Development and characterization of a saturated transposon mutant library of Salmonella enterica serovar Enteritidis LA5
Jérôme Trotereau, Anne-Sophie Huguet, Romain Jouan, Florent Kempf, Delphine Naquin, Catherine Schouler, Philippe Velge, Peter Mergaert, Isabelle Virlogeux-Payant

TL;DR
This paper describes creating a comprehensive genetic library in Salmonella to study essential genes and virulence factors.
Contribution
A saturated transposon mutant library was developed for Salmonella Enteritidis LA5 to identify essential and virulence-related genes.
Findings
A saturated transposon mutant library was successfully created in Salmonella Enteritidis LA5.
The study identified essential genes required for growth in rich medium.
The library serves as a functional genomics tool for studying virulence determinants.
Abstract
We created and characterized a saturated Tn-seq mutant library in Salmonella enterica subsp. enterica ser. Enteritidis strain LA5. The set of essential genes after culture in rich medium was determined. This functional genomics tool will aid in identifying the roles of chromosomal and plasmid virulence determinants.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
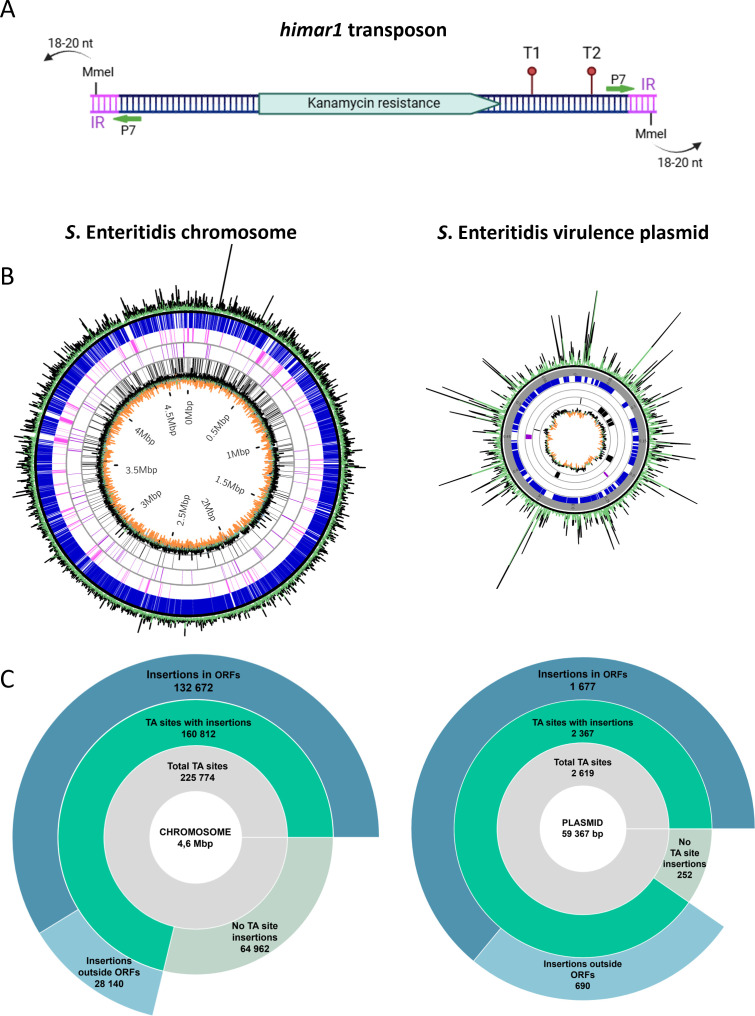 Fig 1
Fig 1| Name | Sequence from 5′ to 3′ | Type | Target (or function) | Reference |
|---|---|---|---|---|
| Bu136 | Adaptor | Bu136 and Bu137 form an adaptor containing the barcode for sample multiplexing | ( | |
| Bu137 | P- | Adaptor |
- —Institut National de Recherche pour l'Agriculture, l'Alimentation et l'Environnementhttp://dx.doi.org/10.13039/501100022077
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Salmonella and Campylobacter epidemiology · Genomics and Phylogenetic Studies
ANNOUNCEMENT
Salmonella enterica subsp. enterica ser. Enteritidis is one of the most commonly isolated Salmonella serovars in humans (1), causing diseases like gastroenteritis in humans, lethal systemic infections in mice, and remaining asymptomatic in poultry (2–4). One promising approach to studying pathogens is the use of transposon-saturated libraries coupled with high-throughput sequencing (Tn-seq). To implement this approach, we constructed a library using the S. Enteritidis LA5 strain, a phage type 4 wild-type field isolate from infected chickens widely characterized in various in vitro and in vivo models by the research community (5–9).
We used a Himar1 mariner transposon (Fig. 1A) (10), which inserts into TA sequences. The LA5 genome contains 225,774 and 2,619 TA sites in its chromosome and plasmid, respectively, enabling the use of the Himar1 transposon to generate a saturated library. Library construction was performed as previously described (11, 12). Briefly, conjugation between E. coli MFDpir (pSAM_EC) and S. Enteritidis LA5 was performed at 37°C on tryptic soy agar (TSA) plates with 300 µg/mL DAP. After 12 hours, transconjugants were enumerated to assess library complexity, and the rest were plated on 400 TSA plates with 50 µg/mL kanamycin, without DAP, to expand the library for 18 hours at 37°C. The library yielded 2 × 10⁶ clones, which were pooled and stored at −80°C in 25% glycerol. Mutant DNA was extracted (Microbial DNA Mini Kit, Macherey-Nagel), processed (restriction, dephosphorylation, adaptor ligation, PCR), and sequenced (75 bp single-read, Illumina NextSeq 550, I2BC, CNRS). A total of 20,111,527 reads (110× coverage) were mapped to TA sites in the S. Enteritidis LA5 775 complete genome, which is a spontaneous streptomycin-resistant mutant of LA5 (K42T mutation in rpsL). Adaptor primers are listed in Table 1. The FastQC (FastQC v0.11.9, available online at: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/) results are as expected, that is, with quality declining after 50 bases due to adapter sequencing. Barcodes were removed using Cutadapt 3.5 (13), transposons eliminated, and sequences filtered to 14–18 bases using SeqKit 2.0.0 (14). Sequences were reverse-complemented, and only those starting with “TA” were selected. The processed sequences were aligned to the LA5 775 complete genome using Bowtie 1.3.1 (15) with no mismatches allowed and only one alignment per read. Finally, a Wiggle file was generated. The cleaned and aligned data were analyzed using TRANSIT v3.3.8 (16), as described previously (11), to identify essential genes and those whose disruption confers a growth advantage or a disadvantage to Salmonella Enteritidis on TSA agar. Quantile normalization was applied to read counts to equalize the distribution of insertion reads across all sites, minimizing biases caused by technical variability or over-represented regions. A total of 160,812 TA sites with insertions were identified on the chromosome, and 2,367 on the plasmid in the S. Enteritidis LA5 Tn-seq library, with an average insertion every 30 base pairs (Fig. 1B). No mutants were detected in four chromosomal ORFs (RUI05_RS05600, RUI05_RS08405, RUI05_RS09135, and RUI05_RS20470) and one plasmid gene (RUI05_RS22785) due to the absence of TA sites and exclusion of the first and last 10% of each gene from TRANSIT analysis.
(A) Representative scheme of the himar1 transposon. Inverted repeat (IR) sequences are shown in purple, with the MmeI recognition site located within them. Arrows indicate that MmeI cleavage sites are outside the transposon. The regions where primers P7 hybridize are marked with green arrows. Transcription terminators T1 and T2, corresponding to rrnB T1 and rrnB T2, are represented as red schematic symbols. (B) Circular representation of the Tn-seq library insertions in the S. Enteritidis genome, generated using the CIRCOS software (17). Both the chromosome and the virulence plasmid are saturated with insertions, depicted as green and black lines, respectively, with the latter being above the average. Through the use of TRANSIT software, we have identified various types of genes: non-essential genes (blue lines), essential genes (pink lines), genes with insertion resulting in a growth defect (purple lines), and/or a growth advantage (black lines). The GC percent of the genome is visualized by black, green, and orange colors on the innermost circle, representing below average, around average, and above average GC content, respectively. (C) The number of TA sites and the distribution of insertions, both within and outside the ORFs, are presented.
On the chromosome, we identified 332 essential genes, as well as 223 and 292 genes whose disruption confers growth disadvantage or advantage on TSA plates, respectively. On the pSEV virulence plasmid, we found 2 and 10 genes with insertion-related growth disadvantage or growth advantage on TSA plates, respectively (Fig. 1C).
This Tn-seq mutant library in Salmonella Enteritidis LA5 will enable researchers to identify genetic determinants important in various conditions, thus allowing the study of pathogenicity in various hosts. The library is available upon request from the corresponding author.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1European Food Safety Authority and European Centre for Disease Prevention and Control. 2022. The european union one health 2021 zoonoses report. EFSA journal. doi:10.2903/j.efsa.2022.7666 PMC 974572736524203 · doi ↗ · pubmed ↗
- 2Bäumler AJ, Tsolis RM, Ficht TA, Adams LG. 1998. Evolution of host adaptation in Salmonella enterica. Infect Immun 66:4579–4587. doi:10.1128/IAI.66.10.4579-4587.19989746553 PMC 108564 · doi ↗ · pubmed ↗
- 3Mancin M, Barco L, Losasso C, Belluco S, Cibin V, Mazzucato M, Bilei S, Carullo MR, Decastelli L, Di Giannatale E, D’Incau M, Goffredo E, Lollai S, Piraino C, Scuota S, Staffolani M, Tagliabue S, Ricci A. 2018. Salmonella serovar distribution from non-human sources in Italy; results from the IT-Enter-Vet network. Vet Rec 183:69–69. doi:10.1136/vr.10490729980593 · doi ↗ · pubmed ↗
- 4Uzzau S, Brown DJ, Wallis T, Rubino S, Leori G, Bernard S, Casadesús J, Platt DJ, Olsen JE. 2000. Host adapted serotypes of Salmonella enterica. Epidemiol Infect 125:229–255. doi:10.1017/s 095026889900437911117946 PMC 2869595 · doi ↗ · pubmed ↗
- 5Allen-Vercoe E, Dibb-Fuller M, Thorns CJ, Woodward MJ. 1997. SEF 17 fimbriae are essential for the convoluted colonial morphology of Salmonella enteritidis. FEMS Microbiol Lett 153:33–42. doi:10.1111/j.1574-6968.1997.tb 10460.x 9252570 · doi ↗ · pubmed ↗
- 6Cazals A, Rau A, Estellé J, Bruneau N, Coville J-L, Menanteau P, Rossignol M-N, Jardet D, Bevilacqua C, Bed’Hom B, Velge P, Calenge F. 2022. Comparative analysis of the caecal tonsil transcriptome in two chicken lines experimentally infected with Salmonella Enteritidis. P Lo S One 17:e 0270012. doi:10.1371/journal.pone.027001235976909 PMC 9384989 · doi ↗ · pubmed ↗
- 7Kempf F, Menanteau P, Rychlik I, Kubasová T, Trotereau J, Virlogeux-Payant I, Schaeffer S, Schouler C, Drumo R, Guitton E, Velge P. 2020. Gut microbiota composition before infection determines the Salmonella super- and low-shedder phenotypes in chicken. Microb Biotechnol 13:1611–1630. doi:10.1111/1751-7915.1362132639676 PMC 7415355 · doi ↗ · pubmed ↗
- 8Naughton PJ, Grant G, Bardocz S, Allen-Vercoe E, Woodward MJ, Pusztai A. 2001. Expression of type 1 fimbriae (SEF 21) of Salmonella enterica serotype enteritidis in the early colonisation of the rat intestine. J Med Microbiol 50:191–197. doi:10.1099/0022-1317-50-2-19111211228 · doi ↗ · pubmed ↗