Generation of six4 -nlsRFP: a red somatic gonadal nuclear marker for live imaging Drosophila gonadogenesis
Everette Rhymer, Lauren Anllo

TL;DR
Researchers created a red fluorescent marker to track cell movements in fruit fly gonad development using live imaging.
Contribution
A new red fluorescent nuclear marker for somatic gonadal precursor cells in Drosophila was developed for live imaging.
Findings
A red fluorescent nuclear marker was generated for live tracking of somatic gonadal precursor cells.
The marker enables visualization of cell interactions during Drosophila gonadogenesis.
The tool complements existing green fluorescent reagents for cytoskeletal and adhesion studies.
Abstract
Intrinsically fluorescent tissue labels are valuable to live image cell behaviors during development. Fluorescent markers of Drosophila gonadal cells have elucidated dynamic cellular interactions that shaped our conception of cell biology. Many green fluorescent protein (GFP) reagents are available to visualize cytoskeletal dynamics, extracellular matrix, and cell adhesion proteins. To facilitate use of these reagents while studying somatic gonadal development, we generated a somatic gonadal precursor (SGP) intrinsic red fluorescent nuclear marker. This tool can track SGP movement live, facilitating the study of somatic cell interactions required for gonadogenesis.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
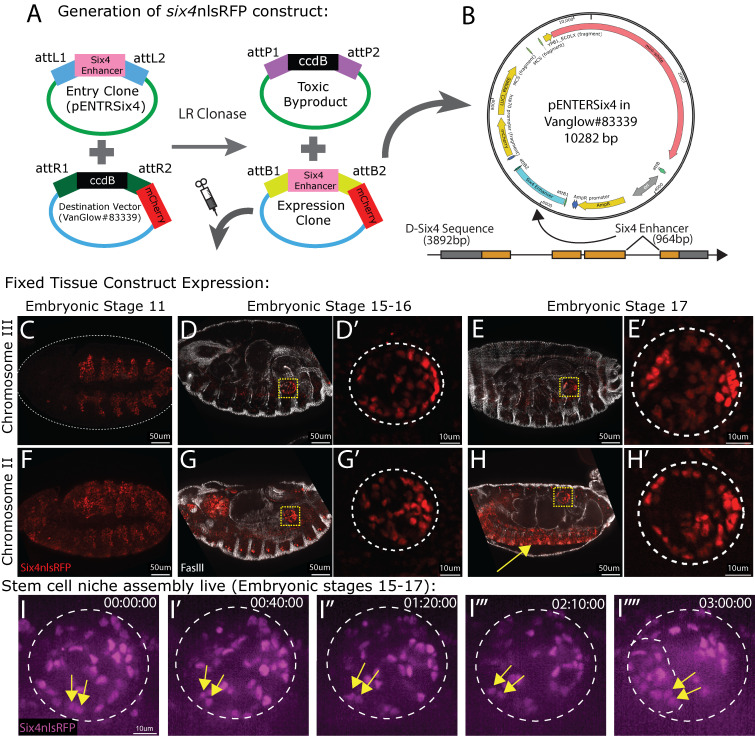 Figure 1
Figure 1|
|
|
|
|
|
y[1] w[67c23]; P{y[+t7.7]=CaryP}attP1 |
Bloomington
|
|
|
y[1] w[67c23]; P{y[+t7.7]=CaryP}attP2 |
Bloomington
|
|
|
Invitrogen, Catalog # C404006 | |
|
|
|
|
|
Vanglow#83339 |
mCherry::Renilla(nls) upstream of ccdB flanked by two AttR cloning sites. Available at Addgene. Catalog # 83339. | |
|
pENTR/D-Topo entry vector |
Invitrogen, Catalog # K240020. | |
|
|
CAC CCA GCA AAG ACC GTG AGT TG |
(Clark et al., 2006) D-
|
|
|
GTT GGA TCC ATT GCC ATC CAG TTG |
(Clark et al., 2006) D-
|
|
|
|
|
|
Rabbit polyclonal anti RFP |
Abcam |
ab62341; RRID:AB_945213 |
|
Mouse monoclonal anti FasciclinIII |
Developmental Studies Hybridoma Bank |
DSHB-7G10 RRID:AB_528238 |
|
Alexafluor Secondary Antibodies (Cy5 conjugated) (minimally cross reactive IgG generated in donkey) |
Jackson ImmunoResearch laboratories |
Catalog # 715-175-151 |
|
Cy3 Affinipure Secondary Antibodies |
Jackson ImmunoResearch laboratories |
Catalog # 711-165-152 |
|
|
|
|
|
Gateway LR Clonase II Enzyme mix |
Thermofisher Scientific |
Catalog # 11791020 |
|
Ampicillin |
Thermofisher Scientific |
Catalog # 11593027 |
|
Kanamycin |
Thermofisher Scientific |
Catalog # 11815032 |
|
|
|
|
|
Leica M165FC |
Leica |
N/A |
|
mCherry Filter set ET560/40x; ET630/75m |
Leica 10450195 |
N/A |
|
Achromat 1.6x objective |
Leica 10450163 |
N/A |
|
Olympus IX83 microscope |
Olympus/Evident |
N/A |
|
Crest X-Light V3 spinning disk confocal microscope |
89 North |
N/A |
|
Silicone oil objective 60x, 1.3 NA |
Evident 1-U2B7122 |
N/A |
|
Zeiss AxioImager.Z1 microscope equipped with ApoTome.2 and X-Cite Xylis LED illuminator |
Zeiss |
N/A |
|
C-Apo 40x water objective 1.2 NA |
Zeiss |
N/A |
|
Plan-Apo 20x objective 0.8 NA |
Zeiss |
N/A |
- —National Institutes of Health (United States)https://ror.org/01cwqze88
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDevelopmental Biology and Gene Regulation · Wnt/β-catenin signaling in development and cancer · Nuclear Structure and Function
Description
Introduction
Fluorescent proteins are commonly used to label subcellular structures during live imaging, which has enabled a way to study cell biological processes *in vivo * during development (Chudakov et al., 2005, 2010; Day & Davidson, 2009). A prominent model that has defined many foundational developmental and reproductive processes is the *Drosophila * embryonic gonad (Heemskerk & DiNardo, 1994; Kunwar et al., 2006; Le Bras & Van Doren, 2006; Santos & Lehmann, 2004; Sheng et al., 2009; Tran et al., 2000; Yamashita Yukiko et al., 2003). At embryonic stage 15, the gonad is a spherical arrangement of germ cells (GCs) encysted by somatic gonadal precursor cells (SGPs). The development of intrinsically fluorescent protein labels has enabled live imaging to describe coalescence of these gonadal cell types, and testis stem cell niche morphogenesis (Anllo et al., 2019; Clark et al., 2006; Lin et al., 2020; Sano et al., 2012; Warder et al., 2024). Libraries of intrinsic labels to visualize cytoskeletal regulators, adhesion proteins, and extracellular matrix components often employ green fluorescent protein (GFP) (Nagarkar-Jaiswal et al., 2015). To use these libraries while simultaneously visualizing formation of somatic gonadal structures including the niche, the construction of red fluorescent SGP labels is crucial. The field previously lacked a tool to intrinsically label SGPs with a red fluorophore. This limitation prevented red fluorescent labeling of SGPs while simultaneously using Gal4/UAS drivers in other gonadal tissues. Here, we used molecular cloning to develop a six4 nlsRFP genetic fly line that intrinsically labels SGP nuclei with an mCherry RFP. This line enables the tracking of somatic gonadal cell migratory paths during live imaging, while simultaneously labeling other proteins of interest in separate fluorescent channels. We show that our six4 nlsRFP lines can be used to visualize SGPs in both fixed tissue immunostaining and live imaging experiments at various stages of embryonic gonadogenesis.
Results
Transgenic expression of six4nls RFP in fixed tissue:
Embryos expressing the six4nls RFP sequence were fixed and immunostained against RFP to visualize the SGP reporter. The adhesion protein Fasciclin III was also immunostained to highlight specific structures in the embryo, including the epidermis and visceral muscle. Embryos with six4 nlsRFP on chromosome 3 (see Methods) showed nuclear expression in a subset of mesodermal cells during mid-embryogenesis ( Fig. 1C ), and more restricted expression later in SGPs during gonad coalescence and formation of the spermatogonial niche ( Fig. 1C- 1E'). Interestingly, embryos with the six4 nlsRFP sequence inserted at the docking site on chromosome 2 showed additional RFP expression in the nervous system, along with expression in early mesoderm and later somatic gonadal cells ( Fig. 1F- H'). Thus, while both insertions allow for clear delineation of SGPs, the positional insertion on Chromosome III enables more specific SGP visualization.
Transgenic expression of six4nls RFP in live imaging:
Once expression was confirmed in fixed tissue experiments, we used our chromosome 3 inserted six4 nlsRFP-expressing embryos to live image assembly of the spermatogonial stem cell niche. The spermatogonial niche derives from a subset of SGPs that cluster together at the anterior of the male gonad during Stages 15-17 of embryogenesis (Le Bras & Van Doren, 2006; Anllo et al., 2019). Endogenous expression of the construct was sufficient to visualize the gonad and mount embryos for live imaging at embryonic stage 15 ( Fig. 1I ). After 3 hours of live imaging, at embryonic stage 17, we observed a niche assembled at the anterior ( Fig. 1I ''''). The six4 nlsRFP construct allowed visualization and tracking of individual niche cells migrating to the anterior ( Fig. 1I- 1I'''', yellow arrows). We show here that the six4 nlsRFP construct can be used to trace the migratory paths and dynamic movements of individual SGPs live during niche assembly.
Discussion
During embryogenesis, the gonad undergoes rapid morphological changes (Clark et al., 2006; Jenkins et al., 2003; Sano et al., 2012). Live imaging the late embryonic *Drosophila * testis has revealed cell migratory paths and cytoskeletal dynamics that could not have been predicted from fixed tissue image analysis (Anllo et al., 2019; Anllo & DiNardo, 2022; Warder et al., 2024). To investigate these behaviors, precise intrinsic labeling of individual cells is required along with separate visualization of extracellular matrix interactors, cell-cell adhesive proteins, and cytoskeletal dynamics. Here, we generated a red fluorescent marker, six4 nlsRFP, to intrinsically mark somatic gonadal precursor (SGP) nuclei, enabling tracking of individual cells while employing GFP-tagged libraries to separately label cell interactions and behaviors. The six4 nlsRFP reporter does not require use of Gal4/UAS induction, thus saving Gal4/UAS for other simultaneous manipulations in SGPs, germ cells, or other extrinsic tissues.
Despite lower quantum yield of red fluorescent proteins compared to green fluorescent variants (Wall et al., 2015), six4 nlsRFP has high intrinsic fluorescence for visualization with a stereofluorescent dissecting microscope. This feature enables precise orientation of embryos mounted for live imaging with the gonad positioned directly adjacent to the coverslip. One copy of the six4 nlsRFP construct provides sufficient fluorescence despite the internal depth of the gonad more than 50 microns from the epidermis. Additionally, chromosomes with the six4 nlsRFP construct are homozygous viable, presenting the opportunity for increased brightness with two construct copies.
The chromosome 3 attP2 insertion of six4 nlsRFP shows more restricted expression than the chromosome 2 attP1 insertion, which has ectopic expression in the nervous system. Positional effects have been documented for attP sites that vary across tissues (Markstein et al., 2008), and strong position effects were found for attP1 in the adult nervous system when an enhancer trap vector was inserted (Peiffer et al., 2010). These position effects might explain the additional RFP expression detected in the embryonic nervous system with the attP1 insertion.
Individual SGPs can be tracked over time using the six4 nlsRFP label. During late-stage gonadogenesis, SGPs encyst germ cells with long cellular extensions that contact one another in dense cellular arrangements (Clark et al., 2006; Sano et al., 2012). This arrangement poses a challenge for tracking individual cell positions with cell cortex labels. The nuclear fluorescence with six4 nlsRFP overcomes this limitation. Our future work will employ this tool to track individual SGPs while using separate GFP fluorophores to measure their dynamic F-actin nucleation patterns during interactions with the germline, extracellular matrix, and other SGPs.
This tool facilitates study of cell behaviors that are not detectable in individual cells over time in fixed immunostaining experiments. Overall, this reagent enhances our ability to investigate dynamics in the developing somatic Drosophila gonadal cells, which has implications for advancing studies of stem cell-niche biology and organogenesis.
Methods
Generation of six4nlsRFP expressing flies:
To generate a six4 nlsRFP DNA sequence, we used two genomic plasmids, the Gateway vector pENTR (ThermoFisher Scientific) and Vanglow#83339 (AddGene, Janssens et al., 2017). We used Gateway cloning to insert the * Six4 * enhancer sequence into pENTR, flanked by 2 attL sites ( Fig. 1A ). Oligonucleotides previously described were used to amplify the * Six4 * enhancer encoded in its third intronic region (Clark et al., 2006). The Vanglow#83339 plasmid contained a ccdB sequence, encoding a cell death casette, flanked by 2 attR sites and followed by a red fluorescent protein (RFP) sequence, mCherry, with a nuclear localization signal. pENTR and contained a kanamycin-resistance gene (KanR), while Vanglow#83339 encoded ampicillin-resistance (AmpR) sequence, allowing for selective growth when transformed in bacteria. Plasmids were amplified using bacterial transformation and validated using restriction enzyme digestion and DNA sequencing (Plasmidsaurus).
An LR clonase reaction was performed to induce recombination at the attL and attR sites between these plasmids (ThermoFisher Scientific). This reaction placed the * Six4 * enhancer sequence upstream of the mCherry RFP sequence in VanGlow#83339 ( Fig. 1B ). Plasmids were then amplified using bacterial transformation and grown selectively on agar plates containing 100 ug/uL ampicillin. Bacterial cultures containing six4nlsRFP were purified and sequenced (Plasmidsaurus), which validated that the LR clonase reaction had worked successfully. The sequence was then injected into flies containing an attP1 docking site on chromosome 2R (55C4) (BDSC#8621), and fly lines containing an attP2 docking site on chromosome 3L (68A4) (BDSC#8622) through PhiC31 integrase-mediated transformation (Bateman et al., 2006). BestGene was used for injections.
Immunostaining and imaging:
Embryos were collected on grape agar plates overnight, then dechorionated using 50% bleach for approximately 2 minutes. Embryos were transferred to a glass screw cap vial with equal parts heptane and 4% paraformaldehyde (PFA) to fix for 20 minutes on a rocking nutator. PFA was then aspirated and replaced with an equal amount of 100% methanol, and the glass vial was vigorously shaken for 1 minute to remove the vitelline membrane. Two washes of 100% methanol were performed to wash away excess PFA and heptane. Embryos were then washed in 50/50 methanol/PBS to rehydrate, then placed in 0.1% phosphate-buffered saline with Triton (PBST) prior to blocking. Tissue was blocked using 4% normal donkey serum (NDS). Primary antibody staining was performed overnight at 4C, and all secondary antibodies were used at 3.75ug/ml (Alexa 488, Cy3, Alexa 647; Molecular Probes; Jackson Immunoresearch) for 2 hours at room temperature. Hoechst 33342 (Sigma) was used at 0.2ug/ml for 5 minutes to stain DNA. We used rabbit antibody against RFP at 1:500 (Abcam 62341) and mouse antibody against Fasciclin III at 1:20 (DSHB 7G10). Fixed embryos were imaged using a Zeiss AxioImager.Z1 with an ApoTome.2, X-cite Xylis fluorescent light source, and Axiocam 705 mono camera, using a 20x 0.8 N.A. lens, or a 40x 1.2 N.A. lens.
in vivo live imaging:
Embryos were prepared for live imaging as described previously by Ong and colleagues (Ong et al., 2019). Embryos were collected overnight (~16hrs) at 25C on a grape agar plate and dechorionated the next morning in a nytex basket using 50% bleach. Embryos were then selected for embryonic stage 15 using gut morphology to determine age according to Campos Ortega and Hartenstein (Campos-Ortega & Hartenstein, 1997). Embryos were mounted dorsolaterally to a slide using double-sided tape dissolved in heptane to create an adhesive solution and then covered in halocarbon 27 oil. Two 18x18mm coverslips were glued to the slide on both sides of the embryos to create a bridge. A 22x30mm coverslip was then glued to the bridge coverslips to allow space for the embryos. Slides were imaged using an Olympus IX83 microscope with a Crest X-Light V3 spinning disk, using a 60x oil immersion objective.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Anllo Lauren Di Nardo Stephen 202241 Visceral mesoderm signaling regulates assembly position and function of the Drosophila testis niche Developmental Cell 5781534-580710091023.e 510.1016/j.devcel.2022.03.00935390292 PMC 9050945 · doi ↗ · pubmed ↗
- 2Anllo, L., Plasschaert, L. W., Sui, J., & Di Nardo, S. (2019). Live imaging reveals hub cell assembly and compaction dynamics during morphogenesis of the Drosophila testis niche. Developmental Biology , 446 (1), 102–118. https://doi.org/10.1016/j.ydbio.2018.12.014 10.1016/j.ydbio.2018.12.014PMC 641074630553808 · doi ↗ · pubmed ↗
- 3Bateman, J. R., Lee, A. M., & Wu, C. (2006). Site-Specific Transformation of Drosophila via ϕC 31 Integrase-Mediated Cassette Exchange. Genetics , 173 (2), 769–777. https://doi.org/10.1534/genetics.106.056945 10.1534/genetics.106.056945 PMC 152650816547094 · doi ↗ · pubmed ↗
- 4Campos-Ortega, J. A., & Hartenstein, V. (1997). The Embryonic Development of Drosophila melanogaster . Springer Berlin Heidelberg. https://doi.org/10.1007/978-3-662-22489-2
- 5Chudakov, D. M., Lukyanov, S., & Lukyanov, K. A. (2005). Fluorescent proteins as a toolkit for in vivo imaging. In Trends in Biotechnology (Vol. 23, Issue 12, pp. 605–613). https://doi.org/10.1016/j.tibtech.2005.10.005 10.1016/j.tibtech.2005.10.00516269193 · doi ↗ · pubmed ↗
- 6Chudakov, D. M., Matz, M. V., Lukyanov, S., & Lukyanov, K. A. (2010). Fluorescent proteins and their applications in imaging living cells and tissues. In Physiological Reviews (Vol. 90, Issue 3, pp. 1103–1163). https://doi.org/10.1152/physrev.00038.2009 10.1152/physrev.00038.200920664080 · doi ↗ · pubmed ↗
- 7Clark, I. B. N., Boyd, J., Hamilton, G., Finnegan, D. J., & Jarman, A. P. (2006). D-six 4 plays a key role in patterning cell identities deriving from the Drosophila mesoderm. Developmental Biology , 294 (1), 220–231. https://doi.org/10.1016/j.ydbio.2006.02.044 10.1016/j.ydbio.2006.02.04416595131 · doi ↗ · pubmed ↗
- 8Day, R. N., & Davidson, M. W. (2009). The fluorescent protein palette: Tools for cellular imaging. Chemical Society Reviews , 38 (10), 2887–2921. https://doi.org/10.1039/b 901966 a 10.1039/b 901966 a PMC 291033819771335 · doi ↗ · pubmed ↗