Isolation and selection of indigenous chicken-derived Bacillus subtilis strains as potential probiotic alternatives to antibiotics against Gram-negative enteropathogens
Hai Vu Phan, Hung Hoang Son Pham, Lai Huu Ngo, Na Thi Tran, Dung Thi Ho, Khuong Dinh Thuy Nguyen, Liem Ngoc Tran, Hoa Xuan Nguyen

TL;DR
This study identifies two Bacillus subtilis strains from chicken feces that can fight harmful bacteria in poultry, offering a potential probiotic alternative to antibiotics.
Contribution
The study isolates and evaluates indigenous Bacillus subtilis strains as effective and stable probiotic candidates against poultry enteropathogens.
Findings
Six B. subtilis strains showed antibacterial activity against E. coli and S. typhimurium.
B. subtilis BSn5 survived freeze-drying for two months and colonized the chicken intestine.
Strains H1 and BSn5 were identified as the most promising probiotic candidates.
Abstract
The increasing antibiotic resistance in poultry pathogens presents a significant public health risk, leading to the exploration of effective alternatives in broiler feed, particularly probiotics such as Bacillus subtilis. This study aimed to isolate B. subtilis strains from indigenous chicken feces that can inhibit Escherichia coli and Salmonella typhimurium strains, common causes of diarrhea in poultry. Bacillus strains were isolated from chicken feces and screened for antibacterial activity using an agar well diffusion assay. Bacillus strains were identified via 16S rRNA gene sequencing. Their probiotic potential was assessed through in vitro assays measuring extracellular enzyme production, adhesion properties, and resilience to acidic and bile salt conditions. Freeze-drying techniques were applied to evaluate strain viability and stability. In vivo studies determined the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
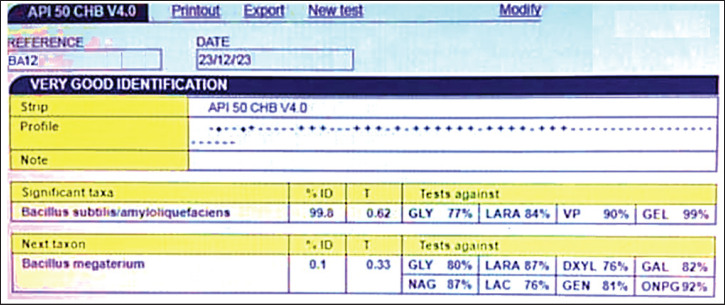 Figure 1
Figure 1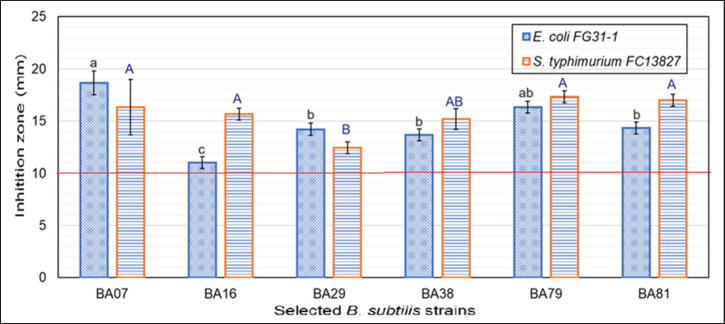 Figure 2
Figure 2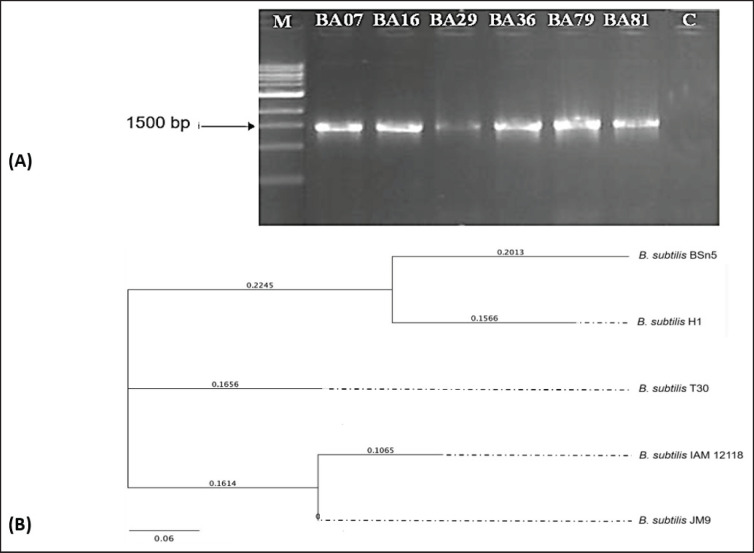 Figure 3
Figure 3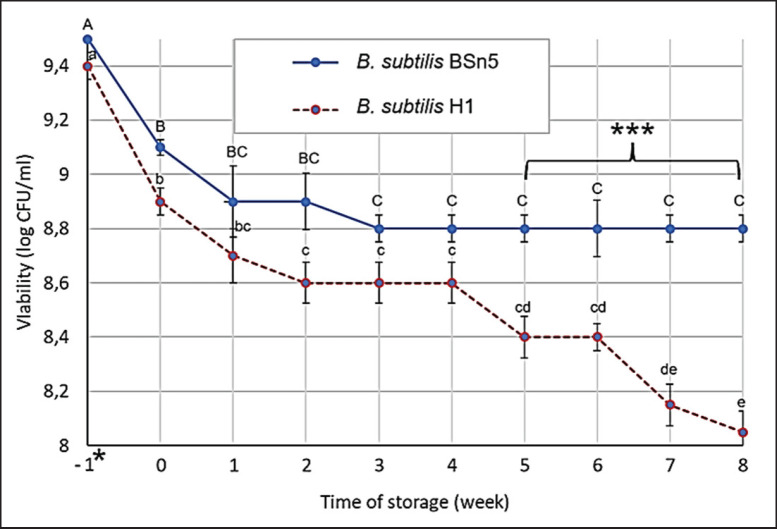 Figure 4
Figure 4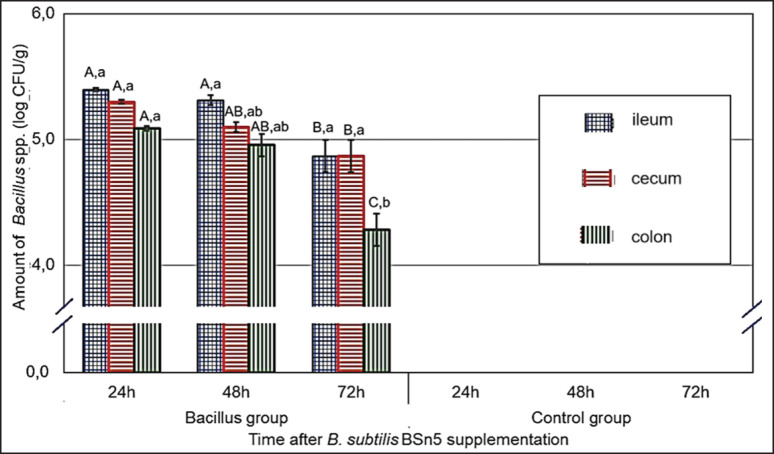 Figure 5
Figure 5| Number of strains | Similarity ratio (%) | Isolate symbol | Species |
|---|---|---|---|
| 2 | 99.9 | BA07, BA91 |
|
| 1 | 99.8 | BA102 |
|
| 2 | 99.3 | BA12, BA56 |
|
| 1 | 99.3 | BA15 |
|
| 2 | 98.7 | BA38, BA92 | |
| 1 | 98.0 | BA16 |
|
| 1 | 98.0 | BA81 | |
| 1 | 99.5 | BA59 |
|
| 2 | 98.5 | BA074, BA69 |
|
| 2 | 90.6 | BA82, BA112 |
|
| Isolate | Similarity ratio (%) | GenBank ID of reference strain | |
|---|---|---|---|
| BA07 | 100 | ||
| BA16 | 100 | NR112116 | |
| BA29 | 100 | ||
| BA38 | 100 | ||
| BA79 | 100 | ||
| BA81 | 100 |
| Properties | BA07 | BA29 | BA79 | BA16 | BA81 |
|---|---|---|---|---|---|
| Extracellular enzyme production (U/ml) | |||||
| amylase | 0.43a ± 0.02 | 0.36ab ± 0.032 | 0.34ab ± 0.02 | 0.34ab ± 0.02 | 0.30b ± 0.02 |
| protease | 1.78c ± 0.25 | 0.93d ± 0.04 | 3.62b ± 0.27 | 5.80a ± 0.31 | 0.73d ± 0.04 |
| lipase | 0.08a ± 0.01 | 0.02c ± 0.03 | 0.02c ± 0.02 | 0.08a ± 0.004 | 0.04bc ± 0.02 |
| Hydrophobicity (%) | 55.31b ± 2.08 | 60.24ab ± 7.56 | 66.98a ± 4.88 | 54.83b ± 1.55 | 47.17c ± 1.41 |
| Co-aggregation (%) | |||||
| with | 38.87a ± 5.15 | 43.98a ± 6.44 | 39.71a ± 5.15 | 41.45a ± 4.01 | 29.86b ± 4.18 |
| with | 42.13a ± 4.01 | 31.55b ± 6.19 | 34.52b ± 6.26 | 28.97b ± 4.42 | 30.36b ± 4.01 |
| Self-aggregation (%) | 62.31a ± 2.22 | 56.74a ± 3.34 | 44.72b ± 2.37 | 35.58b ± 4.5 | 23.21c ± 3.74 |
| Acid tolerance (%) | |||||
| at pH = 2 | 83.91 ± 3.61 | 85.3 ± 2.92 | 83.33 ± 2.53 | 84.48 ± 2.11 | 84.78 ± 2.24 |
| at pH = 3 | 89.06 ± 2.24 | 90.95 ± 3.33 | 87.22 ± 2.84 | 90.17 ± 2.01 | 87.78 ± 3.28 |
| Bile salt tolerance (%) | |||||
| at 0.3% | 96 ± 1.8 | 95.5 ± 1.4 | 96.8 ± 0.9 | 94.3 ± 1.2 | 85.1 ± 1.0 |
| at 0.5% | 82.4 ± 2.1 | 82.7 ± 1.6 | 82.6 ± 1.4 | 83.5 ± 2.1 | 78.2 ± 2.3 |
| Total high probiotic properties* | 5 | 5 | 3 | 4 | 1 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Bacteriophages and microbial interactions · Viral gastroenteritis research and epidemiology
Introduction
Currently, Escherichia coli and Salmonella spp. are prevalent Gram-negative bacteria that cause gastrointestinal diseases in poultry, leading to substantial economic losses for the poultry industry [1]. These pathogens, particularly, have zoonotic potential, posing a serious public health threat through foodborne transmission [2]. At present, there are no effective vaccines to protect chickens against these infections due to the diversity of bacterial serotypes and limited cross-protection [3]. Both E. coli and Salmonella spp. are listed by the WHO as pathogens for which antibiotic susceptibility testing is recommended before treatment [4]. The rising issue of antibiotic resistance, fueled by the overuse of antibiotics in livestock, has resulted in widespread resistance in E. coli [5] and Salmonella spp. [6], diminishing antibiotic efficacy, altering the gut microbiota, and negatively impacting health. Consequently, the search for antibiotic alternatives, such as probiotics, has become imperative.
Probiotics comprise live microorganisms and their metabolites, which, when administered orally, confer health benefits to the host. In addition to producing antimicrobial compounds such as organic acids and bacteriocins, probiotics can compete with pathogenic bacteria by adhering to the intestinal epithelium, thereby preventing pathogen colonization, disrupting cell-to-cell communication, and inhibiting biofilm formation and virulence [7]. Unlike antibiotics, probiotics do not induce resistance, leave no residues, and are environmentally friendly [8]. Among various probiotics, bacteria from the Bacillus genus are widely used in both humans and animals due to their antimicrobial compound production and spore-forming ability, which provides a dual advantage for survival in diverse environments [9].
Notably, Bacillus subtilis strains are garnering interest as feed additives due to their beneficial impact on animal health, as these bacteria can form spores that withstand harsh conditions, such as high pH, acidic environments, and elevated temperatures [10]. This resilience probably allows B. subtilis to survive the extreme conditions of the gastrointestinal tract, enhancing its stability during production, storage, and feed formulation while also extending shelf life and increasing gastrointestinal stability in animals. In commercial probiotic products, the efficacy of B. subtilis varies by strain, and the specific characteristics of these strains are often not fully documented. This lack of detail can lead to mislabeling of products and may result in the inclusion of strains that contain harmful factors, such as toxins, posing risks to both animal and public health.
Moreover, numerous researchers emphasize that probiotics isolated from the host organism offer a better chance of survival and efficacy compared to those derived from other sources, as they are more likely to overcome the challenges associated with introducing foreign bacteria [11]. This study is the first to focus on the isolation and selection of potential B. subtilis from free-ranging chicken feces to combat Gram-negative pathogens (E. coli and Salmonella spp.) responsible for diarrhea in broilers, based on their antimicrobial capabilities and probiotic properties.
Materials and Methods
Ethical approval
In this study, all procedures related to the care, housing, and slaughtering of experimental chickens were conducted following the standards and approvals of the Animal Ethics Advisory Committee, Hue University, Vietnam (Approval No: HUVNO39).
Isolation of Bacillus spp.
Bacillus spp. was isolated from the feces of healthy, semi-free-range indigenous broiler chickens (Ga Kien) that had not been supplemented with probiotics. The chickens were provided with a healthy and enjoyable diet that meets all their nutritional needs while allowing them to express their natural foraging behaviors. The isolated strains were screened using serial dilution and heat shock treatment in a water bath (Unitronic^®^ 300) at 80°C for 10 min, as described by Cazorla et al. [12], to eliminate vegetative cells, retaining only spore-forming strains for Bacillus isolation. Subsequently, 0.1 ml of each sample was streaked onto nutrient agar plates (Merck, Germany) and incubated for 24 h at 37°C. Single colonies, selected based on morphological differences, were transferred to fresh plates until consistent monocultures were obtained after three rounds of subculturing. Morphologically distinct colonies were purified, examined for Gram staining, and subjected to biochemical tests, including lactose, glucose, mannitol, and xylose fermentation; starch hydrolysis; gas production; motility; indole; urease; catalase; H_₂_S production; and Voges–Proskauer, as outlined in the Manual of Systematic Bacteriology. Pure colonies were then cultured in LB broth, with the pure cultures preserved in Eppendorf tubes supplemented with 40% glycerol and stored at –80°C.
Species identification of Bacillus using API 50 CHB Kit
The API 50 CHB kit (bioMérieux, France), consisting of 50 biochemical tests, was used to identify Bacillus strains at the species level. Following incubation, the bacteria were introduced into the kit wells. After 24–48 h of incubation, results were interpreted based on color changes: positive (red to yellow) or negative (no color change). The outcomes of the 50 reactions were input to the API web software to identify the species and determine the similarity percentage.
Evaluation of antibacterial activity against chicken gastrointestinal pathogens
Pathogenic bacteria: Escherichia coli FG31-1 and Salmonella typhimurium FC13827 (GenBank IDs: CP142680.1 and MN704402.1), carrying the virulence genes invA and stn, were isolated from the diarrhea feces of chickens suspected of E. coli or Salmonella spp. infections. These isolates were first identified based on their phenotypic and biochemical characteristics, followed by confirmation through 16S rRNA gene sequencing (Table 2). These strains were maintained at the Microbiology Laboratory, Faculty of Animal Science and Veterinary Medicine, Hue University of Agriculture and Forestry. The antibacterial activity of selected Bacillus strains against these pathogens was assessed using the agar well diffusion method. Muller Hinton Agar (MHA, Thermo Fisher Scientific) plates with 4 mm height and 100 µl of the bacterial suspension were overlaid with 0.5 OD 630 pathogenic bacterial suspension. After allowing the suspension to settle for 15–20 min, six wells (with a diameter of 6 mm and spaced 30 mm apart) were made on the MHA plates by marking positions along a straight line and creating the wells at those marked points. Each well received 100 μl of overnight LB broth culture of the selected Bacillus strains (adjusted to 0.5 OD 630). The plates were allowed to diffuse for one hour at 4°C and then incubated at 37°C for 24 h. Evaluation formula: Diameter (D, mm) = Diameter of inhibition zone (DIZ). DIZ of ≥ 10 mm was considered indicative of antibacterial activity [13].
Identification of suspected B. subtilis strains via gene sequencing
Suspected B. subtilis strains were identified through 16S rRNA gene sequencing. Genomic DNA from a single colony of each isolated strain was extracted using the Bacterial DNA Kit TM (Zymo Research, Cat. No. D6005, USA) targeting a 1,500 bp gene fragment. Amplification of the 16S rRNA gene was performed using universal primers with the sequences 27F (5’-AGA GTT TGA TCM TGG CTC AG-3’) and 1492R (5’-TAC GGY TAC CTT GTT ACG ACT T-3’) [14]. The thermal cycling conditions were 1 cycle at 94°C for 5 min, followed by 35 cycles at 94°C for 1 min, 55°C for 1 min, 72°C for 1 min, and a final extension at 72°C°C for 15 min. PCR products were purified using the ZR-96 DNA Sequencing Clean-up Kit (Zymo Research, USA) according to the manufacturer’s instructions and sequenced using the CLC 7 sequencing system (QIAGEN, Germany). The PCR products were stained with SYBR Green for visualization under ultraviolet light and electrophoresed on an agarose gel, with a DNA ladder included for size estimation of the PCR bands. After Sanger sequencing and BLAST (Thermo Fisher Scientific) analysis, the data were compared with the GenBank database of the NCBI for Bacillus species identification.
The phylogenetic tree of B. subtilis strains was constructed using Geneious Prime software (Biomatters, New Zealand), using the neighbor-joining method. The branch lengths correspond to the ladder with a mutation rate of 0.06 mutations/site.
Probiotic properties analysis of B. subtilis
Amylase activity was measured qualitatively in starch agar and quantitatively by the DNS (dinitrosalicylic acid) method as described by Sharif et al. [15]. Amylase hydrolyzes in starch agar, which then reacts with DNS to form a colored complex, measured at 540 nm. Protease activity was quantified as directed by Zhang et al. [16], using the casein plate method with absorbance read at 660 nm. Lipase activity was determined according to Bharathi and Rajalakshmi [17], using triglycerides as the substrate. Lipase catalyzes the hydrolysis of triglycerides to release glycerol and free fatty acids, which were quantified by titration or colorimetric methods.
The hydrophobicity assay, based on Krausova et al. [18] with minor adjustments, assessed microbial adhesion to xylene. Bacillus subtilis cultures were grown in LB medium, centrifuged, washed with Ringer’s solution, and resuspended to ~0.08 OD at 600 nm. Xylene was added and vortexed, and the aqueous phase was evaluated at 600 nm after separation. The percentage hydrophobicity was calculated as follows: [(OD0—OD) / OD0] × 100, where OD0 and OD represent the optical densities before and after mixing with xylene.
Auto-aggregation and co-aggregation assays were conducted as described by Mallappa et al. [19]. Bacillus subtilis strains were centrifuged at 8,500 rpm for 10 min and then resuspended in phosphate-buffered saline and incubated at 37°C for 4 h. A 0.2 ml sample of the suspension was taken, and OD at 600 nm was measured before and after incubation. Auto-aggregation was measured using the formula 1—[At / Ao] × 100, where Ao and At are the initial and final optical densities. For co-aggregation, B. subtilis was prepared similarly. An E. coli suspension in BHI medium was standardized to approximately 1 × 10_8_ CFU/ml. Bacillus suspension was mixed with E. coli suspension (1 ml/1 ml) and shaken for 10 sec and then allowed to settle. A control containing only 2 ml of the bacterial suspension was prepared. The absorbance at 600 nm was measured after 5 h of incubation at 37°C. Co-aggregation was measured using the formula: (OD600 (x) + OD600 (y)—OD600 (x + y)) / (OD600 (x) + OD600 (y)) × 100.
Acid and bile tolerance of B. subtilis was assessed following the method by Mallappa et al. [19]. The B. subtilis strains were cultured overnight and suspended in 10 ml LB with pH adjusted to 2 and in 50 ml LB containing 0.3% bile salts (Himedia, India). The inoculum size was standardized to 0.5 McFarland (~1.5 × 10_8_ CFU/ml). The tubes were incubated at 37°C for 3 h, with 300 μl samples taken at 0, 1, 2, and 3 h for growth dynamics measurements using a spectrophotometer at 600 nm. Concurrently, 100 µl samples were taken to determine viable cell counts by the standard plate count method.
Evaluation of viability of selected B. subtilis strains during storage
Freeze-drying of bacterial strains cultured on skim milk was conducted in a medium enriched with maltodextrin and trehalose. Twenty-four-hour cultures of each isolated strain (20 ml each) were transferred to sterile, disposable polypropylene containers (60 ml capacity). The samples were initially stored at –20°C for 72 h, followed by freeze-drying under vacuum conditions at a constant pressure of 63 Pa for 96 h, with a shelf temperature of 30°C on a lyophilized (Mactech MST50GD, Meta Company, Vietnam). The freeze-dried samples of the five bacterial strains were then ground and thoroughly mixed in equal proportions to produce a prototype feed additive.
The stability of the five freeze-dried strains and the feed additive was determined by comparing bacterial counts on plates immediately after freeze-drying and after 60 days of storage at 4°C. The samples were placed in sterile, sealed containers to evaluate their storage stability. The number of viable cells or spores was measured on agar plates both before and after freeze-drying and throughout two months of storage. The samples of each preparation form were rehydrated, serially diluted, and plated. Survival rates were evaluated based on Lo Curto et al. [20] with some modifications by exposing the probiotic powder to an HCl solution (pH 1.5, at 37°C for 2 h) and subsequently to a 0.05 M phosphate buffer (pH 7.4) at 37°C. All preparations were stored over silica gel in closed glass containers (desiccators) at 48°C for 2 months.
Assessment of bacterial viability in the chicken gut
A total of 36 one-day-old 3FViet chicks were divided into two groups, each containing three cages (6 chicks per cage). The experimental group received 1 ml of preparation containing 10^9^ CFU/ml *B. subtilis *BSn5, while the control group was given orally 1 ml of distilled water by syringe. The chicks were housed in iron cages (0.9 × 0.5 × 0.5 m) under continuous light and maintained at 35°C throughout the experiment (1–3 days of age). Before the experiment, the cage system and floors were sterilized using heat (gas torch) and disinfectant (Povidone 10%).
The chicks were fed a diet of locally sourced ingredients such as rice bran, cornmeal, peanut meal, and soybean meal, meeting the standards set by the Ministry of Agriculture and Rural Development, Vietnam (10 TCN 661-2005). Before use, feed and water were sterilized by UV light (300 µW-s/cm², 30 min) and provided ad libitum to the chicks. At 24, 48, and 72 h, three chicks from each group (one per cage) were randomly selected and sacrificed, and samples were collected from the ileum, cecum, and colon. These samples were washed and heat-treated at 80°C for 20 min to eliminate vegetative cells and other bacteria. Finally, the counts of B. subtilis spores were determined at the specified time points. Results were expressed as the average number of spores per gram in the ileum, cecum, and colon.
Statistical analysis
All assays were performed in triplicate, and data were expressed as mean ± standard deviation or percentage. Bacterial counts were converted to log_10_ CFU/ml. Statistical analysis was conducted using IBM SPSS software (version 22), with significance assessed by one-way ANOVA followed by Tukey’s post hoc test, and results were statistically significant at α = 0.05.
Results and Discussion
Isolation and identification of Bacillus spp.
Isolation of Bacillus spp.
A total of 121 bacterial isolates with morphological characteristics suggestive of *Bacillus *strains were obtained with the following characteristics: Gram-positive, opaque white, dry colonies with irregular edges, cell size greater than 3 µm, and the presence of centrally located endospores that did not distort the cell shape.
Biochemical characterization
Among the 121 isolates, 100% were catalase-positive, 58.7% (71/121) were VP-positive, 73.2% (52/71) were amylase-positive, 52.5% (40/52) were able to grow at 50°C, and 52.5% (21/40) were cellulase-positive. These 21 isolates were selected for further analysis.
Identification using the API CH50B kit
Following biochemical characterization, 21 isolates exhibited characteristics consistent with B. subtilis. To confirm their taxonomic identity, these isolates were further analyzed using the API CH50B kit (Table 1, Fig. 1). The results revealed that 16 isolates were identified as B. subtilis/B. amyloliquefaciens, with a high degree of similarity (> 90%).
Selection of B. subtilis strains based on their antibacterial properties on chickens
Grethel Milián et al. [20] reported that Bacillus strains C-31, C-34, and E-44 produced antimicrobial substances that completely inhibited the growth of indicator strains such as Aerobacter, Staphylococcus, Klebsiella, Proteus, Listeria innocua, L. monocytogenes, S. aureus 29737, Klebsiella 130300, S. cholermidis 12228, and P. vulgaris 13315.
The observed antibacterial activity can be attributed to the production of bacteriocins, antimicrobial peptides that suppress the growth of harmful bacteria by disrupting cell membrane integrity or interfering with protein or DNA synthesis [9]. Additionally, some Bacillus strains produce antibiotics such as polymyxin and gramicidin, which exhibit broad-spectrum antibacterial activity, including against E. coli and Salmonella spp. [22]. This antibacterial activity, coupled with competitive exclusion mechanisms, whereby probiotic strains compete with pathogens for nutrients and attachment sites, effectively prevents pathogen colonization in the gut.
The assessment of antibacterial activity is a crucial step in identifying potential probiotic candidates for antibiotic replacement. The antagonistic activity of 21 B. subtilis isolates against two Gram-negative enteropathogens were evaluated. As shown in Figure 2, six *B. subtilis *strains (28.57%) exhibited inhibitory activity against both pathogens, with DIZ ≥ 10 mm. Eight strains (38.09%) inhibited only one of the two pathogens, while seven strains (33.33%) did not exhibit inhibitory activity against either pathogen (data was not shown). Among the six strains with inhibitory activity against both pathogens, strain BA07 displayed the most potent activity, with significantly larger DIZ (16.4–18.2 mm) compared to the other strains (11.3–17.4 mm) (p < 0.05). Strains BA29, BA38, BA79, and BA81 exhibited relatively similar DIZ, ranging from 12.6 to 17.4 mm. S. typhimurium FC13827 appeared to be more susceptible to the B. subtilis strains than E. coli FG31-1, as evidenced by larger DIZ for 4 out of 6 B. subtilis strains.
Reading the identification results using the API software.
Tentative identification of B. subtilis by gene sequencing
Agarose gel electrophoresis revealed that all selected strains yielded a 1,500 bp amplicon, consistent with the expected size of the 16S rRNA gene. The obtained 16S rRNA gene sequences were deposited in GenBank, and a phylogenetic tree was constructed based on these sequences.
The six Bacillus strains, designated as BA7, BA16, BA29, BA36, BA79, and BA81, were identified as B. subtilis strain H1, B. subtilis strain IAM 12118, B. subtilis strain BSn5, B. amyloliquefaciens strain G341, B. subtilis ssp. strain JM9, and B. subtilis strain T30, respectively, through 16S rRNA gene sequencing (Table 2, Fig. 3). These sequences were compared against the NCBI database, revealing high similarity (100%) to sequences with the following accession numbers: CP026662.1, NR112116.1, CP002468.1, CP011686, MT605298.1, and CP011051, respectively. Five identified B. subtilis strains were used for further studies.
Selection of B. subtilis strains based on probiotic properties
The ability of probiotic strains to produce extracellular enzymes is a crucial probiotic trait that aids digestion and reduces fecal waste [23]. All selected B. subtilis strains exhibited the capacity to produce key digestive enzymes, including amylase (starch hydrolysis), protease (protein hydrolysis), and lipase (lipid hydrolysis).
Strain BA07 displayed the highest amylase activity (0.43 U/ml), which was significantly higher (p = 0.016) than that of strain BA81 (0.30 U/ml). Strain BA16 exhibited superior protease activity (5.80 U/ml, p < 0.05) compared to all other strains, while strains BA29 and BA81 showed the lowest protease activity (0.73–0.93 U/ml). Lipase activity was significantly higher (p < 0.05) in strains BA07 and BA16 (0.08 U/ml) compared to the other strains (0.02–0.04 U/ml).
The production of extracellular enzymes, such as amylase and protease, is a key characteristic of B. subtilis strains that contribute to their widespread industrial applications [24]. The ability of B. subtilis to secrete these enzymes is attributed to its sophisticated protein secretion systems (primarily Sec and Tat) and transcriptional regulatory mechanisms (including sigma factors and two-component systems) [25]. These systems coordinately regulate the production and release of enzymes, enabling B. subtilis to adapt to its environment and compete with other microorganisms.
The presence of starch, protein, and fat in poultry feed necessitates the production of amylase, protease, and lipase for efficient digestion. Furthermore, efficient fat digestion is crucial for mitigating the incidence of diarrhea [26]. Therefore, probiotics intended for use in poultry to prevent gastrointestinal diseases should ideally possess the ability to produce all three enzymes.
Cell surface hydrophobicity influences the overall adhesion capacity of bacteria and can facilitate interaction between probiotic bacteria and the host’s epithelial cells. This allows probiotic bacteria to compete with pathogens and produce digestive enzymes and indicates a greater ability of bacteria to adhere to the intestinal mucosa [27]. In this study, strain BA79 exhibited the highest hydrophobicity (66.98%), which was significantly higher (*p *< 0.05) than that of the other strains. Our results are consistent with those of Shahbaz et al. [28], who reported that B. subtilis strains isolated from chicken intestines exhibited hydrophobicity with water contact angles ranging from 62% to 68%.
Antibacterial activity of selected B. subtilis strains. Values with different letters (a-c for E. coli or A, B for S. typhimurium) indicate statistically significant differences (p < 0.05).
The ability of probiotic bacteria to form cellular aggregates through auto-aggregation or co-aggregation enhances their persistence in the gut and can also antagonize pathogenic microorganisms [29]. Strain BA07 exhibited superior co-aggregation ability with both pathogens (38.87%–42.13%) compared to the other strains. Strain BA81 showed the lowest co-aggregation ability (29.86%–30.36%). Strain BA29 displayed the highest co-aggregation ability with S. typhimurium (43.98%) but low co-aggregation with E. coli FG31-1 (31.55%).
Similar to our findings, Ogbuewu et al. [30] reported that B. subtilis and B. amyloliquefaciens isolated from chickens exhibited auto-aggregation abilities of approximately 77% and 72%, respectively, after 2 h. These strains also demonstrated good co-aggregation abilities with other bacteria, reaching approximately 70%. Auto-aggregation in B. subtilis is primarily regulated by cell surface hydrophobicity and surface proteins [20]. Co-aggregation in B. subtilis spp. with other bacteria is modulated by environmental factors and interactions between surface proteins and polysaccharides.
The ability to withstand acidic conditions and bile salts is essential for probiotic bacteria to survive in the challenging environment of the chicken gastrointestinal tract, making tolerance to low pH and bile salts a fundamental requirement when selecting probiotic strains for animal feed [31]. In this study, the tolerance of B. subtilis strains to simulated gastric juice (pH 2 and pH 3) and bile salt conditions (0.3% and 0.5%) mimicking those encountered in the chicken small intestine was evaluated. As proposed by Prabhurajeshwar and Chandrakanth [32], bacteria of host origin often exhibit superior adaptation to the digestive conditions of their host, facilitating more effective colonization compared to bacteria from other sources. Consistent with this notion, all B. subtilis strains in this study demonstrated robust tolerance to both acidic and bile salt conditions, exhibiting high survival rates after 3 h of exposure to acidic environments (83.33%–90.95%) and 4 h of exposure to bile salt environments (82.66%–96.75%). These findings align with those of Penaloza-Vazquez et al. [33], who reported that B. subtilis strains exhibit good tolerance to pH 2.5–3.0 and can survive in the acidic conditions of the chicken stomach. The acid tolerance of B. subtilis can be attributed to its sophisticated intracellular pH regulatory systems, which include proton pumps, ion exchange mechanisms, and the production of intracellular buffers, enabling it to maintain a stable intracellular pH in acidic environments [34]. Furthermore, some B. subtilis strains possess the ability to produce bile salt hydrolase, an enzyme that hydrolyzes bile salts, mitigating their toxicity to bacterial cells [31].
Based on a comprehensive evaluation of probiotic properties, strains BA7 (B. subtilis strain H1) and BA29 (B. subtilis strain BSn5) displayed superior characteristics compared to the other strains (5 vs. 1–4 properties; Table 3) and were therefore selected for further evaluation in the production of experimental biopreparations.
A: PCR products of B. subtilis strains used in this study (M: 100 bp DNA ladder; C: negative control). B: Neighbor-Joining phylogenetic tree of selected B. subtilis strains with an average nucleotide change rate of 0.06 mutations/site.
Viability of probiotic bacteria during storage
Freeze-drying is a widely employed method for preserving probiotic bacteria; however, this process can induce stress that can compromise bacterial viability. To mitigate this, various studies have utilized cryoprotective agents such as skim milk, whey protein, sugars, and biopolymers to enhance the survival rate of probiotics during freeze-drying [35].
Maintaining a sufficient number of viable cells in probiotic preparations is crucial for ensuring their efficacy in animal feed applications. A common starting point of probiotics is approximately 109 CFU/gm; however, this number can significantly decrease to 103–106 CFU/gm during storage [36].
Figure 4 illustrates the viability of the two selected probiotic strains in the fermented preparation after 2 months of storage at 4°C. The initial concentrations of both B. subtilis BSn5 and B. subtilis H1 were approximately 9.4–9.5 log_10_ CFU/ml. Following freeze-drying, the concentration decreased slightly to 9.2 log_10_ CFU/ml for B. subtilis BSn5 and 8.8 log_10_ CFU/ml for B. subtilis H1. During the first week of storage, a significant decrease in the concentration of both strains was observed (*p *< 0.05). However, from the second week onward, the concentration of B. subtilis BSn5 remained stable at around 8.8 log_10_ CFU/ml, while B. subtilis H1 exhibited a gradual decline, reaching approximately 8.0 log_10_ CFU/ml at week 8. The difference in viability between the two strains became statistically significant (*p *< 0.05) from week 5 onward.
These results indicate that B. subtilis BSn5 exhibits superior viability compared to B. subtilis H1 during storage, particularly over the long term, suggesting that B. subtilis BSn5 is a more promising candidate for the production of biopreparations.
Viability of bacteria in the chicken gut
Throughout the 72 h (3-day) monitoring period following oral administration of B. subtilis at a dose of 109 CFU/ml, the chickens exhibited no abnormal clinical signs and maintained normal feed intake. This observation indicates that the B. subtilis strains used in this study are safe for chickens.
*Viability of probiotic strains during storage. *Indicates the number of viable bacteria before and after freeze-drying; **indicates statistically significant differences (p < 0.05) between B. subtilis strains within the same storage week; values with different letters (a–e, A–C) indicate statistically significant differences (p < 0.05) within the same B. subtilis strain.
Persistence of B. subtilis BSn5 in the ileum, cecum, and colon of chickens at different time points after probiotic administration.
Figure 5 depicts the persistence of B. subtilis BSn5 in the ileum, cecum, and colon of chickens at 24, 48, and 72 h post-administration. Overall, the viability of B. subtilis BSn5 in the chicken gut decreased gradually over time. While the strain exhibited relatively good persistence in the first 24 h (4.09–4.20 log_10_ CFU/gm), a significant decline was observed at 48 h (3.36–3.60 log_10_ CFU/gm) and 72 h (2.88–3.11 log_10_ CFU/gm). Furthermore, the concentration of B. subtilis BSn5 was higher in the ileum and cecum compared to the colon (4.87–5.40 log_10_ CFU/gm vs. 4.28–5.09 log_10_ CFU/gm). This observation can be attributed to the rapid transit of digesta through the ileum, coupled with the presence of natural defense mechanisms such as digestive enzymes and bile, which may contribute to the faster decline in bacterial viability in this segment. In contrast, the cecum provides a more stable environment and harbors a diverse indigenous microbiota that may offer support and facilitate the persistence of B. subtilis.
As noted by Chandrasekaran et al. [37], probiotics are often transient inhabitants of the gut, persisting only for a limited time, and are not considered permanent members of the host’s microbiota. Therefore, regular probiotic supplementation at appropriate concentrations is recommended to maintain their beneficial effects.
Conclusion
This study successfully isolated and characterized B. subtilis BSn5 from indigenous chicken feces, demonstrating its potential as an antibiotic alternative in poultry production. This strain not only exhibited potent antibacterial activity against E. coli and Salmonella spp. but also possessed key probiotic properties, including the ability to produce extracellular enzymes, cell surface hydrophobicity, auto-aggregation and co-aggregation capabilities, and tolerance to the harsh conditions of the gastrointestinal tract. Furthermore, this probiotic strain demonstrated good viability during storage. In vivo studies revealed that while B. subtilis BSn5 can survive in the chicken gut, its persistence is transient, necessitating regular supplementation to maintain its beneficial effects. Further in vivo studies in chickens are warranted to confirm its efficacy and develop more stable biopreparations.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Newman DM Barbieri NL de Oliveira AL Willis D Nolan LK Logue CM Characterizing avian pathogenic Escherichia coli (APEC) from Colibacillosis cases, 2018 Peer J 20219 e 11025 https://doi.org/10.7717/peerj.110253371771310.7717/peerj.11025 PMC 7937341 · doi ↗ · pubmed ↗
- 2Altayb HN Badri RM Chaieb K Moglad E Detection and characterization of the most common foodborne pathogens by using multiplex PCR procedure Saudi J Biol Sci 2023306103653 https://doi.org/10.1016/j.sjbs.2023.1036533722364010.1016/j.sjbs.2023.103653 PMC 10200967 · doi ↗ · pubmed ↗
- 3Mehat JW van Vliet AH La Ragione RM The avian pathogenic Escherichia coli (APEC) pathotype is comprised of multiple distinct, independent genotypes Avian Pathol 202150540216 https://doi.org/10.1080/03079457.2021.19159603404764410.1080/03079457.2021.1915960 · doi ↗ · pubmed ↗
- 4World Health Organization WHO bacterial priority pathogens list, 2024: Bacterial pathogens of public health importance to guide research, development and strategies to prevent and control antimicrobial resistance 2024 January 01 2025 Geneva, Switzerland World Health Organizationhttps://www.who.int/publications/i/item/9789240093461
- 5Mudenda S Malama S Munyeme M Matafwali SK Kapila P Katemangwe P Antimicrobial resistance profiles of Escherichia coli isolated from laying hens in Zambia: implications and significance on one health JAC-Antimicrob Resist 202353 dlad 060https://doi.org/10.1093/jacamr/dlad 0603722339210.1093/jacamr/dlad 060PMC 10202439 · doi ↗ · pubmed ↗
- 6Farhat M Khayi S Berrada J Mouahid M Ameur N El-Adawy H Salmonella enterica serovar gallinarum biovars pullorum and gallinarum in poultry: review of pathogenesis, antibiotic resistance, diagnosis and control in the genomic era Antibiotics 202313123 https://doi.org/10.3390/antibiotics 130100233824758210.3390/antibiotics 13010023 PMC 10812584 · doi ↗ · pubmed ↗
- 7Nasreen S Ali S Andleeb S Mechanisms of medicinal, pharmaceutical, and immunomodulatory action of probiotics bacteria and their secondary metabolites against disease management: an overview Folia Microbiol 20246954965 https://doi.org/10.1007/s 12223-024-01155-23853205710.1007/s 12223-024-01155-2 · doi ↗ · pubmed ↗
- 8Kazemi SA Ahmadi H Karimi TMA Evaluating two multistrain probiotics on growth performance, intestinal morphology, lipid oxidation and ileal microflora in chickens Anim Physiol Anim Nutr 201910351399407 https://doi.org/10.1111/jpn.1312410.1111/jpn.1312431141245 · doi ↗ · pubmed ↗