Complete genome sequence of Helicobacter pylori isolated from residents in southwestern Colombia using Oxford Nanopore sequencing technology
Lizeth Mejia, Julie Benavides-Melo, Ernesto Argoty, Liliana Montenegro, Nelson Rivera-Franco, Diana López-Alvarez, Álvaro Pazos

TL;DR
This paper reports the full genome sequences of Helicobacter pylori from Colombian patients, using new sequencing technology to study its potential link to gastric cancer.
Contribution
The study provides complete genome sequences of H. pylori strains from Colombia and identifies mobilizable plasmids with MOBP-type relaxases.
Findings
Genome sequences of H. pylori from Colombian patients were assembled using Nanopore sequencing.
Some strains contained mobilizable plasmids with MOBP-type relaxases.
Virulence genes in H. pylori may explain its association with gastric cancer.
Abstract
Genome sequences of Helicobacter pylori strains isolated from patients in Nariño, Colombia, with gastric lesions were assembled using Nanopore sequencing. Plasmids were detected in some strains and were predicted to be mobilizable, with relaxases of the MOBP type. H. pylori’s virulence genes may explain the link with gastric cancer.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
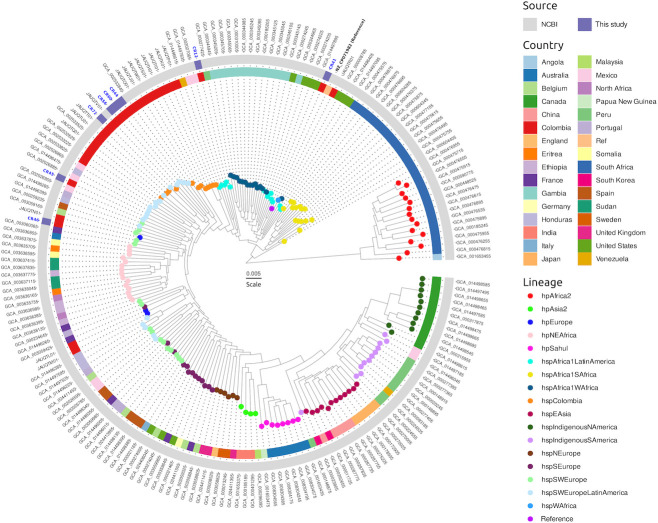 Fig 1
Fig 1| Strain ID | GenBank accessions | Diagnosis | Age | Sex | Host residence | SRA accession | Raw reads | Reads | Genome size (bp) | Mean coverage (×) | No. of contigs | Contig | CDS | rRNAs | tRNAs | ncRNAs | Lineage |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CR12 | NAG | 33 | F | Florida |
| 809,317 | 536 | 1,706,154 | 193.6 | 2 | 1,700,838 | 1,601 | 6 | 36 | 3 | hspAfrica1LatinAmerica | |
| CR41 |
| NAG | 30 | F | Barbacoas |
| 225,971 | 477 | 1,612,862 | 49.9 | 31 | 108,384 | 1,596 | 4 | 36 | 3 | hspAfrica1LatinAmerica |
| CR44 |
| AGIM | 36 | M | Pasto |
| 1,192,402 | 2,111 | 1,681,553 | 787.4 | 1 | 1,681,553 | 1,586 | 6 | 36 | 3 | hspSWEuropeLatinAmerica |
| CR45 |
| NAG | 34 | M | Pasto |
| 1,635,951 | 453 | 1,734,444 | 345.7 | 70 | 44,824 | 1,683 | 6 | 38 | 3 | hspSWEuropeLatinAmerica |
| CR46 |
| NAG | 45 | M | Samaniego |
| 1,842,573 | 840 | 1,623,746 | 653.4 | 1 | 1,623,746 | 1,528 | 6 | 36 | 3 | hspSWEurope |
| CR56 |
| DGC | 24 | M | Pasto |
| 282,596 | 600 | 1,712,951 | 78.8 | 36 | 111,822 | 1,669 | 4 | 37 | 3 | hspSWEuropeLatinAmerica |
| CR60 |
| NAG | 47 | F | Pasto |
| 3,332,538 | 833 | 1,672,799 | 1,129.5 | 1 | 1,672,799 | 1,561 | 6 | 36 | 3 | hspSWEuropeLatinAmerica |
| CR71 |
| NAG | 34 | F | Pasto |
| 803,778 | 403 | 1,683,646 | 151.1 | 31 | 132,000 | 1.652 | 5 | 36 | 3 | hspSWEuropeLatinAmerica |
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —Universidad del Vallehttp://dx.doi.org/10.13039/501100007329
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHelicobacter pylori-related gastroenterology studies · Galectins and Cancer Biology · Genomics and Phylogenetic Studies
ANNOUNCEMENT
Helicobacter pylori, a pathogen colonizing the gastric mucosa of over half the global population (1), induces gastric inflammation, though most infected individuals remain asymptomatic. However, a significant percentage develop gastric or duodenal ulcers (10%–15%) or gastric cancer (1%–3%) (2).
This study analyzed gastric biopsies from the residents of high- and low-risk gastric cancer regions in Nariño, Colombia. Histopathological diagnosis and culturing of antral biopsies were performed according to established protocols (3). Histopathology revealed non-atrophic gastritis (six patients), atrophic gastritis with intestinal metaplasia (one patient), and diffuse gastric cancer (one patient) (Table 1). H. pylori strains were identified by Gram staining and urease, oxidase, and catalase tests. The gastric biopsy sample was cultured on Columbia agar (Oxoid, UK) supplemented with 10% sheep defibrinated blood, selective supplement Dent (Oxoid, UK), and 1% enrichment supplement Isovitalex (Oxoid, UK), under 10% CO_2_ at 37°C for 7–10 days, then cryopreserved in NUNC tubes with 80% sterile thioglycolate and 20% glycerol at −80°C. For DNA extraction, bacterial cultures were grown under the same conditions, but without Dent, using the UltraClean Blood DNA Isolation Kit (MOBIO) and quantified with a Qubit version 3.0 fluorometer (Thermo Fisher Scientific).
MinION sequencing libraries were prepared with ligation sequencing kit (SQK-LSK109) and native barcoding expansion kit (SQK-NBD104) (Oxford Nanopore Technologies, ONT). No DNA shearing or size selection was performed before library preparation. Sequencing was performed on a flow cell (R9.4.1) for 72 h on the MinION (ONT). Default parameters were used for all software unless otherwise specified. Fast5 reads were converted to fastq with Guppy version 6.5.7 in high-accuracy mode (4). Raws fastq were trimmed by cutadapt version 4.9 (5) according to the needs of each sample. The reads were assembled using Flye version 2.9.4 (6) and polished with Pilon version 1.24 (7) (--fix “bases,” “gaps”), Medaka version 2.0.1 (ONT, 2018), and homopolish version 0.4.1 (8). Genome completeness was assessed with BUSCO version 5.5.0 (9) and quality with checkM version 1.2.2 (10). Circularity was determined using Flye version 2.9.4 (6), and coverage depth with Qualimap version 2.2.2 (11). Gene predictions and annotations were performed using PGAP (12). The assembled genomes showed an average GC content of 38%, a size of 1.7 Mbp, and 1,593 protein-coding genes (Table 1).
Four plasmids were detected using MOB-Suite version 3.1.0 (13) in strains CR12 (CP175942), CR41 (JBLDXM010000005 and JBLDXM010000021), and CR71 (JBLDXJ010000026). Plasmids ranged in size from 4,260 to 9,178 bp, with three plasmids classified as mobilizable and containing MOBP-type relaxases (AB508, AE952, and novel plasmid from CR71).
Phylogenetic analysis, based on SNP alignments generated with RealPhy version 1.12, was performed with IQ-TREE version 2.2.6 (14) and visualized with ggtree version 3.12.0 (15) and ggtreeExtra version 1.14.0 (16) packages. A total of 184 H. pylori genomes from NCBI (PRJNA529500) were included, representing previously defined genetic populations and subpopulations (17, 18). The analysis identified four genomes related to hspSWEuropeLatinAmerica, two to hspAfrica1LatinAmerica, one to hspSWEurope, and one to hspColombia (Fig. 1). Raw FASTQ and assembled genomes are available in NCBI under BioProject PRJNA1037030 (Table 1).
Maximum likelihood tree of the full database (192 sequences) of genomes of H. pylori. Color annotations are given in the circles around the terminal nodes, indicating the country of origin for the H. pylori isolates included in the tree. The color of the tips represents the lineage from which each genome was reported. The gray area indicates NCBI genomes, and the blue one the genomes obtained in this study.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ramis IB. 2017. Molecular methods for detection of Helicobacter pylori infection: could they be the gold standard? J Bras Patol Med Lab 53:4. doi:10.5935/1676-2444.20170005 · doi ↗
- 2Thorell K, Bengtsson-Palme J, Liu OH-F, Palacios Gonzales RV, Nookaew I, Rabeneck L, Paszat L, Graham DY, Nielsen J, Lundin SB, Sjöling Å. 2017. In vivo analysis of the viable microbiota and Helicobacter pylori transcriptome in gastric infection and early stages of carcinogenesis. Infect Immun 85:e 00031-17. doi:10.1128/IAI.00031-1728694295 PMC 5607399 · doi ↗ · pubmed ↗
- 3Kodaman N, Pazos A, Schneider BG, Piazuelo MB, Mera R, Sobota RS, Sicinschi LA, Shaffer CL, Romero-Gallo J, de Sablet T, Harder RH, Bravo LE, Peek RM Jr, Wilson KT, Cover TL, Williams SM, Correa P. 2014. Human and Helicobacter pylori coevolution shapes the risk of gastric disease. Proc Natl Acad Sci USA 111:1455–1460. doi:10.1073/pnas.131809311124474772 PMC 3910595 · doi ↗ · pubmed ↗
- 4Wick RR, Judd LM, Holt KE. 2019. Performance of neural network basecalling tools for Oxford Nanopore sequencing. Genome Biol 20:129. doi:10.1186/s 13059-019-1727-y 31234903 PMC 6591954 · doi ↗ · pubmed ↗
- 5Martin M. 2011. Cutadapt removes adapter sequences from high-throughput sequencing reads. EM Bnet J 17:10. doi:10.14806/ej.17.1.200 · doi ↗
- 6Kolmogorov M, Yuan J, Lin Y, Pevzner PA. 2019. Assembly of long, error-prone reads using repeat graphs. Nat Biotechnol 37:540–546. doi:10.1038/s 41587-019-0072-830936562 · doi ↗ · pubmed ↗
- 7Walker BJ, Abeel T, Shea T, Priest M, Abouelliel A, Sakthikumar S, Cuomo CA, Zeng Q, Wortman J, Young SK, Earl AM. 2014. Pilon: an integrated tool for comprehensive microbial variant detection and genome assembly improvement. P Lo S One 9:e 112963. doi:10.1371/journal.pone.011296325409509 PMC 4237348 · doi ↗ · pubmed ↗
- 8Huang Y-T, Liu P-Y, Shih P-W. 2021. Homopolish: a method for the removal of systematic errors in nanopore sequencing by homologous polishing. Genome Biol 22:95. doi:10.1186/s 13059-021-02282-633789731 PMC 8011154 · doi ↗ · pubmed ↗