Exploring the structural landscape of universal stress proteins in Archaea
L. Matarredona, M.J. García-Bonete, B. Zafrilla, J. Esclapez

TL;DR
This paper reviews the structure and function of universal stress proteins in Archaea, focusing on how they help these organisms survive harsh environments and their potential for biotechnology.
Contribution
The paper provides a comprehensive structural and functional analysis of USPs in Archaea, emphasizing their biotechnological potential.
Findings
USPs in Archaea are crucial for surviving environmental stresses like oxidative stress and heavy metal toxicity.
Structural studies of USPs from halophilic and thermophilic Archaea reveal insights into their functional mechanisms.
USPs in Archaea have potential applications in protein engineering and industrial processes.
Abstract
The universal stress protein (USP) superfamily is widely distributed in the three domains of life. These proteins play a key role in the cell response to biotic and abiotic stresses. Although the discovery of the first USP was more than 30 years ago, little is known about its specific function and the molecular mechanisms underlying it. Focusing on the Archaea domain, USPs represent one of the mechanisms developed to survive under environmental stresses, such as nutritional deprivation, oxidative stress, or heavy metal toxicity. This review provides a comprehensive understanding of USPs in this domain, with a focus on their structure and function. It delves into the distribution of USPs in several families of Archaea and explores the structures of selected USPs, paying special attention to those of halophilic and thermophilic microorganisms. Additionally, the biotechnological…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
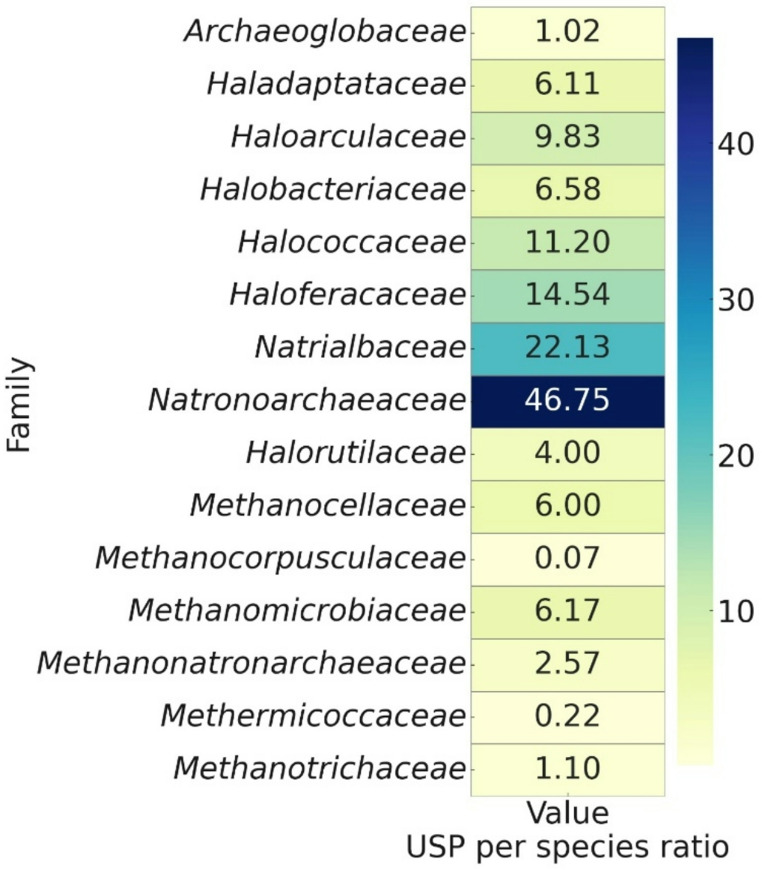 Figure 1
Figure 1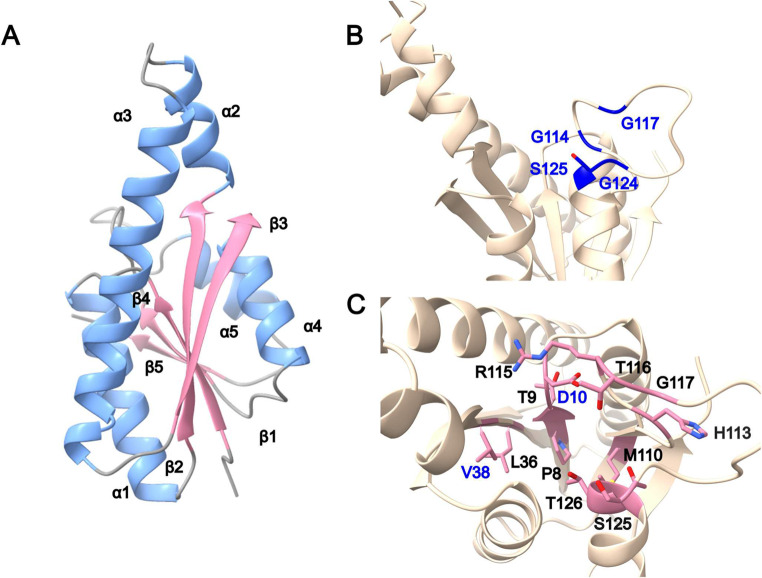 Figure 2
Figure 2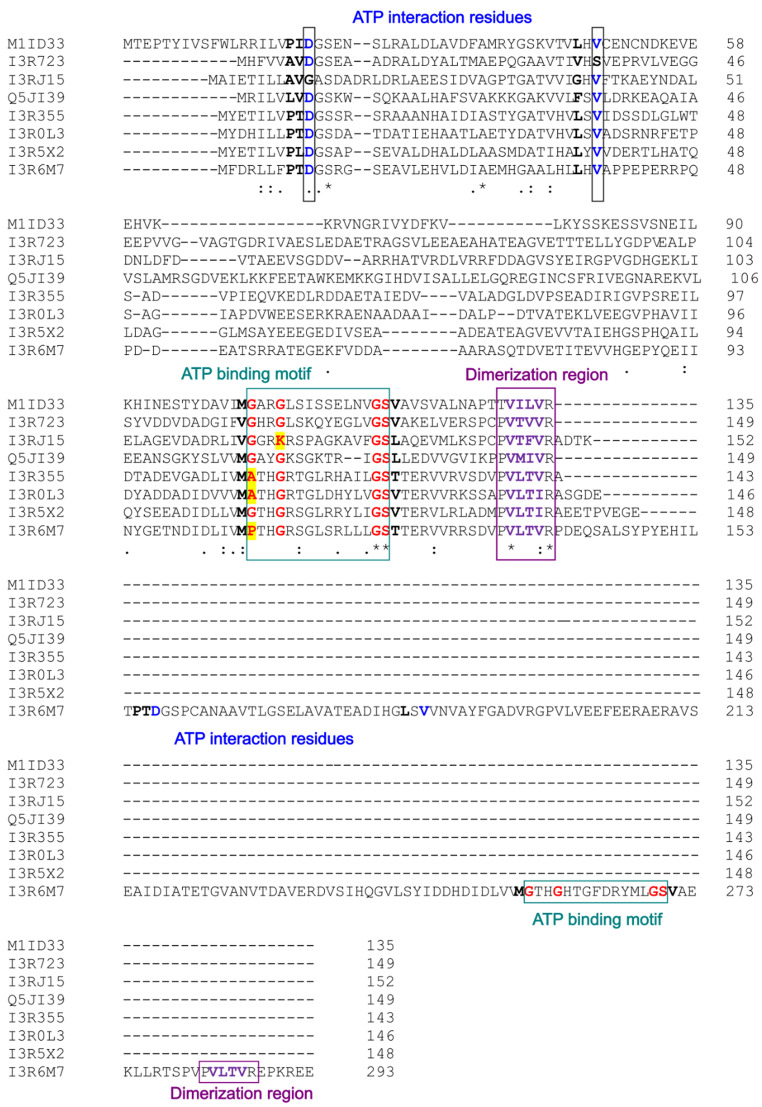 Figure 3
Figure 3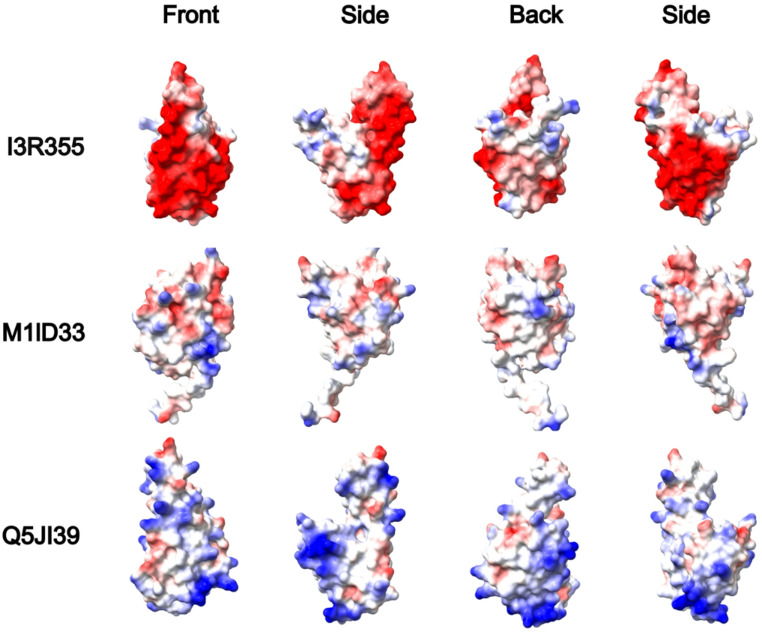 Figure 4
Figure 4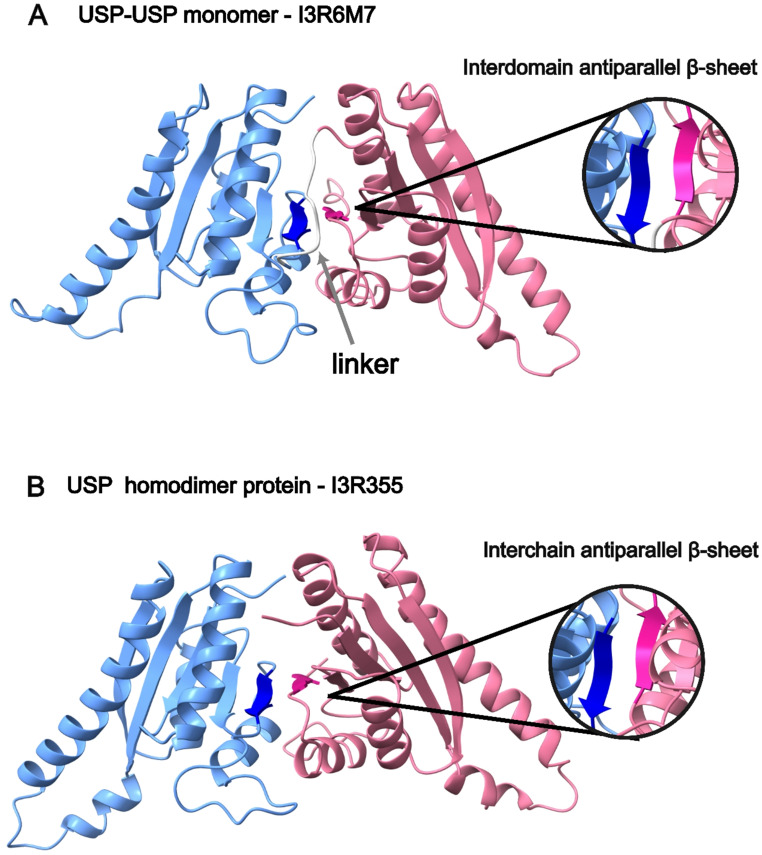 Figure 5
Figure 5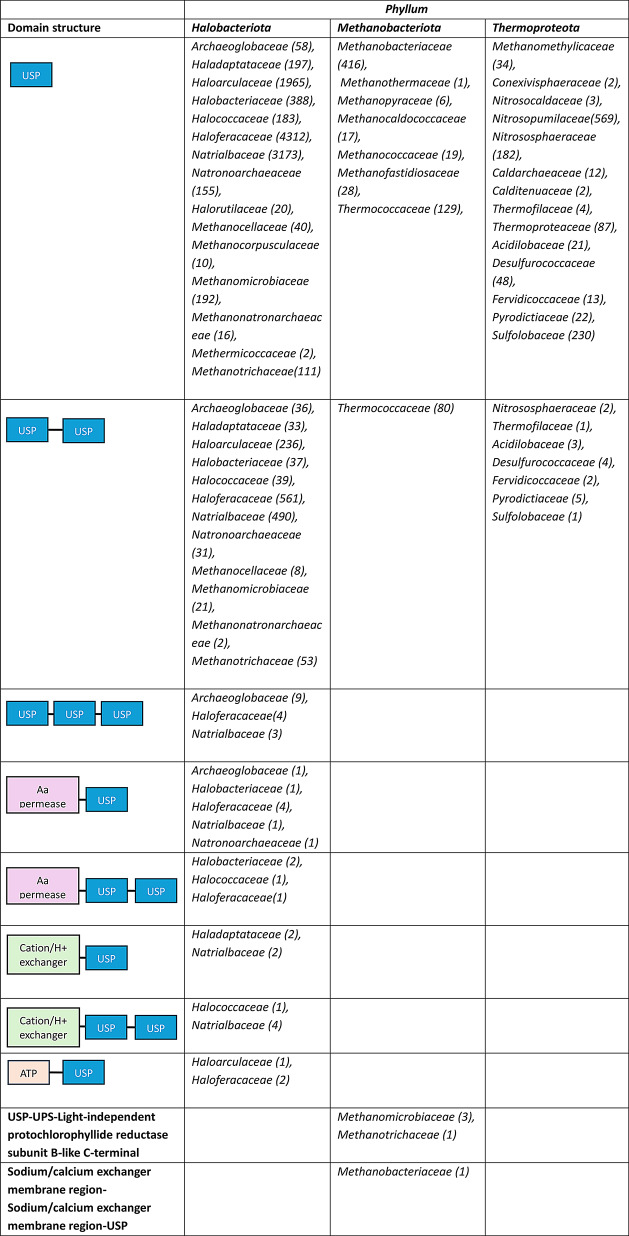 Figure 6
Figure 6- —Universidad de Alicante
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEnzyme Structure and Function · Protein Structure and Dynamics · Bacterial Genetics and Biotechnology
Introduction
Universal stress proteins (USPs), initially discovered in Escherichia coli (Nyström and Neidhardt 1992), are widely distributed in organisms from all three domains of life, including plants, fungi, bacteria and archaea, but they are absent in humans (Siegele 2005; Hingley-Wilson et al. 2010). Generally, these proteins help organisms to cope with unfavorable conditions or biotic and abiotic stresses, playing a role in protein scaffolding, biomolecules transport, preventing protein or DNA denaturation, and electron transport (Gustavsson et al. 2002; Kvint et al. 2003).
The best-known USPs are those for E. coli. Specifically, a family of six homologous USPs (UspA, UspC, UspD, UspE, UspF, and UspG) has been described in this microorganism. Based on sequence homology and structural characteristics, E. coli USPs are classified into four different classes: Class I (UspA, UspC, UspD) lacks an ATP binding motif, whereas Class II, which includes UspF and UspG, contains ATP binding motifs. UspE possesses two tandem USP domains E1 and E2, which are categorized as Class III and IV, respectively (Kvint et al. 2003; Vollmer and Bark 2018). It has been demonstrated that each USP class contributes to different stress tolerance: Class I USPs are involved in processes of oxidative stress resistance, nutrient starvation (carbon, nitrogen, phosphate and sulphur), heavy metals tolerance and iron scavenging; Class II USPs participate in cellular adhesion, cell motility and swimming processes and also in oxidative stress protection; Class III involves signalling molecules that detect stress and activate adaptive responses; Class IV ensures cellular survival and adaptation through synergistic interactions among different USPs under stress (Nabi et al. 2024).
USPs in other bacteria exhibit structural and functional diversity, often through fusion with catalytic motifs such as protein kinases, ion exchangers, and permeases (Luo et al. 2023). These adaptations not only enhance stress tolerance but also increase pathogenicity. For example, in Acinetobacter baumannii, one of the greatest threats to global public health, UspA protects against oxidative stress, low pH and respiratory toxin (Elhosseiny et al. 2014) and in Salmonella typhimurium UspA makes an important contribution to its in vivo virulence (Liu et al. 2007). Similarly, Listeria monocytogenes USPs confer resistance to oxidative and acid conditions (Seifart Gomes et al. 2011), and Mycobacterium tuberculosis Rv2623 is involved in bacterial persistence and chronic infection (Drumm et al. 2009; Glass et al. 2017).
Like the bacterial USPs, plant USPs have multiple functions related to environmental stress protection. The model plant Arabidopsis thaliana possesses more than 40 USPs, which act as molecular chaperone, under heat and oxidative stress, RNA chaperone under cold stress or regulator of ROS production under anoxia (Chi et al. 2019). In Solanum lycopersicum the USP known as SIRd2 is a phosphorylation target of the calcineurin B-like interacting protein kinases (CIPKs). It confers resistance to osmotic and salt stress (Gutiérrez-Beltrán et al. 2017). In addition, it has also been described that plant USPs regulate the photosynthetic efficiency under drought conditions through redox regulation (Loukehaich et al. 2012).
Despite significant research in bacteria and plants, the physiological roles of USPs in Archaea remain largely unexplored, even though two structures have been described in hyperthermophilic microorganisms. The structure of Methanococcus jannaschii MJ0577 was the first USP structure solved in Archaea (Zarembinski et al. 1998) Its ATP binding site was coordinated by a divalent cation and composed of the following residues: G_127_-SH-G_130_-(9X)- G_140_. MJ0577 alone showed negligible ATPase activity, but ATP hydrolysis was detected in the presence of M. jannaschii cell extract suggesting a regulatory role as an ATP-dependent molecular switch. Archaeoglobus fulgidus AF0826 USP was described as a dimeric protein whose structure contains a nearly canonical ATP binding motif that accommodates of the adenosine part of ATP (Tkaczuk et al. 2013). Although these studies described USP structures in Archaea, their function remains still unknown. In recent years, studies with Sulfolobus acidocaldarius USPs have elucidated some of the roles of USPs (Ye et al. 2020a, b). This aerobic thermoacidophile archaeon has three different USP encoded in its genome. SaUspA forms a stable homodimer, conserves the ATP binding motif, and interacts with the serine/threonine phosphatase PP2A, a eukaryotic PP2A catalytic subunit homolog. This interaction was enhanced by ATP binding and stimulates PP2A activity, suggesting a regulatory role in cellular signalling. Deletion mutants of SaUspsA revealed that this USP was involved in nitrogen starvation and salinity stress response acting as a stress modulator via PP2A interaction (Ye et al. 2020a). Focusing on halophilic Archaea, bioinformatic analysis of USPs in six halophilic archaeal families (Halococcaceae,* Halorubraceae*,* Natrialbaceae*,* Haloferacaceae*,* Haloarculaceae*,* and Halobacteriaceae*) revealed that most halophilic USPs contain a single USP domain (140–160 amino acids). However, the presence of the USP domain fused with other catalytic motifs, such as amino acid permease or a Na^+^/H ^+^ exchanger, is also frequent. These domain combinations suggest functional diversification, aiding in stress resistance. Strikingly, several genera, including Halorubrum,* Haloferax*, and Haloarcula, have a relatively high number of USPs (1939, 1301, and 970, respectively), reinforcing their essential response to osmotic stress and cellular homeostasis (Matarredona et al. 2020). In addition, recent studies on Haloferax mediterranei have highlighted the complexity of USP expression under stress, suggesting a coordinated response involving multiple USPs to modulate growth and improve survival under adverse conditions (Matarredona et al. 2024). Transcriptional expression of the 37 annotated USPs in the Hfx. mediterranei genome was examined under three different stress conditions. A clear relationship between the number of USPs and the growth phase was observed, but it was difficult to associate the expression of a single USP with a particular type of stress. Besides, approximately one-third of all USPs remained expressed regardless of the stressor. These results suggested that the USP system acts as a recruitment mechanism, where the number of stress proteins expressed increases depending on the strength of the stressful stimulus. Three Hfx. mediterranei USPs were studied in detail (USP5, USP21, and USP28), showing that all three form stable homodimers. The physiological characterization of the overexpression strains (USP5-HM26, USP21-HM26, and USP28-HM26) under various stress conditions revealed significant changes in growth kinetic parameters, particularly in the presence of nitrate as nitrogen source, as well as in low and high salinities, and under oxidative stress (Matarredona et al. 2024).
Although further research is needed to enhance our understanding of USP functions and their molecular mechanisms, available data highlight the biotechnological potential of these proteins across various fields. Understanding the structure, distribution, and function of USPs in Archaea could provide valuable insights into their adaptation strategies and open up new avenues for industrial and medical applications. This review will focus on the occurrence of USPs in Archaea, examining their structural features, potential functions, and the promise they hold for future biotechnological innovations.
Distribution of USPs in Archaea
Universal Stress Proteins (USPs) in Archaea play a significant role in adapting to extreme environmental conditions, offering insights into these organisms’ survival mechanisms. This section examined the distribution of USPs across the key archaeal phyla: Halobacteriota,* Methanobacteriota*, and Thermoproteota, as classified in the List of Prokaryotic names with Standing in Nomenclature (LPSN) database. These phyla encompass families that thrive in various extreme environments—such as hypersaline (halophiles), anaerobic (methanogens), and high-temperature (thermophiles) habitats. For this study, an analysis of the USPs found in selected archaeal families was performed using the UniProt database. The number of annotated USPs per family was obtained, and these values were normalised by the number of species classified in each family to create a USP per-species ratio. This approach allows comparisons to be made about the number of USPs in different families that are independent of species richness.
The differences in the number of USPs across these phyla provide valuable insights into the stress-response mechanisms that have evolved in Archaea to deal with various environmental pressures. USPs are known to play a crucial role in protecting cells from damage caused by oxidative stress, heavy metal toxicity, nutrient starvation, osmotic stress, and heat shock (Vollmer and Bark 2018; Ye et al. 2020a; Matarredona et al. 2024). However, in contrast to the extensive research on USPs in bacteria and plants (Nabi et al. 2024), studies on archaeal USPs are limited to a few well-studied species, specifically M. jannaschii, S. acidocaldarius, A. fulgidus, and H. mediterranei. The distribution of USPs could reflect the diversity of environmental challenges these organisms face, highlighting their ability to adapt and thrive in such hostile conditions. To make fair comparisons across families with different numbers of species, USP per-species ratio were calculated (Table 1). This USP per species ratio enables us to understand better how much each family invests in stress-response mechanisms, independent of species richness.
Table 1. Number of annotated USPs across Halobacteriora, Methanobacteriota and Thermoproteota archaeal phylaArchaeal phyllumFamilyUSP/Number species Halobacteriota
Archaeoglobaceae 104/102 Haladaptataceae 232/38 Haloarculaceae 2202/224 Halobacteriaceae 428/65 Halococcaceae 224/20 Haloferacaceae 4884/336 Natrialbaceae
Natronoarchaeaceae
Halorutilaceae
Methanocellaceae
Methanocorpusculaceae
Methanomicrobiaceae
Methanonatronarchaeaceae
Methermicoccaceae
Methanotrichaceae 3673/166187/420/548/810/152216/3518/72/9165/150 Methanobacteriota
Methanobacteriace A
Methanothermace B
Methanopyraceae 417/7512/16/9 Methanobacteriota a
Methanocaldococcaceae 17/18 Methanococcaceae 19/53 Methanobacteriota B
Methanofastidiosaceae
Thermococcaceae 28/19209/161 Thermoproteota
Methanomethylicaceae
Conexivisphaeraceae
Nitrosocaldaceae
Nitrosopumilaceae
Nitrososphaeraceae
Caldarchaeaceae
Calditenuaceae
Thermofilaceae
Thermoproteaceae
Acidilobaceae
Desulfurococcaceae
Fervidicoccaceae
Pyrodictiaceae
Sulfolobaceae 34/352/93/17569/617184/16712/131/175/3287/4024/9752/6715/1327/26231/190
In the Halobacteriota phylum, significative variability was observed in USP counts across families, suggesting differential adaptation strategies. Haloferacaceae exhibits the highest USP count (4,884 USPs across 336 species), suggesting a complex and adaptable stress-response system, likely due to fluctuating salinity in its habitat. This family includes H. mediterranei, whose 37 USPs have been studied under different environmental conditions, revealing diverse responses to osmotic stress and nutrient deprivation (Matarredona et al. 2024). Other families, such as Haloarculaceae and Natrialbaceae, also display high USP counts (2,202 and 3,673, respectively), which reflects the high diversity within these families and their adaptation to variable salinity.
In contrast, Archaeoglobaceae, with 104 USPs across 102 species, demonstrates one of the lowest USP per-species ratios (1.02), possibly reflecting adaptation to more stable conditions where extensive stress-response proteins are unnecessary, or suggesting that alternative mechanisms may exist to cope with detrimental conditions. The Natronoarchaeaceae family, with 187 USPs distributed across four species, shows the highest USP per-species ratio (46.75), suggesting a significant reliance on USPs. This reliance might be due to the extreme variability in environmental conditions. Given that members of this family are likely exposed to fluctuating conditions, such as high salinity and alkalinity, the high number of USPs per species may support a broad range of cellular responses, enhancing their resilience to environmental stressors. The elevated USP count per species in Natronoarchaeaceae may support a broad range of cellular responses, enhancing resilience to environmental stressors and suggesting a sophisticated stress management system that remains unknown.
The *Methanobacteriota *phylum, which includes methanogenic archaea adapted to anaerobic environments, displays a wide range of USP counts and ratios across its families. This variability likely reflects the diverse ecological niches and adaptive strategies employed by methanogens to thrive in environments with limited oxygen and variable nutrient availability (Wen et al. 2017). For instance, Methanobacteriaceae, with 417 USPs across 751 species, has a low USP per-species ratio (0.55), suggesting that these microorganisms may inhabit relatively stable anaerobic environments where broad stress-response mechanisms are less critical, or they have different mechanisms to cope with stress conditions. Methanocaldococcaceae and Methanococcaceae exhibit very low USP counts relative to species diversity, indicating limited reliance on these proteins. Methanocaldococcaceae has 17 USPs for 18 species (ratio of 0.94), while Methanococcaceae, with only 19 USPs across 53 species, has one of the lowest ratios (0.36) in this group. Such low values could imply minimal reliance on USPs, as these organisms may have developed specialized adaptations to consistent anaerobic conditions. In Methanobacteriota B, Methanofastidiosaceae has 28 USPs across 19 (ratio of 1.47), while Thermococcaceae has 209 USPs across 161 species (ratio of 1.3).
The *Thermoproteota *phylum, primarily composed of thermophilic archaea, exhibits considerable variability in USP counts and ratios, reflecting distinct adaptive strategies that extend beyond thermal tolerance. This diversity reflects the different ecological niches and adaptive strategies of these organisms, especially in managing stressors beyond temperature, such as pH fluctuations and nutrient limitations. Higher USP counts are observed in families such as Thermoproteaceae (87 USPs across 40 species, with a ratio of 2.17), suggesting a better adaptation to changing environmental factors. Similarly, Sulfolobaceae, which includes well-studied extremophiles like S. acidocaldarius, has 231 USPs across 190 species, yielding a ratio of 1.21. In contrast, other families within Thermoproteota, such as Nitrosocaldaceae (3 USPs across 17 species, ratio of 0.18) and Calditenuaceae (1 USP across 17 species, ratio of 0.06), possess minimal USP counts. This minimal investment implies that these organisms inhabit highly stable thermal environments, where a broad array of USPs may be unnecessary. Their reduced reliance on USPs could indicate a specialization for consistent, high-temperature conditions, with other proteins, such as heat shock proteins (HSPs), fulfilling critical roles in maintaining cellular stability.
The heatmap emphasizes the density of USPs per species across archaeal families (Fig. 1), illustrating distinct adaptive strategies. High ratios in families such as Methanocorpusculaceae and Haloferacaceae indicate a strong reliance on USPs, potentially due to the fluctuating and extreme environmental conditions these organisms face. Conversely, low ratios in families such as Methanocaldococcaceae and Methanobacteriaceae suggest a reliance on specialized proteins or stable habitat conditions, thereby reducing the need for extensive USP-based stress responses.
Fig. 1. Heatmap of the ratio of USPs per-species in Archaea families
Archaeal USP domain structure analysis
Archaeal USPs exhibit considerable structural diversity, with various domain combinations that may represent specialized adaptations to extreme environments, such as high salinity, elevated temperatures, and anaerobic conditions. Analysis of USPs across three major archaeal phyla—Halobacteriota, Methanobacteriota, and Thermoproteota—reveals distinct patterns of domain structures and distribution, suggesting that these proteins play crucial roles in enhancing stress resilience in specific ecological niches.
The structural domains of USPs typically consist of conserved regions made up of 140–160 amino acids and are present either as single-domain proteins or fused with additional structural domains. Common architectures in archaeal USPs include single USP domains, tandem repeats, and combinations with other functional motifs, such as amino acid permeases and cation/H^+^ exchangers. These configurations suggest functional differentiation, potentially enhancing archaeal resilience by integrating stress-sensing and regulatory roles with membrane transport or ATP-binding activities. This structural variety within archaeal USPs may thus play a critical role in their capacity to endure and respond to environmental stresses.
The Halobacteriota phylum exhibits the highest structural diversity, with families like Haloferacaceae, Natrialbaceae, and Haloarculaceae dominating the landscape (Table 2). These families, particularly Haloferacaceae, display a wide diversity of USP domain structures, including singular USP domain, tandem USP-USP structures, and more complex configurations involving other catalytic domains such as the amino acid permease (AA_permease) and Na^+^/H^+^ exchanger.
This structural diversity likely equips Halobacteriota with the flexibility required to adapt to the highly variable osmotic pressures and ionic concentrations characteristic of hypersaline environments. Moreover, the elevated USP counts observed in families like Natronoarchaeaceae and Natrialbaceae suggest a significant evolutionary investment in these proteins, possibly as an adaptive strategy to withstand extreme salinity fluctuations and periodic desiccation.
The *Methanobacteriota *phylum, which comprises methanogenic archaea adapted to anaerobic environments, exhibits relatively low diversity and complexity in USP domain configurations (Table 2). Simple structural types, including single USP domains and basic tandem configurations, primarily characterize families such as Methanobacteriaceae and Methanomicrobiaceae. This structural simplicity suggests that Methanobacteriota may have reduced reliance on broad-spectrum stress-response proteins, likely due to the more stable and consistent conditions found in anoxic environments where they predominantly reside.
Notably, some Methanobacteriota families, such as Methanomicrobiaceae, contain unique domain combinations like the light-independent protochlorophyllide reductase subunit B-like C-terminal domain (LI-POR_suB-like_C) linked with a USP domain. These automatic annotation results are unusual, as the LI-POR_suB-like_C domain has been described in enzymes from cyanobacteria, such as Synechocystis or red algae (Dong et al. 2020). In other non-photosynthetic organisms, this domain is associated with oxidation-reduction reactions (Chernomor et al. 2021). Consequently, this specialized configuration in Methanomicrobiaceae may support metabolic pathways involved in methanogenesis, highlighting a functional adaptation to their anaerobic lifestyle. Additionally, Methanobacteriaceae includes a domain arrangement with a sodium/calcium exchanger, potentially providing ion homeostasis in environments with variable mineral compositions. These unique domain architectures could demonstrate how evolutionary pressures have refined USP structures in the phylum Methanobacteriota, enabling these organisms to adapt to environmental challenges by developing specific structural adaptations rather than possessing a wide variety of USP types.
The *Thermoproteota *phylum, which comprises thermophilic archaea adapted to extremely high temperatures, exhibits a unique pattern in its USP structures. Remarkably, all families within this group contain USPs characterized either by a single USP domain or by simple tandem arrangements of two USP domains (USP-USP) (Table 2). Unlike the other two phyla, no catalytic domains have been identified in the USP of microorganisms belonging to the *Thermoproteota *phylum. This limited structural diversity could suggest that these organisms have evolved in a specialized way to enable them to endure extreme thermal environments.
This is the first work that analyses the diversity of USP domain structures across Archaea. These insights could help guide future research into how USPs support archaeal survival and resilience, with potential applications in biotechnology.
Table 2. Distribution of domain structures in USPs across Halobacteriota,* Methanobacteriota*, and Thermoproteota families
USPs oligomerization state
Generally, USPs in bacteria, archaea and plants exhibit a dimeric quaternary structure, which appears to support their stability and functionality under stress conditions. In Archaea, the homodimeric structure has been observed in M. jannaschii MJ0577 and A. fulgidus AF0826, where studies using techniques like size-exclusion chromatography and dynamic light scattering have consistently demonstrated this dimeric arrangement (Zarembinski et al. 1998; Tkaczuk et al. 2013). In addition, both enzymes contain the ATP binding motif although ATP hydrolysis has not been confirmed in vitro suggesting the requirement for one or more soluble components to stimulate ATPase activity. USP AF1760 from A. fulgidus is a USP-tandem protein which form another dimer of tandem “dimers”, the resulting protein contains four USP domains in total. This oligomerization state has also described in other tandem fusion USPs, such as Thermus thermophilus HB8 TTHA0350 (Iino et al. 2011). The X-ray structure of the TTHA0350 protein showed that the N-terminal and C-terminal domains bind one ATP, although the ATP-binding motif could not be detected on sequence alignment (Iino et al. 2011).
Dimeric structures have also been described in USP5, USP21, and USP28 from H. mediterranei, which form homodimers with molecular weights of 46, 32, and 36 kDa, respectively (Matarredona et al. 2024). Sequence analysis reveals that USP5 and USP21 contain a putative ATP binding motif but not USP28. The USP TeaD from the halophilic bacterium Halomonas elongata showed different oligomerization state depending on the concentration of ATP. In the absence of ATP, TeaD is a homodimer of 32 kDa. Still, incubating the protein with ATP resulted in a higher oligomeric state, corresponding to a tetramer with a molecular weight of 62 kDa. This ATP-dependent oligomerization might have a functional role in the regulatory mechanism (Schweikhard et al. 2010).
Apart from dimeric and tetrameric structures, USPs also exist in other native structures, including monomers, trimers and oligomeric complexes formed by combinations of USP proteins (Jung et al. 2015). The oligomeric state switches in response to various parameters, such as external stresses or intracellular ATP concentrations, and appears to underlie some regulatory mechanisms.
Overall, the oligomerization state of USPs across the three domains of life is likely a key factor in modulating their biochemical and physiological functions, including ATP-binding affinity, protein-protein interactions, and activity regulation, which may determine their role in adapting to cellular stress.
USP structure prediction in Archaea
Single-USP proteins consist of a globular structure with five parallel β-sheets (β1- β2- β3- β4- β5) and 4 or 5 α-helix (α1-“α2”-α3-α4-α5) depending on the specie (Fig. 2A). This extra helix (α2, Fig. 2A) is observed in particular species of Halobacteriota and Methanobacteriota in addition to a longer α3 helix than in Thermoproteota species. This difference is due to a shorter sequence between the β2 sheet and the α3 helix in Thermoproteota (Figure S1). The USP domain proteins also presents a representative motif, G-2X-G-9X-G-(S/T) (Tkaczuk et al. 2013), that allows proper arrangement of the tertiary structure to allocate an ATP binding pocket and ion binding site (Fig. 2B). In some USP proteins, this motif can degenerate presenting a larger or shorter number of non-specific residues between the specific glycine residues (G-2X-G-5/12X-G-(S/T) (Tkaczuk et al. 2013), or in some case a substitution of the first glycine (which is at the end of the β4-strand) by alanine or proline (I3R355, I3R6M7 Fig. 3). Alphafold models (e.g., I3R355, I3R6M7) (Mirdita et al. 2022; Varadi et al. 2024) suggest that several of these USP proteins with degenerative motifs still maintain the tertiary structure of the ATP binding pocket, suggesting the possibility of binding ATP and an ion (Fig. 2B). Other USP proteins (e.g., I3R1J5, IR3R723) (Mirdita et al. 2022; Varadi et al. 2024) do not contain this complete motif, suggesting its inability to bind with ATP. Although this motif is essential in forming the ATP binding pocket, other residues outside this motif are involved in binding the ATP and ions. Considering the crystallographic structure of a single USP protein from M. jannaschii (PDB-ID 1mjh) (Madeira et al. 2024) where ATP is bound, several residues have been identified in the interaction with the ATP molecule. Residue D12 and V30 (reference molecule I3R355) are essential in this interaction and probably in its hydrolyzation (Fig. 2C). Sequence alignment performed with ClustalW Omega shows these differences in ATP binding motif and essential residues (Fig. 3).
Fig. 2I3R355 single USP protein structure model generated with Alphafold. (A) Secondary structure representation. In pink the 5 β-sheet core of the USP domain and in blue the 5 α-helix, including the α_2_ which it is not always present in all the USP. (B) ATP binding domain representation; A-2X-G-9X-G-(S/T), for I3R355. (C) ATP binding pocket including the amino acids involved in the interaction through their side chains or peptide bond
Fig. 3. Sequence alignment of single USP and USP-USP (I3R6M7) proteins from Archaea using ClustalW omega. In red are represented the residues involved in the ATP binding motif (green square). In black are represented residues involved in the interaction with the ATP according to the PDB-ID 1mjh (Madeira et al. 2024) and in blue are the most critical residues. In purple are represented the residues involved in the dimerization of USP domains and the formation of the antiparallel β-sheet
Depending on the microorganism host, the USP proteins can differ in size and surface charge. In Figure S1, we compared three single USP protein from Halobacteriota,* Methanobacteriota* and Thermoproteota (I3R355, MD1ID33 and Q5JI39, respectively), suggesting a more compact domain for MD1ID33 than for I3R355 and Q5JI39. In Fig. 4, we compare the USP domain surface and their electrostatic potential, showing that halobacterium species present a much higher negatively charged surface (red) than other microorganism groups. The origins of the decrease in nonpolar surface in halophilic proteins, and one of the most striking differences between the surfaces of the halophilic and nonhalophilic enzymes, is a significant reduction in the percentage of hydrophobic surface accessible area due to lysine side chains as has been described in glucose dehydrogenase from H. mediterranei (Britton et al., 2006).
Fig. 4. Surface electrostatic potential representation of single USP proteins from each group. Halobacterium proteins present much higher negatively charged surface in comparison with other non-halophilic species. The M1ID33 protein model presents a small tail at the bottom of the structure that corresponds to the N-terminus, which is flexible and does not contribute to the core structure. Red: negative charge residues, blue: positive charge residues and white: non-charge residues
Several microorganisms also present USP-USP proteins, which contain two USP domains connected through a linker (Fig. 4). According to previous crystallography structures (PDB-ID: 5vb0) (Xu et al. 2016) and Alphafold models predictions, USP domains interact by forming an antiparallel intradomain β-sheet (one from each domain; Figs. 3 and 5A). Both domains can also contain the ATP binding domain (as in the example described in Figs. 3 and 5A) suggesting the possibility of binding two ATP molecules simultaneously. In addition, upon closer examination of their structure and symmetry in the available experimental structures at the Protein DataBase, these proteins may be able to oligomerize into dimers, allowing the simultaneous binding of four ATP molecules and the possibility of allosteric regulation.
Single USP protein models have also suggested the possibility of dimerization and tetramerization. A dimeric model generated with Alphafold2 from the single USP-domain protein, I3R355, shows the formation of an antiparallel interchain β-sheet for dimer formation similarly to that of the USP-USP proteins (Fig. 5B). A tetrameric model also suggests the possibility of tetramerization of single USP proteins and the formation of heterooligomers (Figure S2).
The models studied in this manuscript are generated by artificial intelligence. Although they show a high level of confidence, domain oligomerization and ATP binding in degenerative motifs still need to be confirmed experimentally.
Fig. 5USP domain interactions. (A) USP-USP protein structure of I3R6M7 predicted by Alphafold. One domain is represented in blue, the other in pink and the linker between domains is represented in grey. The antiparallel β-sheet involved in the domain interaction is represented in darker colours. (B) Single protein structure of I3R355 predicted as a dimer by Alphafold. Both structures present the antiparallel β-sheet between the two USP domains, regardless of whether they are single or double USP domain proteins. One subunit is represented in blue, and the other in pink. The antiparallel β-sheet involved in the oligomerization is represented in darker colours
Biotechnological applications
USPs play an essential role in the survival and adaptation of organisms to diverse environmental stresses. Their wide distribution and functional versatility in pathogens, plants, bacteria and archaea position them as valuable tools in biotechnology.
In pathogens, such as M. tuberculosis (Banerjee et al. 2024) or S. typhimurium (Liu et al. 2007), USPs enable them to adapt to the conditions found in the human niche, including oxidative or osmotic stress, nutrient starvation or acid stress, thereby facilitating infection. Similarly, bacterial USPs respond to various stresses, including nutrient deprivation, osmotic stress, and exposure to heavy metals (Kvint et al. 2003). On the other hand, USPs in plants are involved in a wide range of stress responses, including drought, salinity, heat, and pathogen attacks (Chi et al. 2019). In the case of Archaea, USPs are directly related to their adaptation to extreme environments such as high salinity. Recently, the interaction of SaUspA with threonine/serine phosphatase PP2 has been described (Ye et al. 2020a, b).
The versatility of USPs across different organisms offers numerous biotechnological applications. Focusing on Archaea, further investigation into their structural characteristics, putative functions, protein-protein interactions, and molecular mechanisms could unlock new applications in biomedicine, agriculture and bioremediation. In biomedicine, novel archaeal USPs structures could be used to develop antimicrobial therapies as Archaea and Eukarya organisms share replication, transcription processes, and molecular machinery. In general, halophilic archaea exhibit an increase in USP encoded in their genomes, which appears to be related to the osmotic stress. It has been suggested that the basal expression of a minimal number of USPs could be attributed to the constant need for a high salt concentration, which is crucial for the survival of halophilic organisms like H. mediterranei (Matarredona et al. 2024). Following this hypothesis, introducing or increasing USP genes in plants could enhance their salinity tolerance during droughts, which are increasing because of climate change. Archaeal USPs could be useful in industrial processes or environmental bioremediation, where archaea can be engineered to enhance their tolerance under harsh conditions or to absorb heavy metals more effectively in contaminated environments, respectively. However, although the number of USPs identified in Archaea has increased significantly, their mechanisms of action remain largely unknown, highlighting the need for further research to explore their potential applications fully.
Conclusion
The distribution and density of USPs in Archaea reflect adaptive strategies to extreme environments. Halophilic families, such as Haloferacaceae, have high levels of USPs, supporting survival in fluctuating salinities, whereas methanogens, such as Methanobacteriaceae, have fewer USPs, relying on stable anaerobic conditions. Thermophilic families, including Thermoproteaceae, show moderate USP investment, adapting to different stressors. The heterogeneity of structural motifs in Halobacteriota,* Methanobacteriota* and Thermoproteota suggests a correlation between domain complexity and stress diversity. High USP diversity enables robust responses to environmental variation, highlighting archaeal resilience and potential biotechnological applications in stress tolerance, industrial processes and biomedicine.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Matarredona L, Camacho M, Zafrilla B, Bonete MJ, Esclapez J (2020) The role of stress proteins in Haloarchaea and their adaptive response to environmental shifts. Biomolecules. 29;10(10):1390. 10.3390/biom 1010139010.3390/biom 10101390 PMC 760113033003558 · doi ↗ · pubmed ↗
- 2Schweikhard ES, Kuhlmann SI, Kunte HJ, Grammann K, Ziegler CM (2010) Structure and function of the universal stress protein Tea D and its role in regulating the ectoine transporter Tea ABC of Halomonas elongata DSM 2581(T). Biochemistry. 16;49(10):2194– 204. 10.1021/bi 901752210.1021/bi 901752220113006 · doi ↗ · pubmed ↗
- 3Siegele DA (2005) Universal stress proteins in Escherichia coli. J Bacteriol 187 186253–6254. 10.1128/JB.187.18.6253-6254.200510.1128/JB.187.18.6253-6254.2005 PMC 123665916159755 · doi ↗ · pubmed ↗