Extracellular vesicles aid in the transfer of long-term associative memory between Caenorhabditis elegans
Monmita Bhar, Tanumoy Nandi, Hari Narayanan, Kamal Kishore, Kavita Babu

TL;DR
This study shows that memory can be transferred between nematodes using extracellular vesicles, which carry learned associations between smells and heat.
Contribution
The paper introduces extracellular vesicles as a novel mechanism for transferring long-term associative memory in nematodes.
Findings
Extracellular vesicles released during training cause loss of attraction to IAA in C. elegans.
Vesicles are specific to the training cues and can transfer memory between species.
The memory transfer mechanism is conserved between C. elegans and C. briggsae.
Abstract
Memory formation is necessary for the survival of animals across phyla. Here, we elucidate the mechanism underlying the formation of long-term associative memory (LTAM) formed by treating Caenorhabditis elegans with a volatile chemoattractant and heat. Previous work has shown that training animals with a paradigm involving heat and isoamyl alcohol (IAA) simultaneously, causes C. elegans to lose their attraction to IAA. In this study, we elaborate on the mechanism behind this LTAM formation and suggest that during training with heat and IAA, C. elegans release extracellular vesicles (EVs) that upon being taken up by the same trained animals or their untrained counterparts causes the organism to lose attraction to IAA. Our data suggests that the vesicles are highly specific to the training paradigms used and differ with differing cues. Finally, we show that this mechanism of transfer of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
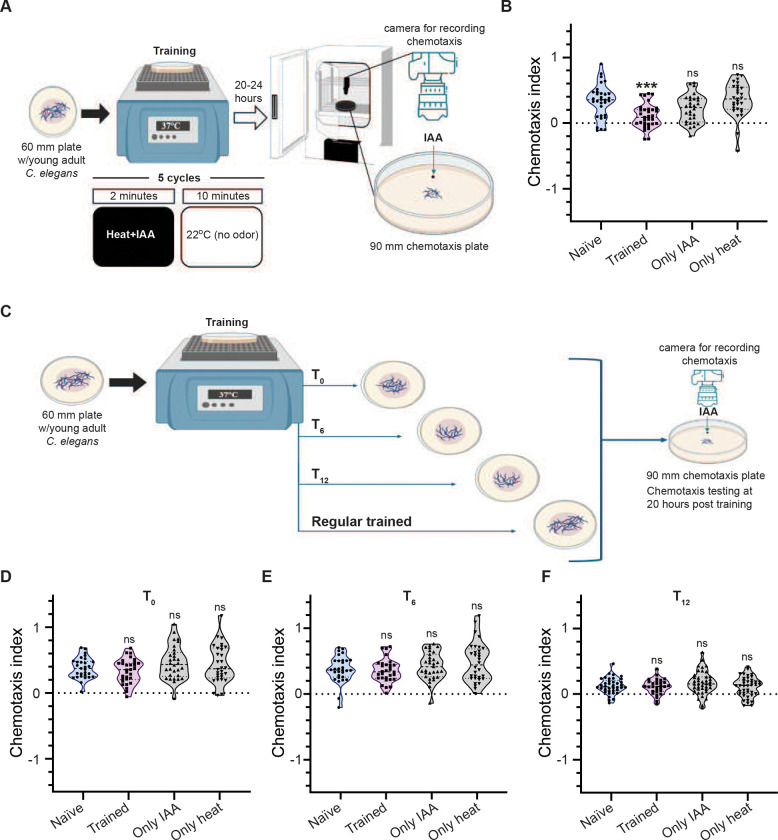 Figure 1
Figure 1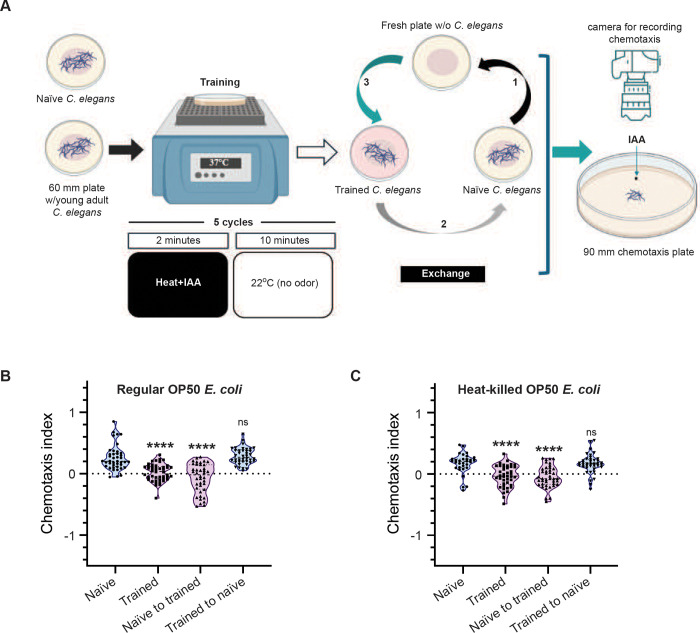 Figure 2
Figure 2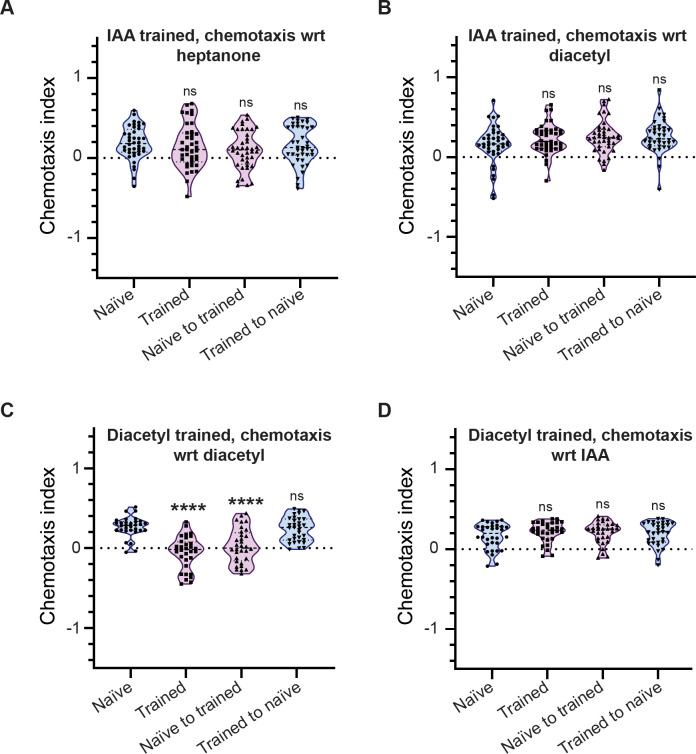 Figure 3
Figure 3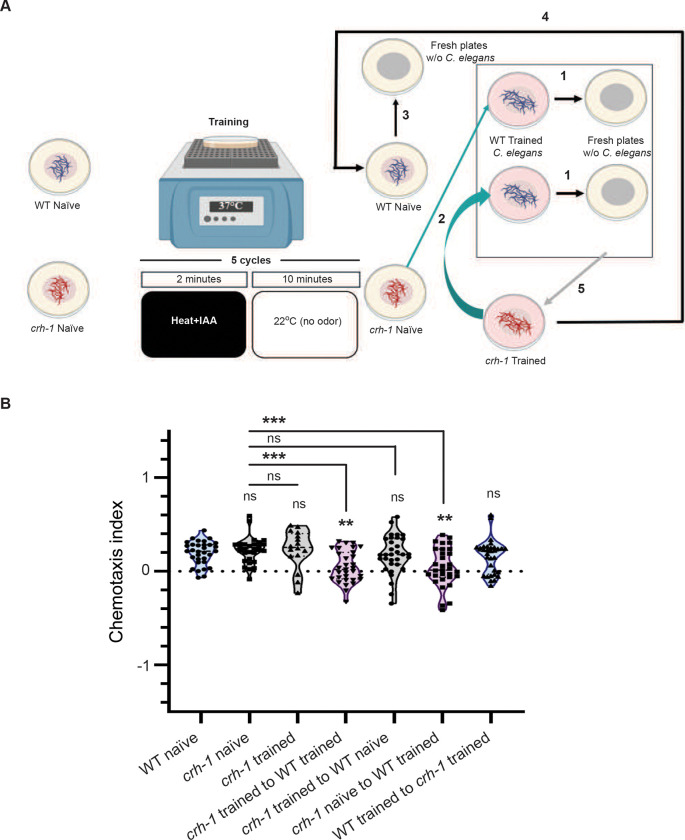 Figure 4
Figure 4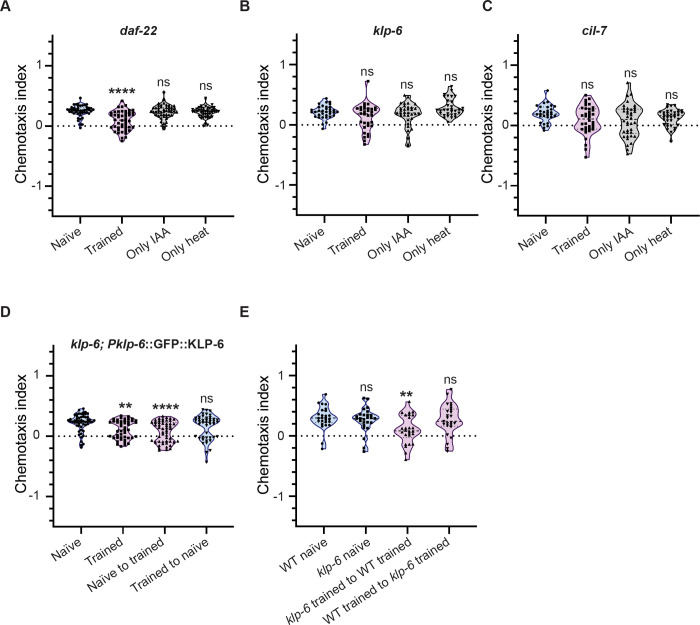 Figure 5
Figure 5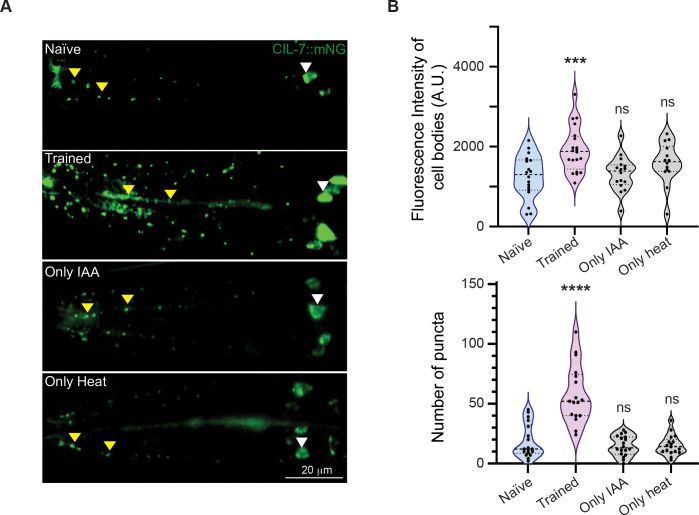 Figure 6
Figure 6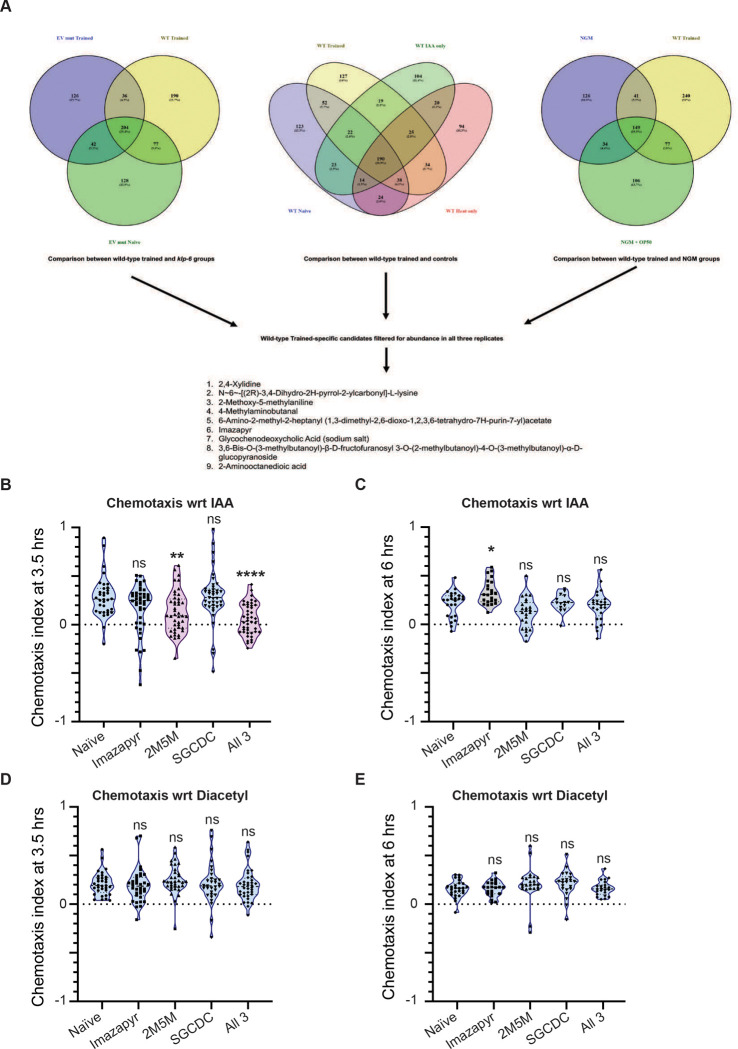 Figure 7
Figure 7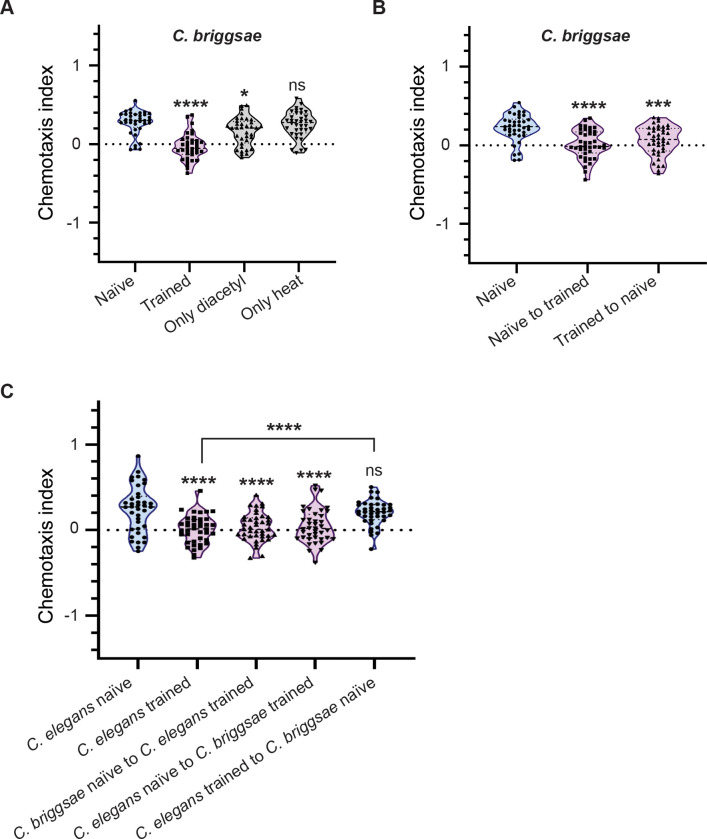 Figure 8
Figure 8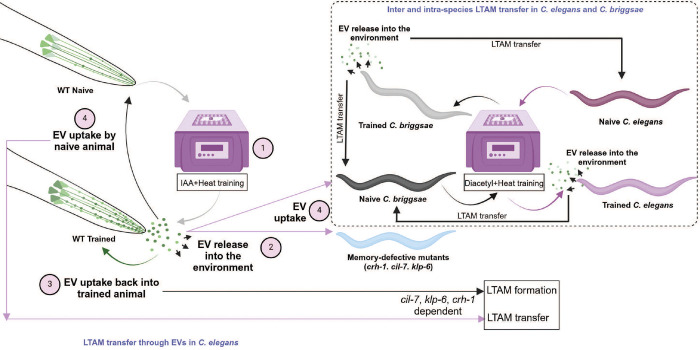 Figure 9
Figure 9- —DBT Janaki Ammal National Women Bioscientist
- —ANRF
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms
Introduction
Native preferences and innate behaviours are essential for the survival of living organisms. Many innate behaviours represent the hardwired neural circuitry put in place during the development of an animal (Bateson & Mameli, 2007; Tierney, 1986). Caenorhabditis elegans has been widely used to study behavioural plasticity because of its ability to learn and modify its innate behaviour (reviewed in (Ardiel & Rankin, 2010)). With a nervous system consisting of 302 neurons, these animals show a variety of robust and adaptable behaviours like chemotaxis and thermotaxis (reviewed in (Ardiel & Rankin, 2010; Hart, 2006)). These behaviours allow C. elegans to move towards food sources, avoid toxic bacteria, and find mates, all of which rely heavily on chemosensation by the animal and innate preferences for certain odours ((Ballestriero et al, 2016) and reviewed in (Ferkey et al, 2021)). For example, under well fed naïve condition, C. elegans are attracted to odorants like isoamyl alcohol (IAA), which can be sensed by the AWC neuron (reviewed in (Bargmann, 2006)). However, this odour to behaviour relationship can be modified if the animal is exposed to periods of starvation along with IAA (Pereira & van der Kooy, 2012). Similarly, neutral concentrations of butanone can be turned attractive when repetitively paired with food. These modified behaviours can persist for up to forty hours post training (Kauffman et al, 2011). The above examples indicate the formation of associative memory, that is brought about as a result of the animal’s interaction with two simultaneous cues.
Associative memory in C. elegans can be classified as short-term or long-term associative memory, depending on how long it can last. Previous studies have shown that pairing cues for a single conditioning period (mass training) forms short-term associative memory (STAM) in C. elegans, that last for around 2 hours. However, multiple conditioning along with periods of rest in between forms long-term associative memory (LTAM), which can last from 20 to 40 hours (Kauffman et al., 2011). LTAM formation in C. elegans has largely been examined by pairing the presence/absence of food (unconditioned stimulus) with a variety of cues (Amano & Maruyama, 2011; Kauffman et al., 2011; Nishijima & Maruyama, 2017). We had shown in a previous study that changes in innate behaviours using the chemoattractive odorant IAA and an aversive heat stimulus leads to loss of attraction to IAA, that lasts more than twenty hours after training, hence giving rise to LTAM formation (Dahiya et al, 2019). Here, we propose a novel mechanism for LTAM formation and show that the effect of training C. elegans with a chemoattractant and heat may lead to the release of training cue specific extracellular vesicles (EVs). Further, these EVs may then be taken up by the same or by untrained counterparts that in turn allow naïve animals to gain LTAM, resulting in impaired attraction to the chemoattractant.
Caenorhabditis elegans can release EVs environmentally through sensory ciliary neurons, which can act as a mode of communication. Previous studies have shown the involvement of EVs in inter-animal communication in the context of mating ((Wang et al, 2014) and reviewed in (Wang et al, 2024a)). There are multiple ciliated EV releasing neurons (EVNs) in C. elegans. 21 of these are male-specific neurons in the head and tail, 6 are inner labial type 2 (IL2) neurons which are present in both males and hermaphrodites (O’Hagan et al, 2017). It has been previously reported that CIL-7, a myristoylated protein regulates EV biogenesis and is known to be an EV cargo. The kinesin-3 protein, KLP-6, is required for the release of EVs into the environment. Both these genes, cil-7 and klp-6, are expressed in the IL2 neurons of hermaphrodite C. elegans (Kahn-Kirby & Bargmann, 2006; Maguire et al, 2015; Morsci & Barr, 2011; Peden & Barr, 2005; Wang et al, 2021). Recent work has also shown that multiple ciliated sensory neurons pack and export ciliary membranes and excess ciliary proteins through EVs (Lobo et al, 2025; Razzauti & Laurent, 2021). Here we report that EVs facilitate the transfer of LTAM from trained animals to both naïve and memory-defective C. elegans. The cAMP response element binding protein (CREB) has been shown to be involved in memory formation across phyla, with mutants in creb1 shown to be defective in long-term memory formation in multiple organisms ((Dash et al, 1990) and reviewed in (Flavell & Greenberg, 2008; Silva et al, 1998)). We show that creb1/crh-1 mutant animals that show no LTAM formation in this IAA and heat-based paradigm (Dahiya et al., 2019), show LTAM from exposure to the extrinsic factors (likely EVs) released by trained wild-type (WT) animals.
These data incentivised us to perform LC-MS experiments to identify the chemicals that may be sent out by trained C. elegans. Using the LC-MS data, we go on to show that a cocktail of three chemicals sent out by C. elegans during the training process, likely through EVs, allows for the animals to show no attraction specifically to IAA recapitulating associative memory without a training paradigm.
This work adds to the growing body of literature describing the role of EVs in multiple processes including cell death, neurodegenerative diseases and therapeutics (Mohamed et al, 2025; Wang et al, 2025; Wiersema et al, 2024; Wright et al, 2025).
Lastly, we report that LTAM can be transferred across species of Caenorhabditis, where C. elegans and C. briggsae can transfer LTAM from trained animals of one species to naïve animals of the other species. Like most previous studies involving LTAM formation in C. elegans and across phyla, we had speculated that the mechanism of LTAM formation as a result of IAA and heat training, was intrinsic to the organism (Dahiya et al., 2019). Contrary to this hypothesis, our findings in this study indicate that LTAM can be transferred from one C. elegans to another through environmentally released extracellular vesicles (EVs), thus challenging the conventional notion of LTAM being intrinsic to an organism.
Materials and Methods
Strain maintenance and transgene generation
All strains were cultured on nematode growth medium (NGM) plates, inoculated with OP50 E. coli bacteria, and maintained under standard conditions at 22°C, following established protocols outlined in previous literature (Brenner, 1974). The N2 Bristol strain served as the standard wild-type (WT) control for comparative analyses across all experimental procedures. A synchronous population of young adult C. elegans was obtained using previously established protocols (Porta-de-la-Riva et al, 2012). The list of strains used in this study is tabulated in Supplementary Table 1 (S1) and primers used for genotyping the strains are tabulated in Supplementary Table 2 (S2). The klp-6 rescue construct Pklp-6::GFP::KLP-6 (gift from Maureen Barr, details in (Wang et al., 2014)) was microinjected in the klp-6 mutant C. elegans to prepare transgenic lines as previously described (C. Mello and Fire 1995; C. C. Mello et al. 1991).
Assay to study long-term associative memory in C. elegans
All assays were performed as previously described in (Dahiya et al., 2019) with minor modifications. Briefly, the training paradigm involves simultaneous exposure of C. elegans or C. briggsae young adults to two cues; heat and a chemoattractant (isoamyl alcohol (IAA)/ Diacetyl (depending on the experiment)), for two minutes followed by ten minutes of rest period at 22°C. This cycle is repeated five times, and the animals are then kept at 22°C for 20 hours (illustrated in Figure 1A). This leads to the formation of long-term associative memory (LTAM), seen in the form of reduced attraction towards the chemoattractant, which can last for as long as 24 hours. LTAM is quantified at 20–24 hours from the time of training, using a readout called the chemotaxis index (CI), where CI= Displacement along IAA, heptanone or diacetyl (depending on the odorant used) gradient/Total distance traversed. The concentrations of the odorants used are as previously published (Dahiya et al., 2019; Zhang et al, 2016). CI is calculated by analysing the videos of the C. elegans undergoing chemotaxis with respect to the chemoattractant. The videos were analysed using FIJI, and an associated plugin, Trackmate (Ershov et al, 2022; Schindelin et al, 2012). Each plot indicates a set of 3–6 replicates with all conditions/genotypes in the graph performed at the same time over multiple days. Each replicate was performed using 8–10 C. elegans. A detailed experimental protocol can be found in the Supplementary section.
Exchange assays
Naïve animals were transferred to an empty plate, followed by trained animals being transferred to the plate that previously had naïve C. elegans. The final step involved transferring the naïve animals to the plate that had had trained C. elegans (illustrated in Figure 2A). This exchange protocol was completed within 30 minutes from the end of training. After exchange, C. elegans were kept in a 22°C incubator and LTAM was tested using the chemotaxis assay 20–24 hours after training. The exchange assays were performed between C. elegans and C. briggsae in a similar manner with naïve C. elegans transferred to plates that had contained trained C. briggsae and naïve C. briggsae transferred to plates that had had trained C. elegans. All plates used for training and transfer were 60 mm in diameter.
Imaging experiments
Freshly trained young adult hermaphrodite C. elegans were used for imaging as previously described with minor modifications (Tikiyani et al, 2018). Briefly, C. elegans were mounted on 1% agarose pads with 34 mg/ml 2,3-Butanedione Monoxime (BDM) solution in M9 medium. Images were acquired as Z-stacks on a Zeiss Apotome microscope using the 63X oil objective within four hours of the training paradigm. The order of imaging of trained and control animals was changed for each of the three replicates. For processing and analysis of images, FIJI (Schindelin et al., 2012) was used to quantify fluorescence intensity of cell body and number of puncta in each condition by selecting specific regions of interest. A threshold was set for analysing particles of each C. elegans to separately quantify cell body intensity and punctal number across the neuronal processes. The summation of mean grey values using “Analyze Particles” from each cell body per worm and the punctal number along each neuronal process was counted using “Analyze Particles”. Both cell body intensity and punctal numbers were plotted as independent graphs. Each replicate had 4–7 C. elegans that were imaged per condition over three replicates and each dot on the plots indicates a single animal.
Liquid Chromatography-Mass Spectrometry (LC-MS)
The protocol used was modified from a previous study (Nikonorova et al, 2022). Sample collection involved four 60 mm plates with approximately 70 C. elegans/plate for each sample. Freshly trained WT animals were washed with M9 buffer and collected as one sample. This was repeated for every group of C. elegans; naïve, trained, only IAA and only heat. Next differential centrifugation was performed. Firstly, the C. elegans suspension was centrifuged at 3,000 g for 15 minutes at 15°C to pellet the animals and bacteria. This supernatant was transferred to fresh tubes and then centrifuged at 10,000 g for 30 minutes at 4°C. This step was repeated thrice. Lastly, the collected supernatant was sent to the IISc LC-MS facility for identification of extrinsic factor(s). Samples were similarly collected from the EV release defective mutants as naïve and trained animals as well as empty plates with and without E. coli. Each sample was processed in triplicate over multiple days.
Chemical supplementation assays
The surface of unseeded NGM plates were supplemented with 300 μl of 100 mM imazapyr (Sigma Aldrich, cat. no. 37877), 2-methoxy 5-methyl aniline (2M5M, Sigma Aldrich, cat. no. 103284) or sodium salt of glycochenodeoxycholic acid (SGCDC, SRL, cat. no. 97971). For the fourth condition, all three chemicals were supplemented together (100 μl each of 100 mM of imazapyr, 2M5M and SGCDC). These plates were freshly seeded with OP50 E. coli bacteria after the chemical solutions had dried. Finally, a synchronized young adult population of naïve C. elegans was transferred (30–35 animals) to each plate. Chemotaxis to IAA and diacetyl was tested at 3.5 hours and 6 hours from the time-point of supplementation. These concentrations and the time for assays were chosen after standardization procedures involving multiple concentrations of the chemicals and multiple time-points at which the assays were performed.
Statistical analysis
All statistical analyses were performed using GraphPad Prism 8. Outliers were identified using Grubbs method (α = 0.05) for all plots. For all plots, the mean values were compared using one-way ANOVA with Dunnett’s correction. The level of significance was set as p ≤ 0.05. The statistical analyses used for each plot is indicated in the figure legends.
Results
Long-term Associative memory in C. elegans requires extrinsic factor(s) released by the animals
We have previously published a training paradigm to study long-term associative memory (LTAM) in wild-type (WT) C. elegans using isoamyl alcohol (IAA) and heat ((Dahiya et al., 2019) and illustrated in Figure 1A). Using this paradigm, we replicated the data showing that unlike naïve animals, their trained counterparts showed a significantly lower chemotaxis index (CI) to IAA (Figure 1B, tracks indicated in Figure S1A). To investigate the underlying molecular mechanism of LTAM formation we planned to perform RNA sequencing experiments using naïve and trained WT animals.
In order to perform this experiment optimally, adult C. elegans had to be synchronized (without eggs and L1 animals) at 20 hours post training for RNA extraction ((Brenner, 1974) and illustrated in Figure S1B)). To remove eggs and L1s from the population, the C. elegans were separated from eggs twice (at 12 and 8 hours) during the twenty-hour resting stage after training and a similar protocol was followed for naïve animals (illustrated in Figure S1B). We initially gently washed the C. elegans and eggs with M9 to remove the C. elegans from the eggs on the plate at both time points. However, upon testing the animals for LTAM after the washes, we found the animals no longer showed LTAM (Figure S1C). This prompted us to pick animals at the standardized 12-hour time point during the 20-hour resting period to circumvent the effects of washing. Again, we found that the animals removed from the plates at the 12-hour time-point no longer showed LTAM (Figure 1F). This prompted us to hypothesize that trained C. elegans may be releasing factor(s) onto the plate that could allow for LTAM formation. In order to test the time-point at which the animals may be releasing unknown factor(s) we tested the animals by picking them from the plate within a few minutes of training (t_0_), 6 hours after training (t_6_), 12 hours after training (t_12_), and keeping some animals in the original plate (illustrated in Figure 1C). We found that only the animals on the original plate showed LTAM formation (Figure 1B), while all other animals lost LTAM associated with heat and IAA (Figures 1D-F).
Our experiments indicated that removal of C. elegans to new plates immediately after training led to a loss of LTAM allowing us to conclude that C. elegans may be releasing unknown factor(s) during training or immediately after training. We then decided to test if these factor(s) could impart LTAM to naïve animals.
Naïve C. elegans on trained plates gain LTAM
To investigate if the factor(s) released by C. elegans during training were indeed required for LTAM formation, we decided to expose naïve C. elegans to the factor(s) released by the animals during training. For this, we performed an exchange assay, where naïve animals were transferred to plates that previously had trained animals, and vice versa (illustrated in Figure 2A). This exchange was performed within 30 minutes of training. Intriguingly, we observed that naïve C. elegans that were transferred to plates that had had trained animals showed LTAM, similar to what we observed in trained control animals (Figure 2B, tracks indicated in Figure S2A). Consistent with our previous results, trained animals that were transferred to plates that had had naïve C. elegans, showed a loss of LTAM (Figure 2B, tracks indicated in Figure S2A). This led us to conclude that these factor(s) released by C. elegans during training are sufficient to induce LTAM in naïve animals.
Next, to make sure that the factor(s) released on the plate were not produced by bacteria on the plate as the animals are well fed with OP50 E. coli throughout their training and resting phases, we performed the same experiments with heat-killed E. coli. The E. coli were heat-killed as previously described (Qi et al, 2017). We observed the same results as previously seen with live E. coli (Figure 2C). To confirm the associative nature of the transferred LTAM, we tested WT C. elegans with heat and IAA in an unpaired assay. In order to perform this experiment, we exposed naïve C. elegans to heat and IAA one after the other as illustrated in Figure S2B and found that in neither case could the animals form LTAM. Further, the plates that had had trained C. elegans did not allow for naïve C. elegans placed on them to gain LTAM (Figures S2C and D).
Our data so far indicates that the associative training paradigm allows C. elegans to release factor(s) onto its surroundings that may be taken up by naïve animals which then go on to show LTAM, hence allowing for the horizontal transfer of LTAM across animals. Next, we wanted to understand the specificity of these released factor(s).
The training cue lends specificity to factor(s) released by trained C. elegans
Our results so far indicate horizontal transfer of LTAM in naïve animals when transferred to a plate that had previously contained trained animals. We wanted to understand the specificity of the released factor(s). Different odours can be sensed by largely distinct neurons in C. elegans (reviewed in (Chou et al, 1996)). IAA is known to be sensed by the AWC neurons, that can also sense heptanone ((Colbert & Bargmann, 1995; Zhang et al., 2016) and illustrated in Figure S3). In order to test the specificity of the secreted factor(s) we took animals trained using the IAA and heat paradigm, along with naïve animals transferred to plates that had contained trained C. elegans and tested their chemoattraction towards heptanone. We observed that these C. elegans showed no LTAM formation with respect to heptanone (Figure 3A). These results indicate a specificity of the released factor(s) that allow for LTAM formation only with respect to the odour used for training.
We next decided to test LTAM formation with respect to another chemoattractive odour sensed by a different neuron. Diacetyl is an odour that is sensed by the AWA neuron ((Sengupta et al, 1996; Sengupta et al, 1994) and illustrated in Figure S3). We trained animals with heat and IAA and found that trained animals and naïve animals transferred to plates that had contained trained animals did not show any defects in chemotaxis with respect to diacetyl (Figure 3B). Our previous work has shown that the heat and chemoattractant based training paradigm may be used with diacetyl and heat in a process similar to that performed with IAA and heat (Dahiya et al., 2019). We next trained the animals with heat and diacetyl and found that they showed LTAM towards diacetyl (Figure 3C). Further, naïve animals placed on a plate that had had trained C. elegans showed LTAM towards diacetyl (Figure 3C). This experiment indicates that a release of factor(s) occurs independent of whether the odour is IAA or diacetyl; the nature of these factor(s), however, appears to be extremely specific to the odorant used during training. Finally, we trained animals with diacetyl and tested them with IAA and found no LTAM formation towards IAA (Figures 3D). These data together indicate a high degree of specificity of the factor(s) released for allowing LTAM formation only with respect to the cue present in the training paradigm.
We next wanted to study the role of these factor(s) in animals that are known to be defective in memory formation.
LTAM defective C. elegans show LTAM from exposure to plates that had contained trained WT animals
A plethora of literature has implicated CREB1 (cAMP Response Element Binding protein 1) in long-term memory formation across phyla ((Bartsch et al, 1998; Timbers & Rankin, 2011; Yin et al, 1995; Yin et al, 1994) and reviewed in (Flavell & Greenberg, 2008; Johannessen et al, 2004; Silva et al., 1998; Yin & Tully, 1996)). We have shown that the mutants of the ortholog of mammalian creb1, crh-1 in C. elegans, show LTAM defects in the heat and IAA paired assay (Dahiya et al., 2019). We decided to test if crh-1 mutants allowed for LTAM formation when transferred to a plate that had contained trained WT animals (illustrated in Figure 4A). Our data indicates that crh-1 mutants that basally show no LTAM, show LTAM when transferred to a plate which had had WT trained animals. We also found that WT trained animals transferred to a plate that had contained trained crh-1 mutant C. elegans do not show LTAM (Figure 4B and tracks indicated in Figure S4). These data allowed us to conclude that crh-1 mutants may have defects in the synthesis or release of the factor(s) that allow for LTAM formation but appear to be able to uptake these factor(s) to show LTAM.
We next wanted to gain insight into the mechanism of secretion of the factor(s) during training of C. elegans.
Extracellular vesicles are required for LTAM formation
A well-studied mechanism of social behaviour in C. elegans is through secreted small molecule signalling by ascarosides (reviewed in (Yang et al, 2023)). Pre-exposure to different ascarosides can change the chemotaxis of C. elegans to different odours (Wu et al, 2023). We decided to test for LTAM formation in the daf-22 mutant C. elegans that are defective in ascaroside biosynthesis (Golden & Riddle, 1985). We found that daf-22 mutants showed no defects in LTAM formation (Figure 5A). These data indicate that the secreted factor(s) may not be daf-22 dependent. Apart from ascarosides, sensory signalling allowing for behaviours like mate sensing through chemoattraction requires extracellular vesicle (EV) release from C. elegans (reviewed in (Wang et al., 2024a)). We next tested mutants in EV biogenesis (cil-7) and release (klp-6) for LTAM formation (Wang et al., 2021). We found that both mutants showed no LTAM formation (Figure 5B and C). In order to further confirm the role of EVs in the process of LTAM, we attempted to rescue the loss of LTAM phenotype in klp-6 mutants by expressing KLP-6 specifically in the IL2 neurons where KLP-6 has been shown to be expressed in hermaphrodites (Peden & Barr, 2005; Wang et al, 2024b). We observed that expressing KLP-6 in the mutant klp-6 animals rescued the loss of LTAM phenotype where both trained C. elegans as well as naïve animals transferred to plates that had contained trained animals now showed LTAM formation (Figure 5D). We next decided to test if klp-6 mutants allowed for LTAM formation when transferred to a plate that had trained WT animals. We found that trained klp-6 mutants transferred to a plates which had had trained WT animals showed LTAM, while trained WT animals transferred to plates that had contained trained klp-6 mutant C. elegans did not show LTAM (Figure 5E and tracks indicated in Figure S5). These data allow us to conclude that EV release may be the basis for LTAM formation in a chemoattractant and heat-based associative training paradigm.
We next investigated changes in EV expression in the nervous system upon training C. elegans with heat and IAA.
Changes in EV biogenesis marker expression upon heat and IAA training
Previous work has elegantly shown that an EV cargo, CIL-7, is released from the IL2 neurons into the external region around C. elegans (Wang et al., 2024b). We tested the same line for changes in fluorescence expression in the IL2 neurons and found that the cil-7 (my61[cil-7::mNG])I; him-5(e1490) line showed increased fluorescence in the IL2 cell bodies and an increase in fluorescent puncta along the IL2 neuronal processes in trained WT animals when compared to the control C. elegans that were either naïve or trained with only IAA or only heat (Figures 6A and B). With these data indicating the involvement of EVs in LTAM transfer. We next decided to perform a Liquid Chromatography-Mass Spectrometry (LC-MS) to study the differences in the released products upon heat and IAA training when compared to control animals.
A mixture of three chemicals mimics memory formation in C. elegans
LC-MS was performed using WT C. elegans as well as klp-6 mutant animals. Multiple LC-MS experiments were performed, each in triplicate as indicated in Figure 7A. This experiment allowed us to find 9 consistent hits that were only present in the heat and IAA trained C. elegans plates (Figure 7A and S6). These data suggest that multiple ions are differentially released onto the plates upon heat and IAA training. These ions may allow for LTAM formation in C. elegans. Based on the availability of chemicals, we supplemented the plates of naïve animals with three chemicals from our candidate list – imazapyr, 2M5M and SGCDC. Each molecule was supplemented on NGM plates onto which naïve animals were placed. To understand their cumulative role, we also supplemented all three chemicals together.
we observed that imazapyr or SGCDC alone did not alter the chemotaxis towards IAA in treated C. elegans, whereas 2M5M supplementation led to the loss of attraction to IAA, seen as a lower chemotaxis index when compared to naïve animals This loss of attraction to IAA phenotype was more pronounced in C. elegans treated with all three molecules (Figure 7B, and track images in S7A). This effect was seen at the 3.5 hours’ time-point from supplementation and is lost by 6 hours after supplementation (Figure 7C), indicating that these molecules allow for memory formation, although not LTAM formation. This could be because of the absence of the other six molecules from our candidate list, along with other cargo including nucleic acids that are usually known to be carried by EVs released into the environment (Nikonorova et al., 2022). We also checked the chemotaxis of naïve and animals trained with IAA and heat towards Diacetyl at the 3.5- and 6-hour time-points.
At both time points we observed no loss of attraction towards Diacetyl (Figures 7D and E). As controls we performed the training and testing protocols with WT C. elegans where testing was performed at 3.5 and 6 hours after training. The aversion to IAA was consistently seen at both time points (Figures S7B and C). These results establish our findings that specific factors released in response to training, could lead to specific loss of attraction towards the odorant used in the training paradigm.
Finally, we wanted to test if LTAM formation is conserved across Caenorhabditis species. In order to perform this experiment, we investigated LTAM formation in Caenorhabditis briggsae.
Caenorhabditis show interspecies transfer of LTAM
We first tested the C. briggsae strain for LTAM formation to heat and IAA and found that C. briggsae were surprisingly not attracted to IAA even in naïve conditions (Figure S8A and B). This precluded us from studying LTAM formation in C. briggsae using the IAA and heat based associative paradigm*.* However, since C. briggsae naïve animals behaved like C. elegans trained animals (Figure S8B), we went on to transfer C. briggsae naïve animals to a plate that had contained C. elegans trained animals and found that there was no change in C. briggsae chemotaxis index. We also transferred trained C. elegans to a plate that had contained naïve C. briggsae (exchange assay) and found that these animals behaved like naïve C. elegans (Figure S8B). These data could not allow us to make any conclusions other than the fact that C. briggsae was not attracted to IAA in the naïve condition unlike C. elegans.
We next tested if C. briggsae was attracted to diacetyl. We found that C. briggsae was both attracted to and formed LTAM with heat and diacetyl (Figure 8A). We went on to transfer C. briggsae naïve animals to a plate that had contained C. briggsae trained animals and vice versa. We found that C. briggsae naïve animals acquired LTAM from the plate that had contained trained C. briggsae, and surprisingly the trained C. briggsae did not lose LTAM from being on a plate that had had naïve C. briggsae (Figure 8B). These data indicate that a similar mechanism may allow for LTAM transfer in C. briggsae as is seen in C. elegans, however the LTAM appears to be more stable in C. briggsae when compared to C. elegans. Finally, we tested if interspecies transfer of LTAM could occur. For this experiment we transferred naïve C. briggsae to a plate that had contained trained C. elegans and transferred naïve C. elegans to a plate that had contained trained C. briggsae and found that both sets of animals showed LTAM (Figure 8C). We also transferred trained C. elegans to a plate that had contained naïve C. briggsae and unsurprisingly found no LTAM formation (Figure 8C).
Together, our data indicates the role of extracellular vesicles in LTAM formation and its horizontal transfer in C. elegans along with its possible role in this phenomenon across species.
Discussion
In this study we report three novel findings that have broad implications. First, C. elegans form long-term associative memory (LTAM) through environmentally released extracellular vesicles (EVs) from the IL2 neurons when the animals undergo training with heat and a chemoattractant. Further, these EVs appear to be cue specific and allow for LTAM formation only with respect to the cue used in the initial training paradigm. Second, LTAM can be transferred to naïve as well as memory-defective mutant animals possibly through EVs. Third, there appears to be conservation of this behaviour across Caenorhabditis species with EVs from C. elegans being able to elicit LTAM formation in C. briggsae and vice versa. These findings are illustrated in Figure 9. Below, we discuss the significance of these findings.
Novel mechanism for horizontal transfer of LTAM
Previous studies have shown that a form of non-associative memory can be transferred from one Aplysia to another by injecting RNA extracted from the trained animal into the naïve counterpart (Bedecarrats et al, 2018). More recent studies have shown that learned avoidance to pathogenic bacteria can be transferred from trained to naïve C. elegans through the Cer1 retrotransposon (Moore et al, 2021). However, there appear to be no studies showing the organic transfer of LTAM from one organism to another without active communication between the animals. In this study, we show that C. elegans when trained using an IAA and heat-based associative learning paradigm may release extrinsic factor(s). Removal of trained animals from these plates results in loss of LTAM, indicating that these factor(s) are responsible for the process of LTAM formation. Our results also show that these factor(s) are likely released during training. Naïve animals exposed to the factor(s) released by trained animals also show LTAM formation, similar to that seen in the original trained animals. This work uncovers a novel mechanism for the horizontal transfer of LTAM through secreted EVs from one Caenorhabditis elegans to another, irrespective of whether the animals have been trained with the associative learning paradigm. Further, our work suggests that EVs released due to training appear to carry extremely specific cargo with respect to the odorant cue used during training. The cargo may dynamically change with changing training cues allowing for cue specific LTAM formation.
LTAM formation in memory defective animals
Apart from allowing for LTAM formation in naïve animals, we show that the factor(s) (EVs) secreted by wild-type (WT) C. elegans also show LTAM in memory defective creb1/crh-1 and EV biogenesis/cargo defective animals using multiple previously described mutants (Dahiya et al., 2019; Maguire et al., 2015; Wang et al., 2021; Wang et al., 2024b). These animals gain LTAM to the specific isoamyl alcohol (IAA) cue by being placed on a plate that had had WT animals trained with heat and IAA. This intriguing finding may allow for a better understanding of the factor(s) released and the mechanism of EV uptake/action that appear to be intact in these mutants that show defects in LTAM formation. Future work may allow for a better understanding of the involvement of CREB/CRH-1 in the observed LTAM formation and transfer phenomenon.
A chemical cocktail that allows C. elegans to behave like it has been trained with heat and IAA
We found that a cocktail of Imazapyr, 2-methoxy 5-methylaniline (2M5M) and Glycochenodeoxycholic acid (SGCDC) elicits a loss of attraction specifically to IAA, albeit for a short time-span of upto 3.5 hours. How these chemicals may act throws open interesting future experiments to understand memory and chemosensation in Caenorhabditis. Imazapyr is known to be a non-selective herbicide which controls vegetation, especially hard-to-control perennial grass by inhibiting the acetolactate synthase (ALS) enzyme. It can disrupt protein synthesis and interfere with the synthesis of DNA and cell growth (Dersch et al, 2016). ALS is the first enzyme in the pathway for the synthesis of branched-chain amino acids (Dezfulian et al, 2017). 2M5M is an organic compound that is largely used in the production of dyes and pigments of inks and paints. It is also used as a catalyst in reactions like nitration of aromatic compounds. (https://chem-iso.com/18630). Finally, SGCDC, usually found as a sodium salt is a bile salt found in the liver, generated from chenodeoxycholic acid and glycine. It has been shown to inhibit autophagosome formation lysosomal function leading to apoptosis of human hepatocyte cells (Lan et al, 2020). However, to our knowledge, none of these compounds have been studied in the context of C. elegans. It would be interesting to test more of the compounds found in our LC-MS experiments to try and get a better understanding of what C. elegans release during heat and IAA training.
LTAM transfer between Caenorhabditis briggsae and Caenorhabditis elegans
We show that C. briggsae, a relative of C. elegans, with a divergence of tens of millions of years (Butler et al, 1981) also show LTAM formation through released factor(s) that are likely to be EV dependent. We found that C. briggsae do not appear to be attracted to IAA in a manner similar to C. elegans, likely due to the high divergence in the olfactory pathways of these animals (Jovelin et al, 2009). However, both animals showed attraction towards diacetyl. We show that LTAM can transfer from trained C. briggsae to naïve C. elegans and trained C. elegans to naïve C. briggsae, indicating a conserved mechanism of LTAM transfer across Caenorhabditis, likely through EVs.
The one interesting difference we found was that unlike C. elegans, C. briggsae does not seem to lose LTAM when removed from the plates that they were trained on. This could be because of two possibilities; C. briggsae release the extrinsic factor(s) continuously for a period of time, long enough for LTAM consolidation, or these factor(s) (EVs) are stable and functional for longer periods of time in C. briggsae. Further studies may allow for parsing out these differences between the two species.
Our work leads to multiple outstanding questions that may allow for a better understanding of the process of LTAM transfer across organisms. We briefly discuss some of these unanswered questions below.
Neuronal circuitry allowing for EV release by IL2 neurons for LTAM formation
Studies have shown that CIL-7, a myristoylated protein regulates EV biogenesis along with being an EV cargo and the kinesin-3 protein KLP-6 is required for the release of EVs. Both these genes, cil-7 and klp-6, are expressed in the IL2 neurons in C. elegans hermaphrodites (Kahn-Kirby & Bargmann, 2006; Maguire et al., 2015; Morsci & Barr, 2011; Peden & Barr, 2005; Wang et al., 2021). Here we report that cil-7 and klp-6 mutant animals are LTAM defective, uncovering a previously unreported role of the IL2 neurons in LTAM formation.
This raises important questions in understanding the neuronal circuitry involved. Different neurons are responsible for sensing different odorants in C. elegans. Our experiments indicate that animals trained with IAA (sensed by the AWC neurons) and heat versus diacetyl (sensed by the AWA neurons) and heat release EVs with different cargo causing cue-specific LTAM formation. Understanding the neuronal circuitry between chemosensory neurons and IL2 neurons may allow for a better understanding of the specificity of LTAM formation.
Dynamic EV cargo for cue-specific LTAM
C. elegans show LTAM formation and transfer, specific to the odorant cue used during training. For examining the extrinsic factor(s) released by trained animals, we performed LC-MS to identify candidate molecules present exclusively on the plates of trained animals (Figure 6B). One of these molecules 4-methylaminobutanal, might be of potential interest. IAA is chemically 3-methylbutanol, and a biological pathway for the conversion of 3-methylbutanol to 4-methylaminobutanal may exist. The steps would include oxidation of 3-methylbutanol to 3-methylbutanal through an alcohol dehydrogenase (Illikoud et al, 2018), followed by a transamination step producing 3-methylbutylamine (Encyclopedia of Sensors and Biosensors, 2023). This intermediate could potentially result in 4-methylaminobutanal through additional reactions. We have shown that a cocktail of three of these LC-MS hits allows for C. elegans to lose attraction to IAA, how these and other molecules function would allow for more insight into what EV cargoes may consist of as well allow for a better understanding of memory towards IAA. Since we see specificity of LTAM formation to the odorant used in the training paradigm, it is plausible that different odorants may allow for release of cue-specific cargo. This in turn may give rise to the specificity of the observed LTAM phenomenon.
Uptake of EV cargo for LTAM transfer and a role for RNAs in this process
Transfer of LTAM from one C. elegans to another entails factor(s) (EVs) released by trained animals, and its uptake by naïve animals. This uptake could occur through a variety of mechanisms. Is the cargo released through EVs retained within the EV structure after release into the environment? Are the individual components of vesicles dispersed onto the plates? Further, does the uptake mechanism involve only neurons/glia or could other tissues like cuticles/hypodermis be involved? Understanding these aspects of the uptake mechanism may shed more light on the process of LTAM transfer across Caenorhabditis species.
RNA has been implicated in memory in multiple model organisms. The role of different types of non-coding RNAs like micro-RNAs (miRNAs) and Piwi-interacting RNAs (piRNAs) have been explored and studied in the context of long-term non-associative memory in Aplysia (Fiumara et al, 2015; Rajasethupathy et al, 2012; Rajasethupathy et al, 2009). Further, it has been shown in Aplysia that memory formed from long-term sensitization (non-associative) can be transferred from trained to naïve animals (Bedecarrats et al., 2018). More recently, multiple studies in have defined the role of non-coding RNAs in transgenerational memory as well (reviewed in (Miska & Rechavi, 2021)). Studies have also shown that small non-coding RNAs expressed in Pseudomonas strains allow for the transgenerational inheritance of avoidance memory in C. elegans (Kaletsky et al, 2020; Legue et al, 2022; Sengupta et al, 2024; Seto et al, 2024). Additionally, Moore et al, 2021 has shown that this avoidance memory can also be horizontally transferred from one animal to another (trained to naïve C. elegans) through virus-like particles encoded by the Cer1 retrotransposon (Moore et al., 2021). Multiple studies have shown that extracellular vesicles from C. elegans carry proteins, non-coding RNAs, miRNAs, lipid nanodomains, enzymes and other chemicals (Nikonorova et al., 2022; Russell et al, 2020). Collectively, these findings suggest the involvement of RNA in learning and memory and may allow for the hypothesis that non-coding RNAs or miRNAs could be involved in LTAM uptake from trained to naïve animals through EVs.
In summary, we report a novel mechanism of LTAM transfer through environmentally released EVs. This, to the best of our knowledge, is the first report of organic transfer of long-term associative memory between organisms and across species.
Supplementary Material
This is a list of supplementary files associated with this preprint. Click to download.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Amano H, Maruyama IN (2011) Aversive olfactory learning and associative long-term memory in Caenorhabditis elegans. Learn Mem 18: 654–66521960709 10.1101/lm.2224411 PMC 3187929 · doi ↗ · pubmed ↗
- 2Ardiel EL, Rankin CH (2010) An elegant mind: learning and memory in Caenorhabditis elegans. Learn Mem 17: 191–20120335372 10.1101/lm.960510 · doi ↗ · pubmed ↗
- 3Ballestriero F, Nappi J, Zampi G, Bazzicalupo P, Di Schiavi E, Egan S (2016) Caenorhabditis elegans employs innate and learned aversion in response to bacterial toxic metabolites tambjamine and violacein. Sci Rep 6: 2928427384057 10.1038/srep 29284 PMC 4935850 · doi ↗ · pubmed ↗
- 4Bargmann CI (2006) Chemosensation in C. elegans. Worm Book: 1–2910.1895/wormbook.1.123.1PMC 478156418050433 · doi ↗ · pubmed ↗
- 5Bartsch D, Casadio A, Karl KA, Serodio P, Kandel ER (1998) CREB 1 encodes a nuclear activator, a repressor, and a cytoplasmic modulator that form a regulatory unit critical for long-term facilitation. Cell 95: 211–2239790528 10.1016/s 0092-8674(00)81752-3 · doi ↗ · pubmed ↗
- 6Bateson P, Mameli M (2007) The innate and the acquired: useful clusters or a residual distinction from folk biology? Dev Psychobiol 49: 818–83118023000 10.1002/dev.20277 · doi ↗ · pubmed ↗
- 7Bedecarrats A, Chen S, Pearce K, Cai D, Glanzman DL (2018) RNA from Trained Aplysia Can Induce an Epigenetic Engram for Long-Term Sensitization in Untrained Aplysia. e Neuro 5 Brenner S (1974) The genetics of Caenorhabditis elegans. Genetics 77: 71–9410.1523/ENEURO.0038-18.2018 PMC 596204629789810 · doi ↗ · pubmed ↗
- 8Butler MH, Wall SM, Luehrsen KR, Fox GE, Hecht RM (1981) Molecular relationships between closely related strains and species of nematodes. J Mol Evol 18: 18–237334524 10.1007/BF 01733207 · doi ↗ · pubmed ↗