Suppression of har-1/CHCHD10 phenotypes for ALS-FTD therapy discovery
Audrey Labarre, Ericka Guitard, Gilles Tossing, J Alex Parker

TL;DR
This study explores using a worm model to find therapies for ALS-FTD by testing compounds that alleviate disease-related symptoms.
Contribution
The study introduces a novel worm model for ALS-FTD and identifies potential therapeutic agents.
Findings
Pioglitazone and Lacticaseibacillus rhamnosus HA-114 reduce har-1 mutant phenotypes in worms.
The har-1 model is suitable for modifier and drug screening studies for ALS-FTD.
The model supports high-throughput screening and microbiome research for therapeutic discovery.
Abstract
Mutations in CHCHD10 are linked to a variety of neurodegenerative diseases, including amyotrophic lateral sclerosis and frontotemporal dementia (ALS-FTD). The Caenorhabditis elegans orthologue of CHCHD10 is har-1 , and we investigated whether har-1 mutants could be used for therapeutic discovery in ALS-FTD. Our results show that the small molecule pioglitazone and the probiotic Lacticaseibacillus rhamnosus HA-114 can alleviate har-1 mutant phenotypes. These findings suggest that har-1 mutants are suitable for modifier screens and could be adapted for high-throughput drug screening and microbiome studies to aid in discovering therapies for ALS-FTD.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
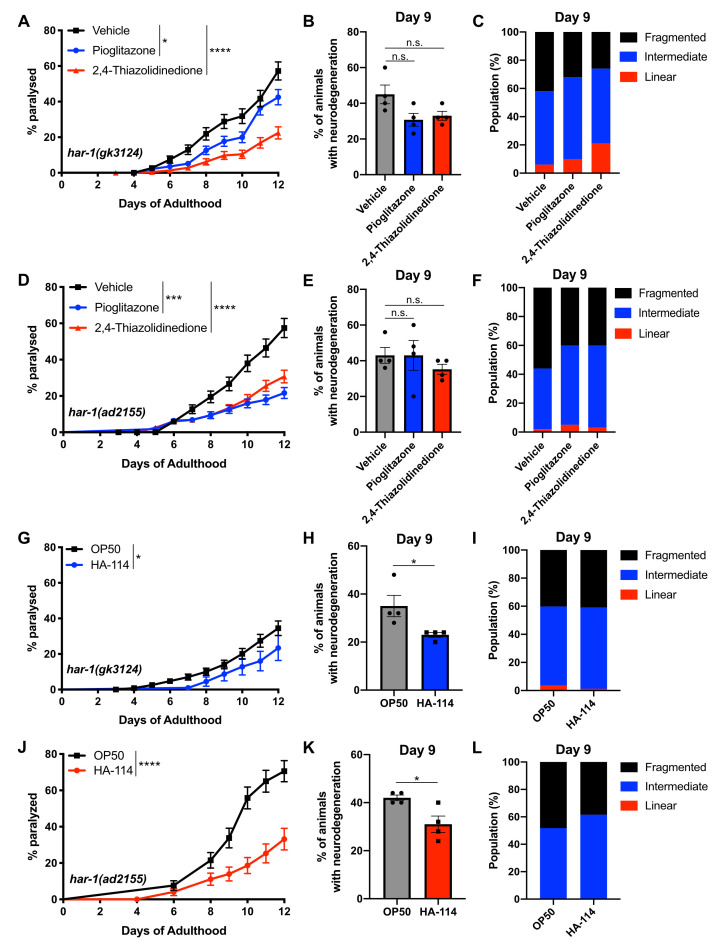 Figure 1
Figure 1|
|
|
|
|
|
Wild type
|
CGC |
|
|
|
CGC |
|
|
|
CGC |
|
|
|
CGC |
|
|
|
CGC |
|
|
|
Dr. Michael M. Francis (University of Massachusetts, Worcester, MA) |
|
|
|
Parker Lab |
|
|
[myo-3p::TOM20::mRFP +
|
Parker Lab |
|
|
|
Parker Lab |
|
|
|
Parker Lab |
|
|
|
|
|
Pioglitazone |
CDS021593 |
Sigma-Aldrich |
|
2,4-Thiazolidinedione |
375004 |
Sigma-Aldrich |
|
|
|
|
|
CGC |
|
|
Lallemand Health Solutions |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAmyotrophic Lateral Sclerosis Research · Ubiquitin and proteasome pathways · Heat shock proteins research
Description
har-1 * mutations lead to impaired motility and abnormal mitochondrial function in * C. elegans * (Woo et al., 2017). The robustness of these phenotypes suggests that they may be suitable for modifier screening. We have previously used * C. elegans * as a screening tool to identify small molecules that suppress paralysis and neurodegeneration phenotypes in multiple disease models, including ALS (Doyle et al., 2021; Fardghassemi et al., 2021; Patten et al., 2017; Schmeisser et al., 2017). However, few compounds emerged as specific modulators of mitochondrial genes or pathways. Recently, pioglitazone (PGZ), an oral antidiabetic drug from the thiazolidinedione family, was identified as a compound specific to disorders characterized by mitochondrial impairment, with the ability to reduce oxidative stress and improve mitochondrial bioenergetics (Mollo et al., 2019; Paciello et al., 2018; Yonutas et al., 2020). Interestingly, pioglitazone has also been recognized as a potential therapeutic for ALS in various models, including Drosophila and mice, and has been the subject of a clinical trial with promising results (Joardar et al., 2015; Schütz et al., 2005). Similarly, 2,4-thiazolidinedione (TZD), the core member of the thiazolidinedione family and the one with the simplest chemical structure, has been identified as a mitochondrial modulator with neuroprotective effects, particularly in Parkinson's disease (Wang et al., 2017).
Therefore, we tested PGZ and TZD in * har-1 ( gk3124 ) * and * har-1 ( ad2155 ) * worms to determine whether these compounds had protective effects. We observed that TZD treatment significantly reduced the paralysis phenotype in * har-1 ( gk3124 ) * , while PGZ had a more modest impact on the motility phenotype ( ** Fig. 1A ** ). However, the reduction in motility defects was not accompanied by a significant decrease in GABAergic neurodegeneration ( ** Fig. 1B ** ). PGZ partially helped restore mitochondrial health by decreasing the number of animals with “fragmented” morphology ( ** Fig. 1C ** ). TZD had a more pronounced effect, as treated * har-1 ( gk3124 ) exhibit * ed a substantial increase in animals with “linear” mitochondria and a decrease in those scored as “fragmented” by day 9 of adulthood ( ** Fig. 1C ** ). Both PGZ and TZD treatment reduced paralysis in * har-1 ( ad2155 ) * mutants ( ** Fig. 1D ** ), but they were ineffective at reducing motor neuron degeneration ( ** Fig. 1E ** ). Nonetheless, PGZ and TZD improved mitochondrial health in * har-1 ( ad2155 ) * animals by significantly decreasing the numbers of both “intermediate” and “fragmented” scored animals ( ** Fig. 1F ) ** . These results suggest that * har-1 ( gk3124 ) * and * har-1 ( ad2155 ) * worms exhibit a variety of features associated with ALS that can be modulated by FDA-approved drugs like PGZ, making these models useful tools for screening therapeutics.
Beyond small-molecule interventions, interest in the microbiome and dietary supplementation has arisen as a potential means to modulate disease phenotypes. Several studies have identified dysbiosis in ALS models and patients (Blacher et al., 2019; Burberry et al., 2020; Figueroa-Romero et al., 2020; Wu et al., 2015). Bacterial metabolites, including short-chain fatty acids like butyrate, play a role in energetic balance through mitochondrial processes (Besten et al., 2013). Although the functions of CHCHD10 remain unclear, mutations in * har-1 * /CHCHD10 may influence energy production by altering mitochondrial function. Furthermore, energy deficits have been consistently recognized in ALS and are widely acknowledged as an integral aspect of the disease (Vandoorne et al., 2018).
In a laboratory context, worms are often cultivated on Petri plates containing *E. coli * OP50 as their primary food source and the main component of their natural microbiome. To determine whether dietary probiotic interventions could modify * har-1 * phenotypes, we tested the probiotic strain *Lacticaseibacillus rhamnosus * HA-114 (hereafter referred to as HA-114) on both * har-1 ( gk3124 ) * and * har-1 ( ad2155 ) * mutants. We previously identified this strain as neuroprotective in transgenic models of ALS and Huntington's disease and speculated whether this neuroprotection extended to our * har-1 * models (Labarre et al., 2022). We found that HA-114 significantly rescued paralysis phenotypes ( ** Fig. 1G, 1J ** ) and motor neuron degeneration ( ** Fig. 1H, K ** ) in the * har-1 * mutants compared to worms fed control OP50 bacteria. Interestingly, HA-114 treatment did not affect mitochondrial morphology in * har-1 ( gk3124 ) * worms ( Fig.1I ). However, HA-114 treatment reduced the proportion of * har-1 ( ad2155 ) * mutants with a fragmented mitochondrial network ( ** Fig. 1L ** ). These findings suggest that dietary probiotic interventions in nematodes can modify paralysis and neurodegenerative phenotypes, and that HA-114 effectively reverses these phenotypes in age-dependent models of neurodegeneration associated with mitochondrial dysfunction. Collectively, these data highlight the potential of * har-1 * mutants for therapeutic screens and encourage further investigation. Combined approaches should perhaps be explored to fully suppress the range of phenotypes observed in * har-1 * mutants.
Methods
** C . elegans maintenance and strains **
Standard culturing techniques for * C. elegans * were used (Stiernagle, 2006) . Worms were kept on NGM agar plates with E. coli OP50 at 20 °C. Strains from the * C. elegans * Genetics Center (University of Minnesota) included N2 , VC3169 * har-1 ( gk3124 ) * , DA2155 ( * har-1 ( ad2155 ) * ), CB307 ( * unc-47 ( e307 ) * ), PS6192 ( * syIs243[myo-3p::TOM20::mRFP + unc-119 (+) + pBS Sk+] * ), and TU3311 ( * uls60[ unc-119 p::YFP + unc-119 p:: sid-1 ] * ). The IZ629 strain ( * ufsl34[P unc-47 p::mCherry] * ) was provided by Dr. Michael M. Francis (University of Massachusetts). All mutant strains underwent four rounds of outcrossing to N2 . Strains were verified by PCR or sequencing. Some experiments were conducted by dissolving pioglitazone (20 uM, Sigma-Aldrich) or 2,4-Thiazolidinedione (20 uM, Sigma-Aldrich) into the NGM plates.
Solid media paralysis assay
From days 1 to 12 of adulthood, 40 L4 worms were transferred daily to fresh NGM plates and monitored for paralysis. Worms were deemed dead if they didn't move their heads when tapped and showed no pharyngeal pumping. Paralysis was the inability to move when prodded. Each experiment was conducted three times in duplicate at 20 °C.
Neurodegeneration assay
On day 9 of adulthood, worms were selected for in vivo motor neuron imaging to assess neural processes for gaps. Animals were immobilized in 5 mM levamisole in M9 buffer on 2% agarose pads. Imaging was done with a Zeiss Axio Imager M2 microscope with a 20X objective and 1.5 Optovar, using AxioVs40 software. Over four studies, at least 100 worms were scored per condition.
Mitochondrial morphology analysis
The structure of body-wall mitochondria was analyzed using the PS6192 ( * Pmyo-3::TOM20::mRFP + unc-119 (+) + pBS SK+ * ) strain, which expresses a red fluorescent protein targeted to the outer mitochondrial membrane. This strain was crossed with both * har-1 * genotypes to assess mitochondrial health. Age-synchronized adults at day 9 were immobilized on 2% agarose pads with 5 mM levamisole. Imaging utilized a Zeiss Axio Imager M2 microscope with a 40X objective and 1.5 Optovar, using AxioVs40 4.8.2.0 software. Mitochondria were classified as linear, intermediate, or fragmented based on Lu et al. (2011) criteria. At least 100 worms were scored per condition across four independent experiments.
Statistics
Survival curves for paralysis, lifespan, and stress resistance were compared using the Log-rank test with 60–100 worms examined per condition at least three times. For the neurodegeneration assay, one-way ANOVA with Dunnett's multiple comparisons was used. Two-way ANOVA with Dunnett's was used for mitochondrial morphology. Data were expressed as mean ± S.E.M., except mitochondrial morphology data, where error bars were removed for clarity. All statistical evaluations were conducted with GraphPad Prism v8.
Reagents
**: **
**: **
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1den Besten G van Eunen K Groen AK Venema K Reijngoud DJ Bakker BM 201372 The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism.J Lipid Res 5490022-22752325234010.1194/jlr.R 03601223821742 PMC 3735932 · doi ↗ · pubmed ↗
- 2Blacher E Bashiardes S Shapiro H Rothschild D Mor U Dori-Bachash M Kleimeyer C Moresi C Harnik Y Zur M Zabari M Brik RB Kviatcovsky D Zmora N Cohen Y Bar N Levi I Amar N Mehlman T Brandis A Biton I Kuperman Y Tsoory M Alfahel L Harmelin A Schwartz M Israelson A Arike L Johansson MEV Hansson GC Gotkine M Segal E Elinav E 2019722 Potential roles of gut microbiome and metabolites in modulating ALS in mice.Nature 57277700028-083647448010.1038/s 41586-019-1443-531330533 · doi ↗ · pubmed ↗
- 3Burberry A Wells MF Limone F Couto A Smith KS Keaney J Gillet G van Gastel N Wang JY Pietilainen O Qian M Eggan P Cantrell C Mok J Kadiu I Scadden DT Eggan K 2020513 C 9orf 72 suppresses systemic and neural inflammation induced by gut bacteria.Nature 58278100028-0836899410.1038/s 41586-020-2288-732483373 PMC 7416879 · doi ↗ · pubmed ↗
- 4Doyle JJ Maios C Vrancx C Duhaime S Chitramuthu B Bennett HPJ Bateman A Parker JA 2021622 Chemical and genetic rescue of in vivo progranulin-deficient lysosomal and autophagic defects.Proc Natl Acad Sci U S A 118250027-842410.1073/pnas.202211511834140407 PMC 8237633 · doi ↗ · pubmed ↗
- 5Fardghassemi Y Maios C Parker JA 2021329 Small Molecule Rescue of ATXN 3 Toxicity in C. elegans via TFEB/HLH-30.Neurotherapeutics 1821933-72131151116510.1007/s 13311-020-00993-533782863 PMC 8423969 · doi ↗ · pubmed ↗
- 6Figueroa-Romero C Guo K Murdock BJ Paez-Colasante X Bassis CM Mikhail KA Pawlowski KD Evans MC Taubman GF Mc Dermott AJ O'Brien PD Savelieff MG Hur J Feldman EL 20191115 Temporal evolution of the microbiome, immune system and epigenome with disease progression in ALS mice.Dis Model Mech 1321754-840310.1242/dmm.04194731597644 PMC 6906635 · doi ↗ · pubmed ↗
- 7Joardar A Menzl J Podolsky TC Manzo E Estes PS Ashford S Zarnescu DC 20141128 PPAR gamma activation is neuroprotective in a Drosophila model of ALS based on TDP-43.Hum Mol Genet 2460964-69061741175410.1093/hmg/ddu 58725432537 PMC 4381760 · doi ↗ · pubmed ↗
- 8Labarre A Guitard E Tossing G Forest A Bareke E Labrecque M Tétreault M Ruiz M Alex Parker J 2022127 Fatty acids derived from the probiotic Lacticaseibacillus rhamnosus HA-114 suppress age-dependent neurodegeneration.Commun Biol 511340134010.1038/s 42003-022-04295-836477191 PMC 9729297 · doi ↗ · pubmed ↗