c). This result might indicate that Drosophila D2hgdh activity influences lysine metabolism in manner that is distinct from that of the C. elegans D2hgdh homolog.
Discussion
Here, we establish a new genetic model for studying the role of D2hgdh in animal metabolism, physiology, and development. Our studies demonstrate that D2hgdh is essential for regulating D-2HG levels in the fly, as loss of this enzyme in larvae, adult males, and adult females results in a significant increase in D-2HG levels when compared with a control strain. Moreover, we find that D2hgdh mutants display reduced fecundity and embryonic lethality. Finally, metabolomics analysis of these mutants reveals significant changes in the abundance of carbohydrates, nucleotides, and lysine catabolic products. Overall, our studies demonstrate that these D2hgdh novel mutations produce a subset of phenotypes consistent with those observed in human D-2-HG aciduria patients and are appropriate for studying D-2HG within the context of human disease models.
While previous studies in Drosophila have explored the effects of ectopic D-2HG production using UAS-Idh-R195H transgenes (Reitman et al. 2015), the D2hgdh loss-of-function mutations described herein provide an opportunity to examine why neomorphic Idh mutations and D2hgdh deficiency lead to divergent disease phenotypes despite both inducing aberrant D-2HG accumulation. Interestingly, we find that our Drosophila D2hgdh mutants display some phenotypes in common with flies expressing the UAS-Idh-R195H transgene. In this regard, both the occasional presence of melanotic masses within D2hgdh mutant larvae and the metabolomics profile of adult D2hgdh mutant males are notable. The presence of melanotic masses within D2hgdh larvae raises several interesting questions about the nature of this phenotype and similarities with human patients. As mentioned above, the human inborn error of metabolism is associated with hypermelanization of the abdominal region (Preston et al. 2019). While the cause of this phenomenon remains unknown, our observations in the D2hgdh mutant, combined with a previous study describing melanotic masses in flies expressing the UAS-Idh-R195H transgene in hemocytes, suggest that a conserved metabolic mechanism links elevated D-2HG metabolism with this poorly understood phenomenon. One interesting, albeit speculative, possibility is that the hypermelanization phenotype displayed by D2hgdh mutants is that it results from altered immune cell activity—a hypothesis supported by previous studies. First, hml-Gal4 driven expression of a UAS-Idh-R195H transgene in Drosophila hemocytes resulted in higher numbers of these cells, demonstrating that D-2HG promotes immune cell development (Reitman et al. 2015). Secondly, excess D-2HG accumulation alters CD8 T lymphocyte activity and D2hgdh knockdown decreased the anti-tumor activity of these cells (Yang et al. 2022). Future studies should determine if D2hgdh mutant larvae display changes in immune cell numbers and/or an altered immune response.
Finally, the metabolomics profile of D2hgdh mutants display several unexpected changes, with 2 trends being of particular interest. First, we observed significant changes in nucleotide metabolism—several nucleotides were significantly upregulated in both mutant background while xanthine and hypoxanthine, products of purine catabolism, are decreased. We are uncertain as to the significance of these findings, however, the results hint at a role for D-2HG in regulating nucleic acid metabolism. The second important observation is that lysine does not accumulate in D2hgdh mutants. We expected to observe elevated lysine because (1) the enzyme that degrades lysine is inhibited by D-2HG (Chowdhury et al. 2011), (2) C. elegans dhgd-1 mutants accumulate lysine (Ponomarova et al. 2023), and (3) L2hgdh mutants exhibit elevated lysine levels (Mahmoudzadeh et al. 2024). Considering that lysine is either unchanged or decreased in Drosophila mutants, this result suggests that the relationship between D-2HG and lysine metabolism is more complex than previously anticipated. We would also note lysine is unchanged in D2HG aciduria patients (Gibson et al. 1993; Kranendijk et al. 2012), thus highlighting a need to further examine this metabolic interaction.
Supplementary Material
jkaf132_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Achouri Y, Noël G, Vertommen D, Rider MH, Veiga-Da-Cunha M, Van Schaftingen E. 2004. Identification of a dehydrogenase acting on D-2-hydroxyglutarate. Biochem J. 381(1):35–42. doi:10.1042/BJ 20031933.15070399 PMC 1133759 · doi ↗ · pubmed ↗
- 2Chou FJ, Liu Y, Lang F, Yang C. 2021. D-2-hydroxyglutarate in glioma biology. Cells. 10(9):2345. doi:10.3390/cells 10092345.34571995 PMC 8464856 · doi ↗ · pubmed ↗
- 3Chowdhury R, Yeoh KK, Tian Y-M, Hillringhaus L, Bagg EA, Rose NR, Leung IKH, Li XS, Woon ECY, Yang M, et al 2011. The oncometabolite 2-hydroxyglutarate inhibits histone lysine demethylases. EMBO Rep. 12(5):463–469. doi:10.1038/embor.2011.43.21460794 PMC 3090014 · doi ↗ · pubmed ↗
- 4Dang L, White DW, Gross S, Bennett BD, Bittinger MA, Driggers EM, Fantin VR, Jang HG, Jin S, Keenan MC, et al 2009. Cancer-associated IDH 1 mutations produce 2-hydroxyglutarate. Nature. 462(7274):739–744. doi:10.1038/nature 08617.19935646 PMC 2818760 · doi ↗ · pubmed ↗
- 5Du X, Hu H. 2021. The roles of 2-hydroxyglutarate. Front Cell Dev Biol. 9:651317. doi:10.3389/fcell.2021.651317.33842477 PMC 8033037 · doi ↗ · pubmed ↗
- 6Eckel-Passow JE, Drucker KL, Kollmeyer TM, Kosel ML, Decker PA, Molinaro AM, Rice T, Praska CE, Clark L, Caron A, et al 2020. Adult diffuse glioma GWAS by molecular subtype identifies variants in D 2HGDH and FAM 20C. Neuro Oncol. 22(11):1602–1613. doi:10.1093/neuonc/noaa 117.32386320 PMC 7690366 · doi ↗ · pubmed ↗
- 7Fan J, Teng X, Liu L, Mattaini KR, Looper RE, Vander Heiden MG, Rabinowitz JD. 2015. Human phosphoglycerate dehydrogenase produces the oncometabolite D-2-hydroxyglutarate. ACS Chem Biol. 10(2):510–516. doi:10.1021/cb 500683 c.25406093 PMC 4340346 · doi ↗ · pubmed ↗
- 8Figueroa ME, Abdel-Wahab O, Lu C, Ward PS, Patel J, Shih A, Li Y, Bhagwat N, Vasanthakumar A, Fernandez HF, et al 2010. Leukemic IDH 1 and IDH 2 mutations result in a hypermethylation phenotype, disrupt TET 2 function, and impair hematopoietic differentiation. Cancer Cell. 18(6):553–567. doi:10.1016/j.ccr.2010.11.015.21130701 PMC 4105845 · doi ↗ · pubmed ↗