Dataset of the complete mitogenomes of the mushroom corals Fungiidae
Liwei Li, Zhangwang Lu, Zhiwei Liu, Cheng Liang, Jun Wang, Yan Wang

TL;DR
This paper presents a dataset of 24 complete mitochondrial genomes from mushroom corals, revealing insights into their genetic diversity and evolutionary relationships.
Contribution
The study provides a comprehensive mitogenome dataset for Fungiidae corals and identifies minisatellite sequences in intergenic regions.
Findings
Fungiidae mitogenomes share the same 13 protein-coding genes and two rRNA genes as other corals.
Intergenic regions show high diversity, with minisatellites found in specific regions of most mitogenomes.
Phylogenetic trees using mitogenome data offer higher resolution for genus- and species-level relationships.
Abstract
Twenty-four mitogenomes of the mushroom corals (Fungiidae), representing 18 species from 12 genera, were sequenced and annotated. These mitogenomes exhibit high similarity, each containing the same 13 protein-coding genes (PCGs) and two rRNA genes as other scleractinian corals. Compared to the genes, the intergenic regions (IGRs) are more diverse. Interestingly, minisatellite sequences were identified in the IGRs between COX1 and trnM (IGRCOX1-trnM) and between ND4 and rrnS (IGRND4-rrnS) of most Fungiidae mitogenomes. Primarily due to the existence of minisatellites in IGRCOX1-trnM, the length of Fungiidae mitogenomes varies from 16,292 to 17,399 bp. Similar to the phylogenetic tree based on partial COI sequences, Bayesian phylogenetic trees constructed using 13 PCGs, IGRCOX1-trnM and IGRND4-rrnS divide Fungiidae into four distinct clades. However, the latter three trees provide a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
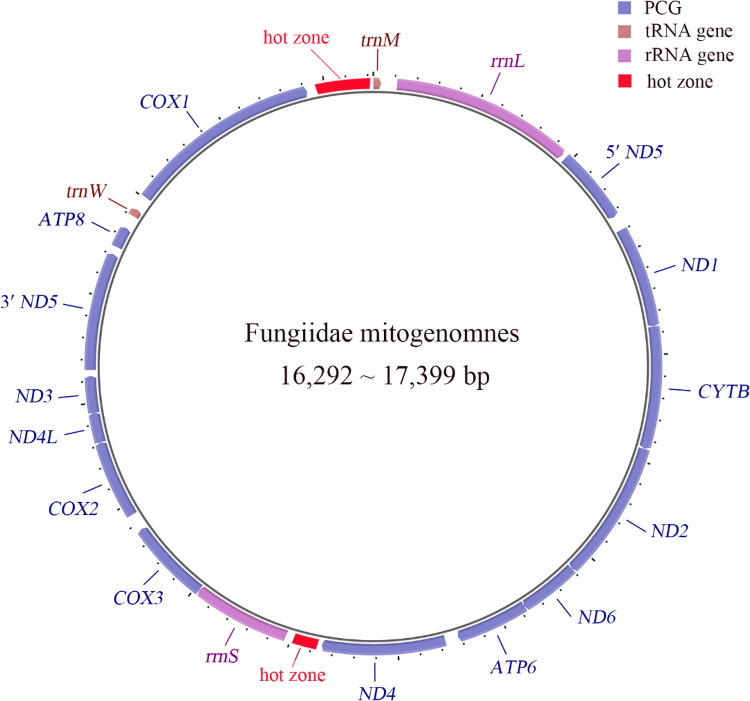 Figure 1
Figure 1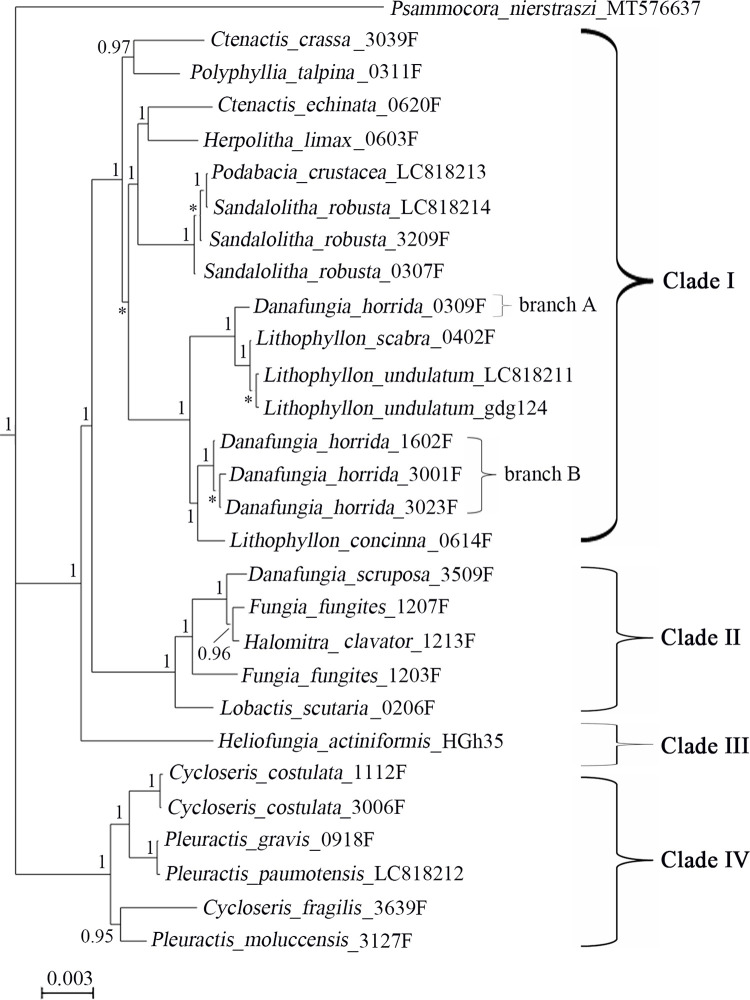 Figure 2
Figure 2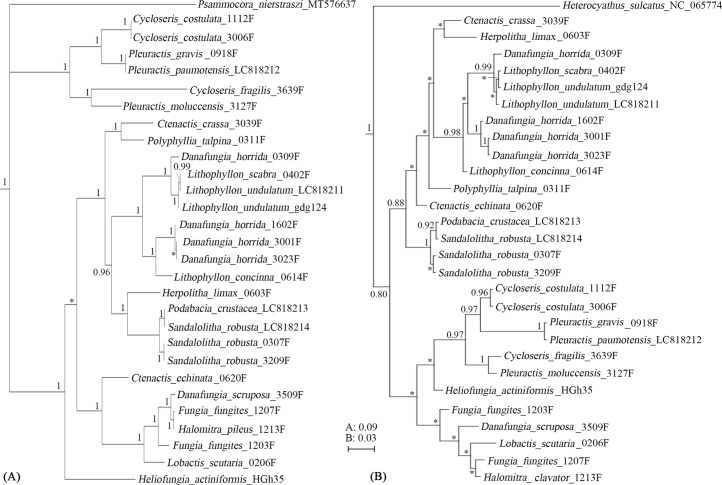 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Fungal Biology and Applications · Plant Pathogens and Fungal Diseases
Specifications TableSubjectBiological SciencesSpecific subject areaBioinformatics, Phylogeny and Evolution, Marine BiologyData formatRaw, Assembled, AnalyzedType of dataFASTA: Mitogenome sequence dataData collectionThe Fungiidae samples were collected at Hainan fringe reefs, Xisha Islands and Zhongsha Atoll (all located in the South China Sea), the longitude and the latitude of sample location ranged from 110°53′48″ to 114°47′14″E and from 15°25′38″N to 19°33′9"N, respectively. Genomic DNA was extracted using the Tianamp Marine Animals DNA Kit (TIANGEN, China) and the sequencing was performed on an Illumina NovaSeq 6000 platform with 150 bp paired-end reads length. The reads were assembled into contigs using MitoZ v2.3 and gene annotation was conducted by MITOS web server (http://mitos.bioinf.uni-leipzig.de/). MrBayes v.3.2.7 was used for the Bayesian inference analysis and ETE Tree Browser was used to visualize the phylogeny tree. The minisatellite sequences in mitogenome were screened with MISA software.Data source locationThe Fungiidae specimens were deposited at School of Marine Biology and Fisheries, Hainan University, Haikou 570,228, China (contact person: Yan WANG, [email protected]).Data accessibilityRepository name: GenBankData identification number: Accession numbers PQ539056, PQ571311-PQ571325, PQ875194-PQ875201Direct URL to data:https://www.ncbi.nlm.nih.gov/nucleotide/PQ539056https://www.ncbi.nlm.nih.gov/nucleotide/PQ571311https://www.ncbi.nlm.nih.gov/nucleotide/PQ571312https://www.ncbi.nlm.nih.gov/nucleotide/PQ571313https://www.ncbi.nlm.nih.gov/nucleotide/PQ571314https://www.ncbi.nlm.nih.gov/nucleotide/PQ571315https://www.ncbi.nlm.nih.gov/nucleotide/PQ571316https://www.ncbi.nlm.nih.gov/nucleotide/PQ571317https://www.ncbi.nlm.nih.gov/nucleotide/PQ571318https://www.ncbi.nlm.nih.gov/nucleotide/PQ571319https://www.ncbi.nlm.nih.gov/nucleotide/PQ571320https://www.ncbi.nlm.nih.gov/nucleotide/PQ571321https://www.ncbi.nlm.nih.gov/nucleotide/PQ571322https://www.ncbi.nlm.nih.gov/nucleotide/PQ571323https://www.ncbi.nlm.nih.gov/nucleotide/PQ571324https://www.ncbi.nlm.nih.gov/nucleotide/PQ571325https://www.ncbi.nlm.nih.gov/nucleotide/PQ875194https://www.ncbi.nlm.nih.gov/nucleotide/PQ875195https://www.ncbi.nlm.nih.gov/nucleotide/PQ875196https://www.ncbi.nlm.nih.gov/nucleotide/PQ875197https://www.ncbi.nlm.nih.gov/nucleotide/PQ875198https://www.ncbi.nlm.nih.gov/nucleotide/PQ875199https://www.ncbi.nlm.nih.gov/nucleotide/PQ875200https://www.ncbi.nlm.nih.gov/nucleotide/PQ875201Related research articleNA
Value of the Data
1
- •This mitogenome dataset can benefit researchers and students working on Fungiidae systematics, phylogeography, and evolution.
- •The diverse IGRs in the mitogenomes would be useful for the development of Fungiidae DNA barcode.
- •The minisatellites found in most Fungiidae mitogenomes can be applied to the study of population biology.
Background
2
The hard coral family Fungiidae (Dana, 1864) belongs to the phylum Cnidaria, subphylum Anthozoa, class Hexacorallia and order Scleractinia [1]. Species in this family grow in a mushroom-like shape, earning them the common name 'mushroom corals' [2]. Currently, this family comprises 16 genera, exhibiting significant evolutionary divergence and rich species diversity [1]. Fungiidae plays a crucial role in maintaining the balance of coral reef ecosystems [2].
Most fungiid corals consist of large, free-living single polyps. They are distinguished from other scleractinian corals by the presence of compound synapticulae between the septo-costal units [2]. Morphological identification at the genus and species levels primarily relies on differences in life cycles and inconsistencies in skeletal architecture, e.g. the corallum wall, septa, and costae [2,3]. However, coral morphological characteristics respond to environmental changes, and many fungiid coral species exhibit diverse and plastic phenotypic variations [4]. Therefore, a combination of molecular analysis and morphological characteristics is typically used for a more accurate classification of Fungiidae. Commonly used molecular markers include partial sequences of cytochrome oxidase subunit I (COI) [5] and the internal transcribed spacer (ITS) [6,7]. However, the resolution of Fungiidae COI is insufficient. In some cases, COI barcode sequences from different genera, e.g. Lithophyllon undulatum (GenBank no EU149887), Lithophyllon scabra (EU149894), and Danafungia horrida (LC191483), are identical. Regarding ITS, while the high diversity among individuals within a species can compensate for the low species resolution of COI barcodes, it may also obscure or complicate phylogenetic relationships [5]. Additionally, individual coral colonies usually host a high degree of intragenomic ITS variation [8], which may lead to the existence of overlapping peaks in sequencing and result in unreliable sequence alignments [9].
Besides COI, other mitogenome sequences including rRNA genes [10] and intergenic regions (IGR) [11], are also used for the species identification of scleractinian corals. Currently, only four Fungiidae mitogenomes have been published [12]. Here we present a dataset of 24 complete Fungiidae mitogenomes, encompassing 12 of the 16 existing Fungiidae genera, and analyzed the nucleotide diversity of their protein coding genes (PCGs), rRNA genes and IGRs, so as to provide basic information for the development of more reliable DNA barcode in Fungiidae. Interestingly, minisatellite sequences were found in the intergenic regions between COX1 and trnM (IGR*^COX1^^-^^trnM^) and between ND4 and rrnS (IGR^ND4-rrnS^*) in most Fungiidae mitogenomes. These minisatellites could be valuable for the population genetic studies of Fungiidae [13].
Data Description
3
A total of 24 Fungiidae mitogenomes, representing 18 species from 12 genera, were sequenced and annotated (Table 1). These mitogenomes exhibit a high degree of similarity (≥ 96.47 %, Supplementary Table 1). In some cases, the similarity between different genera is greater than interspecific similarity, e.g. the similarity between Danafungia scruposa_3509F and Fungia fungites_1207F is 99.81 %, which is greater than that of Danafungia scruposa_3509F and Danafungia horrida_0309F (98.37 %) (Supplementary Table 1).Table 1. Information of samples, mitogenomes and sequence read archive (SRA).Table 1. Sample nameSample locationLatitude/LongitudeSpeciesMitogenome GenBank No.Mitogenome length (bp)SRA accession No.3039FLingyang Reef16°26′46″N/111°38′35″ECtenactis**crassaPQ87519416,292SRR342228710620FZhizhang Shoal15°59′23″N/114°38′10″ECtenactis**echinataPQ57131116,663SRR341704761112FZhongbei Shoal16°04′17″N/114°24′51″ECycloseris**costulataPQ57131216,451SRR342251463006FLingyang Reef16°26′46″N/111°38′35″ECycloseris**costulataPQ87519516,450SRR341694293639FBeidaoIsland16°35′10″N/112°11′15″ECycloseris**fragilisPQ87519616,522SRR342228700309FHaijiu Shoal15°37′59″N/114°35′21″EDanafungia**horridaPQ57131317,332SRR342251473001FLingyang Reef16°26′46″N/111°38′35″EDanafungia**horridaPQ87519716,771SRR342251491602FMeixiShoal15°25′38″N/114°12′28″EDanafungia**horridaPQ53905616,577SRR342168633023FLingyang Reef16°26′46″N/111°38′35″EDanafungia**horridaPQ87519816,771SRR342168223509FHuaguang Reef16°9′41″N/111°24′18″EDanafungia**scruposaPQ87519917,061SRR342251481207FMeibin Shoal16°02′14″N/114°11′32″EFungia**fungitesPQ57131416,455SRR342011601203FMeibin Shoal16°02′14″N/114°11′32″EFungia**fungitesPQ57131516,342SRR342081421213FMeibin Shoal16°02′14″N/114°11′32″EHalomitra**clavatorPQ57131616,551SRR34216823HGh35Huaguang Reef16°15′26″N/111°38′58″EHeliofungia**actiniformisPQ57131717,065SRR342168620603FZhizhang Shoal15°59′23″N/114°38′10″EHerpolitha**limaxPQ57131816,619SRR342011610614FZhizhang Shoal15°59′23″N/114°38′10″ELithophyllon**concinnaPQ57131916,459SRR342081350402FJimeng Shoal15°42′09″N/114°41′05″ELithophyllon**scabraPQ57132117,014SRR34201159gdg124Qinglan19°33′9″N/110°53′48″ELithophyllon**undulatumPQ57132017,026SRR342228690206FLexiShoal15°51′41″N/114°25′23″ELobactis**scutariaPQ57132216,811SRR342011580918FBiwei Shoal16°11′24″N/114°47′14″EPleuractis**gravisPQ57132316,497SRR341738383127FQuanfu Island16°35′1″N/111°41′17″EPleuractis**moluccensisPQ87520016,854SRR342168640311FHaijiu Shoal15°37′59″N/114°35′21″EPolyphyllia**talpinaPQ57132417,108SRR341738373209FYinyuReef16°32′2″N/111°39′36″ESandalolitha**robustaPQ87520117,309SRR342168650307FHaijiu Shoal15°37′59″N/114°35′21″ESandalolitha**robustaPQ57132517,399SRR34173836
The 24 Fungiidae mitogenomes all contain the same 13 PCGs (protein-coding genes) and two rRNA genes as other scleractinian corals (Fig. 1 and Supplementary Table 2). Comparing to the PCGs and rRNA genes, the intergenic regions (IGRs) are more diverse (Table 2). The nucleotide diversity of IGR*^COX1^^-^^trnM^* and IGR*^ND4^^-^^rrnS^* is highest (referred to as "hot zones") and in these two IGRs of most Fungiidae mitogenomes, minisatellite sequences were found (Table 3 and Supplementary material 1). The minisatellites located in IGR*^COX1^^-^^trnM^* have an average length of 277 bp, which is longer than those in IGR*^ND4^^-^^rrnS^* (average 214 bp). Furthermore, the length of IGR*^COX1^^-^^trnM^* minisatellites is strongly positively correlated with the length of mitogenomes (Pearson correlation coefficient 0.786), whereas the length of IGR*^ND4^^-^^rrnS^* minisatellites shows no significant correlation (Pearson correlation coefficient 0.092).Fig. 1. The maps of Fungiidae mitogenomes. The Fungiidae mitogenomes contain the same 13 PCGs (protein-coding genes) and two rRNA genes as other scleractinian corals. Comparing to the genes, the intergenic regions (IGR) are more diverse, especially in IGR*^COX1^^-^^trnM^* and IGR*^ND4^^-^^rrnS^* (the hot zones) of most Fungiidae mitogenomes, some minisatellite sequences were located. Primarily due to the existence of minisatellites in IGR*^COX1^^-^^trnM^, the length of Fungiidae mitogenomes ranges from 16,292 to 17,399 bp.Fig 1. Table 2The nucleotide diversity (Pi) of mitochondrial genes and IGRs.Table 2. Gene/IGRNucleotide diversity (Pi)ND4L0.0029ND10.0037CYTB0.0040ND40.0044rrnL0.0054ND20.0055ATP60.0060ND50.0069COX30.0076COX20.0076IGR (COX3-COX2)0.0079ND60.0079rrnS0.0080IGR (3′ND5-ND1)0.0117ND30.0130ATP80.0183COX10.0309IGR (trnM-rrnL)0.0506IGR (ND4-rrnS)0.1329IGR (COX1-trnM)0.3425Note:* Nucleotide diversity was calculated for only five IGRs, other IGRs were excluded due to their short length (<100 bp), making them unsuitable for DNA barcode development.Table 3. Information of minisatellites located in the IGRs.Table 3. SpeciesGenBank No.IGR*^COX1^^-^^trnM^IGR^ND4^^-^^rrnS^Minisatellite length (bp)Minisatellite motifMinisatellite positionMinisatellite length (bp)Minisatellite motifMinisatellite positionCtenactiscrassa*PQ875194102(GGCAATAATGTCGATAATAATTTGGATGGAG GGAATATTTGGATGCG)_2_15,785–15,886–(None)^#^–*Ctenactisechinata*PQ571311483(AATGTTAATGATAATTTTGAGATCGAATTTC CAGTTAATGAAGGGGGCCAG)915,576–16,058–(None)–Cycloseris**costulataPQ571312–(None)^#^––(None)–Cycloseris**costulataPQ875195–(None)––(None)–Cycloseris**fragilisPQ875196–(None)––(None)–Danafungia**horridaPQ571313595(AGATACGGAGAAAGAGTGTGAGAAAGATTTA AGGACAAAAACATT)8(GAAAAAGAGTGTGAAATAGATTTAATGAAAA AGAGTGTGAAATAGATTTAATGAAAAA GAGTGTGAAAAATATTTG)316,585–17,179194(TTAAAAAGCCTTTGGTCTAAGTTAGTCTTTTAGTTTTGGGGATTTAAAAACTTTTGGTCTAAGTTAGACAGACGGGCCTGTCCTTTGGTTTTAAGGA)28885–9078Danafungia**horridaPQ875197217(CTAATAATTTAAATGAGGGGGTTCCAATGCC TGCCG)616,092–16,308484(TTTGGTCTAAGTTAGTCTTTTAGTTTTGGGGATTTAAAAACTTTTGGTCTAAGTTAGACAGGCGGGTCTGTCCTTTGGTTTTAAGGATTAAAAAGCT)58896–9379Danafungia**horridaPQ539056217(CTAATAATTTAAATGAGGGGGTTCCAATGCC TGCCG)615,898–16,114290(TTTGGTCTAAGTTAGTCTTTTAGTTTTGGGGATTTAAAAACTTTTGGTCTAAGTTAGACAGGCGGGTCTGTCCTTTGGTTTTAAGGATTAAAAAGCT)38896–9185Danafungia**horridaPQ875198217(CTAATAATTTAAATGAGGGGGTTCCAATGCC TGCCG)616,092–16,308484(TTTGGTCTAAGTTAGTCTTTTAGTTTTGGGGATTTAAAAACTTTTGGTCTAAGTTAGACAGGCGGGTCTGTCCTTTGGTTTTAAGGATTAAAAAGCT)58896–9379Danafungia**scruposaPQ875199260(TGATTAGACAAGGAGCAATAACATTGAGTTC TTTAAAAAAAAGTTGCTACCC)516,790–17,049387(TAAGGATTAAAAAGCTTTTGGTCTAAGCTAGTCTTTTGGTTTCGGAGATTAGAAAACTTTTGGTTTAAGTTAGACAAGCTGATTAGTCTTTTGGTTT)48960–9346Fungia**fungitesPQ57131454(CTGAAACTGTTAGTGCTGCAGCTTCCT)216,204–16,257291(TAGTCTTTTAGTTTTGGGGATTAAAAAGCTTTTGATCTAAGT)68946–9236Fungia**fungitesPQ571315153(TCCAGTAAATCAGGAATTAAATAATGCAAAT CAGGGTGTTGACGGAGAACA)315,692–15,844110(TCTAAGTTAGACAGGCTAGTTTGTCCTTTGGTTTTGGGGATTAAAAAGCTTTTGG)28994–9103Halomitraclavator*PQ571316108(CTGAAACTGTTAGTGCTGCAGCTTCCT)416,246–16,353333(TAGTCTTTTAGTTTTGGGGATTAAAAAGCTTTTGATCTAAGT)78947–9279*Heliofungiaactiniformis*PQ571317335(CTAAAACGGAATCCACTAATGCTGACGTACA AATTAGTGAAGATG)5; (TTTGTATTTACTGGTGGGGGGTCTTTTGTTGA TGTTGTGTGATTATTTTTATATT)_2_16,205–16,339;16,479–16,58874(GTTAGTCTTTTGGTTTTGGGGATTAGAAAGCTGGCTG)28954–9027Herpolitha**limaxPQ571318180(GAAAGTTGTAATCAAGAATCATCGGATGCTG GGGGGGAAATAAAT)416,020–16,199169(CTTTTGGTCTAACTTAGACGACAGGCTGGTCTGTCCTTTGGTTTTGGGGATTAACCA)38623–8791Lithophyllon**concinnaPQ57131984(CCCTGCCGCTAATAATGTTAATGAAGGGGTT CAGGTACCTGC)_2_15,812–15,895110(CTTTTGGTCTAAGTTAGACAGGCGGGTCTGTCCTTTGGTTTTGGGGATTAAAAAG)28936–9045Lithophyllon**scabraPQ571321225(AGAGTGTGAGAAAGATTTAAAGACAAAAACATCAGATACGGAGAA)2(ATTTAATGAAAAAGAGTGTGAAATAG)416,570–16,794174(GATTAAAAAGCTTTTGGTCTAAGTTA)58884–9057Lithophyllon**undulatumPQ571320294(AGAGTGTGAGAAAGATTTAAAGACAAAAACATCAGATACGGAGAA)2(ATTTAATGAAAAAGAGTGTGAAATAG)3(GAAAAAGAGTGTGAAAAATATTTGGAAAAAGACCTGTTTAATGA)_2_16,570–16,863174(GATTAAAAAGCTTTTGGTCTAAGTTA)58884–9057Lithophyllonundulatum*LC818211 ^✝^225(AGAGTGTGAGAAAGATTTAAAGACAAAAACATCAGATACGGAGAA)2(ATTTAATGAAAAAGAGTGTGAAATAG)316,570–16,794174(GATTAAAAAGCTTTTGGTCTAAGTTA)58884–9057*Lobactisscutaria*PQ571322152(GTAAAAAAGCAGGTAGCCATCCTGGAAACAGGTAG)2(AAAAAGCAGGTAGCCATCCTGGAAACAGGTAGGTAGCCATCCTGGAAACAGGTAGGT)2(AGGTAGCCATCCTGGAAACAGGT)416,629–16,780304(TTTGGTCTAAGTTAGACACGCTGATTAGTCTTTTAGTTTTAAGGATTAAAAAGCG)4; (AAAGCTTTTGGTCTAAGTTAGTCTTTTGGTTTCGGAGATTCG)28933–9152; 9244–9327Pleuractis**gravisPQ571323–(None)––(None)–Pleuractis**moluccensisPQ875200–(None)––(None)–Pleuractis**paumotensisLC818212 ^✝^–(None)––(None)–Polyphyllia**talpinaPQ571324505(GATTTACAAACCGAATTATTGAATATGGAGGGGGAAATTAATACCGATTTACAAACCGAATTATTGAATATGGAGGGGGAAGTTAATACC)4(GAGTGGAAATTAATGAGGATTTACAAACCGAATTATTGAATATGGAGGGGGAAACTAATACCGATTTAA)_2_16,373–16,877110(AGCTTTTGGTCTAAGTTAGACAGGCTGATCTGTCCTTTGGTTTTGGGGATTAAAA)29237–9346Sandalolitha**robustaPQ875201448(TTTGCTACTGAGGGAGAGGTTGATGAAAGTTTGGAAAAAGAGTTA)3(GTTTAAAAAAAGAATAATAATGATGACATGATAAATAAGGATTTTTGGTTTTGGG)2(TAAAAAAAGAATAATAATGATGACATGATAAATAAAAAAAGAATAATAATGATGACATAATAAA)_2_16,745–17,192104(CTTTTGGTCTAAGTTAGACAGGTTGGTTAGTCTTTTAGTTTTGGGGATT)28936–9039Sandalolitha**robustaPQ571325601(AGTTTGGAAAAAGAGTTATTTGCTACTGAGGGAGAGGTTGATGAAAGTTTGGAAGAAGAATTGTTTGCTACTGAGGGAGAGGTTGATGAA)2(TTTGCTACTGAGGGAGAGGTTGATGAAAGTTTGGAAAAAGAGTTA)2(GTTTAAAAAAAGAATAATAATGATGACATGATAAATAAGGATTTTTGGTTTTGGG)2(TAAAAAAAGAATAATAATGATGACATGATAAATAAAAAAAGAATAATAATGATGACATAATAAA)_2_16,682–17,282104(CTTTTGGTCTAAGTTAGACAGGTTGGTTAGTCTTTTAGTTTTGGGGATT)28936–9039Sandalolitha**robustaLC818214 ^✝^315(GGTTTGGAAAAAGAAGCATCCGATATTAACGAAGAAATTGACAAC)2; (TTTGCTACTGAGGGAGAGGTTGATGAAAGTTTGGAAAAAGAGTTA)516,109–16,198; 16,757–16,981104(CTTTTGGTCTAAGTTAGACAGGTTGGTTAGTCTTTTAGTTTTGGGGATT)28936–9039Podabacia**crustaceaLC818213 ^✝^315(GGTTTGGAAAAAGAAGCATCCGATATTAACGAAGAAATTGACAAC)2; (TTTGCTACTGAGGGAGAGGTTGATGAAAGTTT GGAAAAAGAGTTA)516,098–16,187; 16,746–16,970104(CTTTTGGTCTAAGTTAGACAGGTTGGTTAGTCTTTTAGTTTTGGGGATT)28936–9039Note: * Imperfect minisatellite, which contains variations within its repeat pattern. ^#^ No minisatellite was found. ^✝^ Sequences retrieved from GenBank.
The same species (e.g. Danafungia horrida and Sandalolitha robusta) may contain the same minisatellite motifs, but the number of repeats of motifs can vary (Table 3).
The Bayesian phylogenetic trees of Fungiidae based on 13 protein-coding genes (PCGs) was constructed. Consistent with the tree inferred from COI [14], the tree based on 13 PCGs divides Fungiidae into four phylogenetic clades (Fig. 2). Most genera are polyphyletic. Notably, only the mitogenomes of clade IV (Cycloseris and Pleuractis) don’t contain the minisatellite sequence (Table 3). On the level of species, the two Fungia fungites individuals locates in different branches: Fungia fungites_1207F is the Clade A described by Oku et al. (2020) and Fungia fungites_1203F is Clade B [15]. Four Danafungia horrida individuals also locates in two different branches (Fig. 2). The three individuals of branch B share the same minisatellite motif, which differs from the motif found in branch A (Table 3).Fig. 2. The Bayesian phylogenetic tree of Fungiidae based on 13 protein-coding genes. Numbers at nodes indicate Bayesian posterior probabilities. Stars indicate branch support values under 0.80.Fig 2
The phylogenetic trees based on the two hot zones (IGR*^COX1^^-^^trnM^* and IGR*^ND4^^-^^rrnS^) (Fig. 3) yield results similar to those of the 13-PCGs tree, but with higher resolution. The scale bars for the IGR^COX1^^-^^trnM^* and IGR*^ND4^^-^^rrnS^* trees are 30 times and 10 times longer, respectively, than that of the 13-PCGs tree.Fig. 3. The Bayesian phylogenetic trees based on IGR*^COX1-trnM^* (A) and IGR*^ND4-rrnS^* (B). Numbers at nodes indicate Bayesian posterior probabilities. Stars indicate branch support values under 0.80. The scale bar of IGR*^COX1-trnM^* tree is 30 times longer than that of 13 PCGs tree.Fig 3
Experimental Design, Materials and Methods
4
Samples collection and identification
4.1
The Fungiidae samples were collected at Hainan fringe reefs, Xisha Islands and Zhongsha Atoll (all located in the South China Sea) from 2020 to 2024 (Table 1). The longitude and the latitude of sample location range from 110°53′48″ to 114°47′14″E, and from 15°25′38″N to 19°33′9"N, respectively. Coral sampling was conducted under permits approved by the Department of Ocean and Fisheries of Hainan Province (China). Morphological species identification was based on colony shape, polyp structure, and septa [1,2], while molecular identification focused on a portion of the cytochrome oxidase I (COI, 500 bp) gene using fungiid-specific primers FungCOIfor1 5′-CTGCTCTTAGTATGCTTGTA-3′ and FungCOIrev2 5′-TTGCACCCGCTAATACAG-3′ [5], as well as an approximately 800 bp region of rDNA using the primers ITS4 5′-TCCTCCGCTTATTGATATGC-3′ [6] and A18S 5′-GATCGAACGGTTTAGTGAGG-3′ [7].
A total of 18 species from 12 genera were identified (Table 1). For samples that were difficult to identify or prone to confusion, e.g. Danafungia horrida, Fungia fungites, Cycloseris costulata and Sandalolitha robusta, we selected two or more samples for mitogenome sequencing. In total, 24 samples were selected (Table 1).
DNA sequencing
4.2
Genomic DNA was extracted using the Tianamp Marine Animals DNA Kit (TIANGEN, CHINA). After DNA isolation, 1 μg of purified DNA was fragmented to ∼500 bp using the Covaris M220 system. The short-insert libraries were constructed according to the manufacturer’s instructions (TruSeq™ Nano DNA Sample Prep Kit, Illumina) and then the sequencing was performed on an Illumina NovaSeq 6000 platform (BIOZERON Co., Ltd, Shanghai, China) with 150 bp paired-end reads length. Raw Illumina sequencing reads have been deposited in the NCBI Sequence Read Archive (SRA), and SRA accession numbers were assigned (Table 1).
Mitogenome assembly and gene annotation
4.3
Prior to assembly, raw reads were filtered by Trimmomatic 0.39 [16] to remove the reads with adaptors, the reads showing a quality score below 20 (Q < 20), the reads containing a percentage of uncalled bases (“N” characters) ≥ 10 % and the duplicated sequences. The filtered reads were assembled into contigs using MitoZ v2.3, which typically produced multiple contigs. Manual curations were conducted for each dataset. Specifically, mitochondrial contigs with >80 % query coverage were identified through BLAST v2.8.1+ alignment against reference mitogenomes. These contigs were then manually ordered, oriented, and joined based on the structure of the corresponding reference mitogenomes. MUMmer 3.23 was subsequently used to confirm the circularity of the assembled mitogenomes. As a result of this process, each final mitochondrial genome assembly consisted of a single circular contig.
The assembled Fungiidae mitogenome sequences were imported into MITOS web server (http://mitos.bioinf.uni-leipzig.de/) [17] for gene annotation. The start and stop codon positions of each gene were manually corrected. Mitogenome visualization was performed using the Proksee online platform (https://proksee.ca/) [18].
Mitogenome analysis
4.4
The 24 Fungiidae mitogenomes sequenced in this study (Table 1), along with four additional Fungiidae mitogenomes retrieved from GenBank [12], were used for the following analysis.
Mitogenome similarity, nucleotide diversity and minisatellite screening
4.4.1
The similarity among Fungiidae mitogenomes was calculated using the BLAST program of NCBI (https://blast.ncbi.nlm.nih.gov/). The nucleotide diversity (Pi) of protein coding genes, rRNA genes and IGRs was calculated using DnaSP version 5.0 [19]. The minisatellite sequences were screened with MISA software [20] and the imperfect minisatellites, which contains variations within its repeat pattern, were checked manually. The correlation between the length of minisatellites and the length of mitogenomes was calculated using the PEARSON function (for Pearson’s correlation coefficient) in Microsoft Excel.
Phylogenetic analysis
4.4.2
Bayesian inference phylogeny trees were constructed based on 13 PCGs and the two hot zones (IGR*^COX1^^-^^trnM^* and IGR*^ND4^^-^^rrnS^). For the 13-PCGs tree and IGR^COX1^^-^^trnM^* tree, the Psammocora nierstraszi mitogenome (GenBank no MT576637) was set as outgroup, and for the IGR*^ND4^^-^^rrnS^* tree, Heterocyathus sulcatus mitogenome (NC_065774) was used as outgroup. Sequences were firstly aligned separately using MAFFT v.7.407 [21], and then Gblocks v.0.91b [22] was applied to remove ambiguously aligned regions using default settings. Based on the corrected Akaike Information Criterion (AICc) [23], the best partition scheme, as well as the best-fit nucleotide substitution models for the respective partitions, were detected by PartitionFinder 2 [24]. MrBayes v.3.2.7 [25] was used for the Bayesian inference analysis, and ETE Tree Browser [26] was used to visualize the resulting phylogeny tree.
Limitations
Not applicable.
Ethics Statement
The authors have read and followed the ethical requirements for publication in Data in Brief. The authors confirm the current work does not involve human subjects, animal experiments, or any data collected from social media platforms.
CRediT authorship contribution statement
Liwei Li: Investigation. Zhangwang Lu: Methodology. Zhiwei Liu: Investigation. Cheng Liang: Investigation. Jun Wang: Data curation. Yan Wang: Conceptualization, Writing – review & editing.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hoeksema B.W.Cairns S.World List of Scleractinia Fungiidae Dana 20251846. Accessed through World Register of Marine Species athttps://www.marinespecies.org/aphia.php?p=taxdetails&id=196100 on 2025-1-21
- 2Hoeksema B.Taxonomy, phylogeny and biogeography of mushroom corals (Scleractinia: fungiidae)Zoologische Verhandelingen.254119891295
- 3Grinblat M.Cooke I.Shlesinger T.Ben-Zvi O.Loya Y.Miller D.J.Cowman P.F.Biogeography, reproductive biology and phylogenetic divergence within the Fungiidae (mushroom corals)Mol. Phylogenet. Evol.164202110726510.1016/j.ympev.2021.10726534274488 · doi ↗ · pubmed ↗
- 4Gittenberger A.Hoeksema B.W.Phenotypic plasticity revealed by molecular studies on reef corals of Fungia (Cycloseris) spp.(Scleractinia: fungiidae) Near river outlets Contributions to Zool.7503–042006195201
- 5Gittenberger A.Reijnen B.T.Hoeksema B.W.A molecularly based phylogeny reconstruction of mushroom corals (Scleractinia: fungiidae) with taxonomic consequences and evolutionary implications for life history traits Contribut. Zool. Bijdragen Tot De Dierkunde 8022011107132
- 6White T.J.Bruns T.Lee S.Taylor J.Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics PCR Protocols: A Guide Methods Applicat 1811990315322
- 7Takabayashi M.A coral-specific primer for PCR amplification of the internal transcribed spacer region in ribosomal DNA Mol. Ecol.7199825931
- 8Vollmer S.V.Palumbi S.R.Testing the utility of internally transcribed spacer sequences in coral phylogenetics Mol. Ecol.1392004276327721531568710.1111/j.1365-294X.2004.02265.x · doi ↗ · pubmed ↗