Draft genome sequence of Flavobacterium aquidurense strain, isolated from untreated wastewater
Alexander D. H. Kingdon, Kara D’Arcy, Anya Breen, Claudia McKeown, Ellie Allman, Priyanka Sharma, Amy McLeman, Adam P. Roberts

TL;DR
Scientists sequenced the genome of a Flavobacterium aquidurense strain from wastewater, revealing genes for pigments and antibiotic resistance.
Contribution
The study provides a new draft genome with insights into pigment biosynthesis and antimicrobial resistance in Flavobacterium.
Findings
The genome contains genes for flexirubin and β-carotene production.
A putative novel β-lactone biosynthetic gene cluster was identified.
A β-lactamase gene and multiple copies of an IS3 family insertion sequence were found.
Abstract
Here, we report the draft 5.8 Mb genome sequence of a Flavobacterium aquidurense isolate from untreated wastewater in Liverpool, United Kingdom. The reported isolate has the potential to produce both flexirubin and β-carotene pigments, and contains an additional biosynthetic gene cluster for a putative novel β-lactone. The genome also contains a gene for a putative β-lactamase blaJOHN-1 analogue, and there are multiple copies of a putative novel insertion sequence of the IS3 family. This genome adds to a growing resource of Flavobacterium spp. sequencing data which can be utilized to investigate microbial pigment production, antimicrobial resistance genes and mobile genetic elements within this genus.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
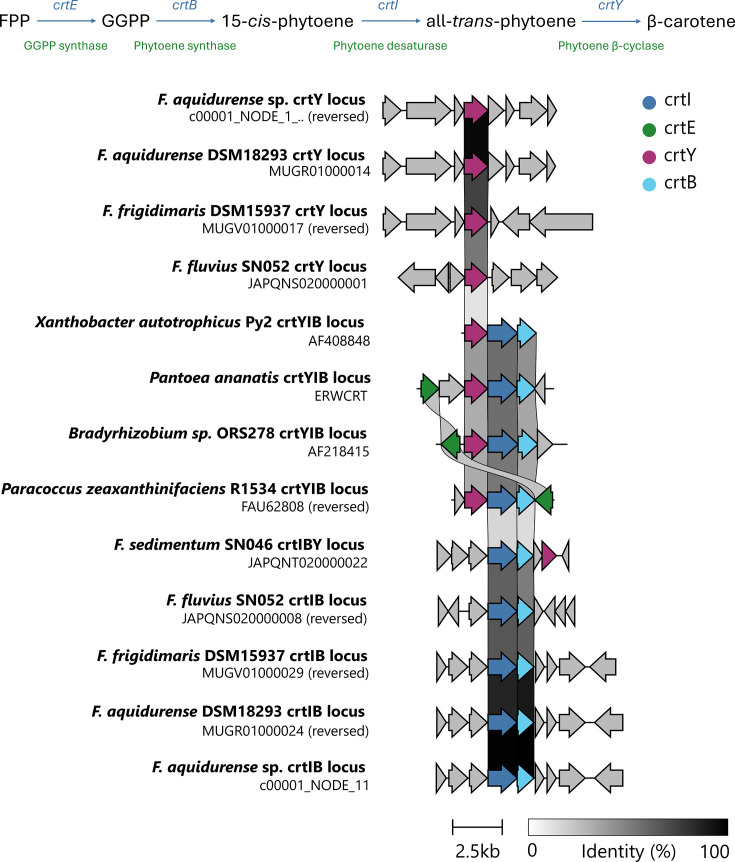 Fig. 1
Fig. 1| Gene name | Gene function | Genome location | Locus tag | Amino acid sequence identity (coverage) | ||||
|---|---|---|---|---|---|---|---|---|
|
| Phytoene synthase | Contig 11:36067–36906 | ACGI8V_20220 | 100% (100%) | 92.1% (100%) | 68.46% (100%) | 57.97% (99%) | 23.9% (92%) |
|
| Phytoene desaturase | Contig 11:34597–36063 | ACGI8V_20215 | 99.2% (100%) | 88.52% (100%) | 76.08% (99%) | 65.42% (98%) | 25.67% (98%) |
|
| Lycopene β-cyclase | Contig 1:107776–108981 | ACGI8V_00530 | 97.8% (100%) | 80.2% (98%) | 59.54% (97%) |
| 19.5% (95%) |
- —http://dx.doi.org/10.13039/100014013 UK Research and Innovation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Metabolism and Applications · Aquaculture disease management and microbiota · Enzyme Production and Characterization
Data Availability
The data are available as part of BioProject PRJNA1161700. The raw reads have been deposited in SRA under the accession no. SRR30678054. The assembled and annotated genome has been deposited in GenBank, under accession no. JBIEAU000000000.
Introduction
Flavobacterium spp. are commonly isolated from soil and water samples [13], and in recent years from Antarctic habitats [46], and are considered opportunistic pathogens in several fish species [79]. There have been limited reports of human infections caused by Flavobacterium spp., specifically F. ceti [1011] and F. lindanitolerans [1213]. We report here the genome sequence of a Flavobacterium spp. isolate from untreated wastewater from Liverpool, United Kingdom.
Methods
Untreated wastewater was collected from the outflow of the Liverpool Life Sciences Accelerator, part of the Liverpool School of Tropical Medicine (53°24′ N, 2°58′ W, United Kingdom) in March 2021 [14]. Sterile swabs were used to spread wastewater onto brain–heart infusion (BHI) agar followed by incubation at room temperature for 2 days. Individual colonies of bacteria were collected and stored as part of the Swab and Send project [15]. The isolate was cultured statically in BHI broth at room temperature for 2 days, before being stored in 20% glycerol at −70 °C. The organism was preliminarily identified using 16S rRNA primers 27F (AGAGTTTGATCCTGGCTCAG) and 1492R (GGTTACCTTGTTACGACTT) in a colony PCR, followed by Sanger sequencing (Azenta). The amplicon sequence was run through BLASTn against the core nucleotide database [16]. The closest match to this partial sequence was Flavobacterium frigidimaris DSM15937, an isolate from Antarctic seawater [5], accession no. NR_041057.1. Due to the large differences in the environment of isolation, we decided to obtain the Flavobacterium aquidurense genome sequence.
A single orange colony was cultured in BHI broth, shaking at 250 r.p.m., at 28 °C, until mid-log was reached. Once at an OD_600_ of 0.9, 10 ml of culture was pelleted, transferred to 500 µl DNA/RNA Shield (Zymo Research, USA) and sent to MicrobesNG (https://microbesng.com/) for processing as follows: An aliquot of this cell suspension (40–50 µl) was lysed using tris ethylenediaminetetraacetic acid (TE) buffer containing lysozyme, metapolyzyme and RNase A (120 µl) incubated at 37 °C for 25 min. To this mixture, proteinase K (final concentration=0.1 mg ml^−1^) and SDS (final concentration=0.5% v/v) were added and incubated at 65 °C for 5 min. DNA was purified using solid-phase reversible immobilization beads, then resuspended in elution buffer (EB, equal volume). Library preparation was performed using the Nextera XT Library Prep Kit (Illumina, USA) following the default protocol, except the input DNA was increased two-fold and the PCR elongation time increased to 45 s. An Illumina NovaSeq 6000 was used for short-read sequencing using a 250 bp paired end protocol. This resulted in 1,091,197 raw reads. For all software processing, the default parameters were used unless stated. The read adapters were trimmed using Trimmomatic v.0.30 with a sliding window quality cut-off of Q15 [17]. Genome assembly was undertaken using SPAdes v.3.7 [18] and annotated using NCBI’s Prokaryotic Genome Annotation Pipeline v.6.8 [19].
Genome description
The reported Flavobacterium sp. isolate genome has a total size of 5,799,299 bp and overall GC content of 34.85%. The mean coverage across the genome was 88.26×. Genome assembly resulted in 60 contigs with an N_50_ of 436,906 bp. The genome assembly had a completeness of 99.22% and contamination of 0.89%, as assessed by CheckM [20]. Across these contigs, there were 4,921 predicted genes, plus 63 tRNA sequences. The draft genome was compared to the closest 16S rRNA match, F. frigidimaris DSM15937 (MUGV00000000), revealing low genome similarity with an average nucleotide identity (ANI) of 82.23 (44.15% coverage) [21]; however, the closest match was to Flavobacterium aquidurense DSM18293 (MUGR00000000), ANI 98.58 (63.37% coverage) [8]. ANI was calculated using EZBioCloud’s webserver (https://www.ezbiocloud.net/tools/ani), which used the OrthoANIu algorithm. Retrospective comparison to the gyrB gene also highlighted F. aquidurense DSM18293 as the closer match, having an amino acid sequence identity of 100% (100% coverage), compared to 96.6% (100% coverage) against F. frigidimaris DSM15937.
As the production of pigments is common for Flavobacterium spp*.* [1], the potentially encoded secondary metabolites were investigated using AntiSMASH v.7.0 [22]. Two main groups of pigments produced by Flavobacterium spp. are flexirubins and carotenoids, with only one group of compounds typically being produced per isolate; however, some isolates do produce both [1]. Analysis of the AntiSMASH output showed potential for flexirubin production, with 32 of the 35 genes within the single biosynthetic gene cluster (BGC) showing greater than 80% nucleotide identity to the flexirubin BGC from Flavobacterium johnsoniae [23]. In addition, the four essential β-ketoacyl synthase genes within this BGC showed between 89% and 94% nucleotide identity (100% coverage). Genes involved in β-carotene production were also present within the sequenced isolate (Table 1), one of each phytoene synthase (crtB), phytoene desaturase (crtI) and lycopene β-cyclase (crtY) genes (Fig. 1) [24]. A pair of crtB and crtI genes was adjacently located on contig 11, with the remaining crtY gene being on contig 1. In many other bacterial species, the majority from the phylum Pseudomonadota and also Flavobacterium sedimentum SUN046, all three genes are co-localized within a single BGC [2527]. However, this is not the case for the F. aquidurense isolate reported here, and several other Flavobacterium spp. (Fig. 1 and Table 1) [5826]. Carotenoid biosynthesis is under-explored in Flavobacterium spp., as the only study previously characterizing Flavobacterium sp. strain R1534 carotenoid biosynthesis; the strain was later re-classified as Paracoccus zeaxanthinifaciens ATCC 21588 [2528]. The genes identified herein were compared for similarity to the crt genes found in * F. aquidurense, F. frigidimaris, F. sedimentum, F. fluvius* and the reclassified P. zeaxanthinifaciens (Table 1). This comparative analysis highlights the high levels of nucleotide similarity within the BGCs of Flavobacterium spp., but low similarity to * P. zeaxanthinifaciens,* with the exception being no significant nucleotide identity to the crtY gene in F. sedimentum, which was reported as a rare crtY_cd_ variant [26]. AntiSMASH analysis also predicted the presence of genes whose encoded proteins could synthesize a novel β-lactone-type compound. β-lactones are strained heterocyclic four-membered rings, which can be used as intermediates during total synthesis, and β-lactone-containing natural products are increasingly being found to have bioactivity as enzyme inhibitors [29]. This prediction is based on the presence of two genes, one encoding an hydroxymethylglutaryl-CoA lyase(HMGL)-like protein and one encoding an AMP-binding protein, within the same BGC [2930]. The putative BGC was also compared to the MIBiG 4.0 database of previously characterized BGCs, but no significant matches were found [2231].
(a) β-Carotene biosynthetic gene pathway from the farnesyl pyrophosphate (FPP) precursor molecule. Gene names are highlighted in blue, while encoded protein functions are highlighted in green. GGPP, geranylgeranyl pyrophosphate (b) Organization of the β-carotene BGC, highlighting the four crt genes that are involved. The identity cut-off was <20%. The gene identities between non-crt genes are not shown. The Flavobacterium spp. loci containing crt genes are split by the presence of crtIB and crtY. All the comparative Pseudomonadota have a single locus that contains the crt genes. Part B was generated using clinker [40].
As some Flavobacterium spp. are considered pathogenic, the resistance gene profile was explored using the Comprehensive Antibiotic Resistance Database (CARD [32]) and ResFinder [33]. We identified the β-lactamase resistance gene bla_JOHN-1_ (77.8% amino acid sequence identity, 81.4% nucleotide sequence identity), originally found in F. johnsoniae [34] and confirmed the presence of the six residues required for zinc coordination in subclass-B1 metallo-β-lactamases (His116, His118, Asp120, His196, Cys221 and His263) (consensus class B1 numbering [35]). The draft genome sequence was also screened for the presence of mobile genetic elements (MGEs) using MobileElementFinder [36]. There were two low identity matches to IS3 family MGEs, ISFnu6 [37], 70.2% nucleotide identity across 35.6% of the sequence, and ISEnfa3 [38], 72.3% nucleotide identity over 26.1% sequence coverage. However, these putative novel insertion sequences were not associated with bla_JOHN-1_ and were present on different contigs.
This work indicates a widespread geographic distribution following this isolation in north-west England in addition to previously being isolated in central Germany [3]. The initial mischaracterization of our strain as F. frigidimaris based on 16S rRNA sequence comparison indicates that using the gyrB sequence for initial Flavobacterium species assignment is a preferable taxonomic indicator, which agrees with previous findings [2]. The discovery of a bla_JOHN-1_ analogue within F. aquidurense indicates that this carbapenemase is more widespread than its initial discovery within F. johnsoniae [34] and suggests that Flavobacterium spp. could generally represent a β-lactamase reservoir. Finally, the pigment and β-lactone biosynthetic gene pathways described above show that F. aquidurense could be a useful source of natural products with multiple industrial applications [2939].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bernardet J-F Nakagawa Y Holmes B Proposed minimal standards for describing new taxa of the family Flavobacteriaceae and emended description of the family Int J Syst Evol Microbiol 2002521049107010.1099/00207713-52-3-104912054224 · doi ↗ · pubmed ↗
- 2Peeters K Willems A The gyr B gene is a useful phylogenetic marker for exploring the diversity of Flavobacterium strains isolated from terrestrial and aquatic habitats in Antarctica FEMS Microbiol Lett 201132113014010.1111/j.1574-6968.2011.02326.x 21645050 · doi ↗ · pubmed ↗
- 3Cousin S Päuker O Stackebrandt E Flavobacterium aquidurense sp. nov. and Flavobacterium hercynium sp. nov., from a hard-water creek Int J Syst Evol Microbiol 20075724324910.1099/ijs.0.64556-017267958 · doi ↗ · pubmed ↗
- 4Mc Cammon SA Bowman JP Taxonomy of Antarctic Flavobacterium species: description of Flavobacterium gillisiae sp. nov., Flavobacterium tegetincola sp. nov., and Flavobacterium xanthum sp. nov., nom. rev. and reclassification of [Flavobacterium] salegens as Salegentibacter salegens gen. nov., comb. nov Int J Syst Evol Microbiol 200050 Pt 31055106310.1099/00207713-50-3-105510843045 · doi ↗ · pubmed ↗
- 5Nogi Y Soda K Oikawa T Flavobacterium frigidimaris sp. nov., isolated from Antarctic seawater Syst Appl Microbiol 20052831031510.1016/j.syapm.2005.01.00115997703 · doi ↗ · pubmed ↗
- 6Humphry DR George A Black GW Cummings SP Flavobacterium frigidarium sp. nov., an aerobic, psychrophilic, xylanolytic and laminarinolytic bacterium from Antarctica Int J Syst Evol Microbiol 2001511235124310.1099/00207713-51-4-123511491319 · doi ↗ · pubmed ↗
- 7Heckman TI Yazdi Z Pomaranski EK Sebastião F de A Mukkatira K et al Atypical flavobacteria recovered from diseased fish in the Western United States Front Cell Infect Microbiol 202313114903210.3389/fcimb.2023.114903237153143 PMC 10161732 · doi ↗ · pubmed ↗
- 8Stine CB Li C Crosby TC Hasbrouck NR Lam C et al Draft whole-genome sequences of 18 Flavobacterium spp Genome Announc 20175 e 00865172914684110.1128/genome A.00865-17PMC 5690318 · doi ↗ · pubmed ↗