Allelic Variations in Phenology Genes in Club Wheat (Triticum compactum) and Their Association with Heading Date
Bárbara Mata, Adoración Cabrera

TL;DR
This study explores genetic variations in wheat genes that control growth timing and how they affect when the plant starts to head, which is important for breeding.
Contribution
The study identifies 10 unique gene profiles in Triticum compactum and links them to heading date differences under various conditions.
Findings
58.4% of Triticum compactum accessions had a spring growth habit with dominant VRN1 alleles.
The Ppd-D1b allele was found in 95.5% of accessions, indicating photoperiod sensitivity.
21 SSR markers revealed high genetic diversity with an average of 3.2 alleles per locus.
Abstract
The allelic diversity within genes controlling the vernalization requirement (VRN1) and photoperiod response (PPD1) determines the ability of wheat to adapt to a wide range of environmental conditions and influences grain yield. In this study, allelic variations at the VRN-A1, VRN-B1, VRN-D1 and PPD-D1 genes were studied for 89 accessions of Triticum compactum from different eco-geographical regions of the world. The collection was evaluated for heading date in both field and greenhouse experiments under a long photoperiod and without vernalization. Based on heading date characteristics, 52 (58.4%) of the genotypes had a spring growth habit, and all of them carried at least one dominant VRN1 allele, while 37 (41.6%) accessions had a winter growth habit and carried the triple recessive allele combination. The photoperiod-sensitive Ppd-D1b allele was detected in 85 (95.5%) accessions and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
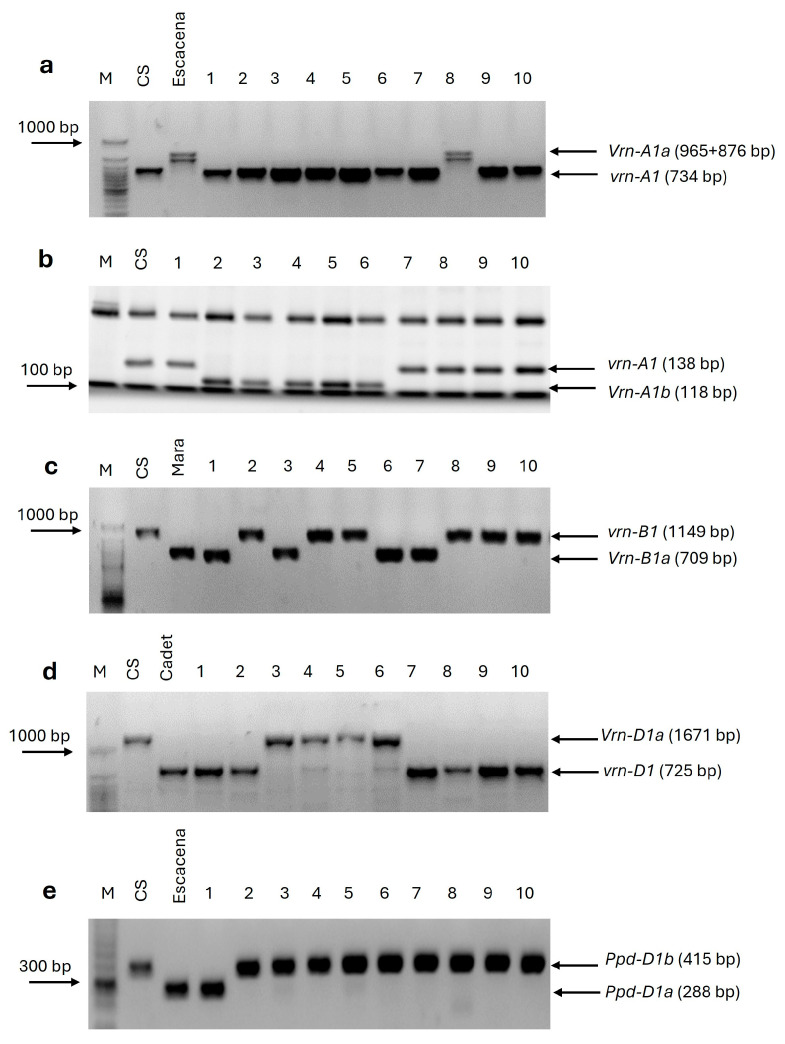 Figure 1
Figure 1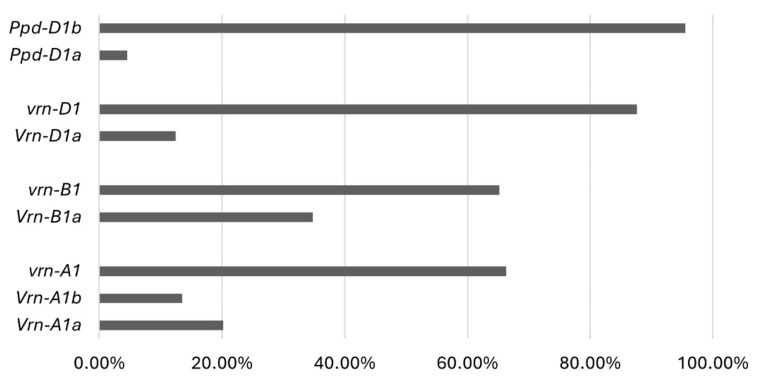 Figure 2
Figure 2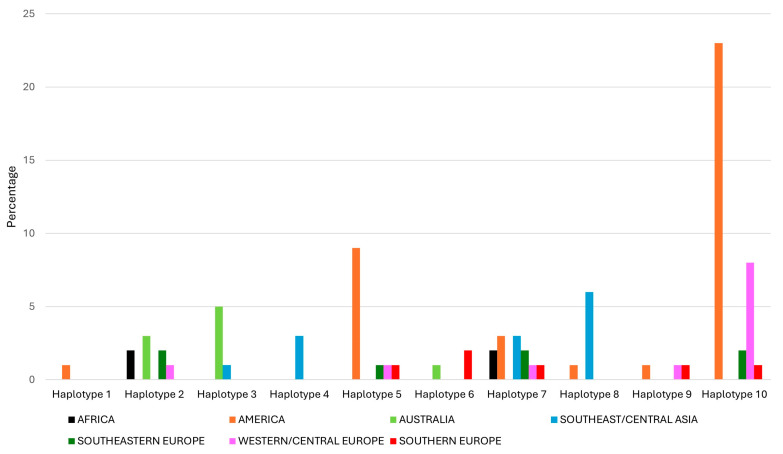 Figure 3
Figure 3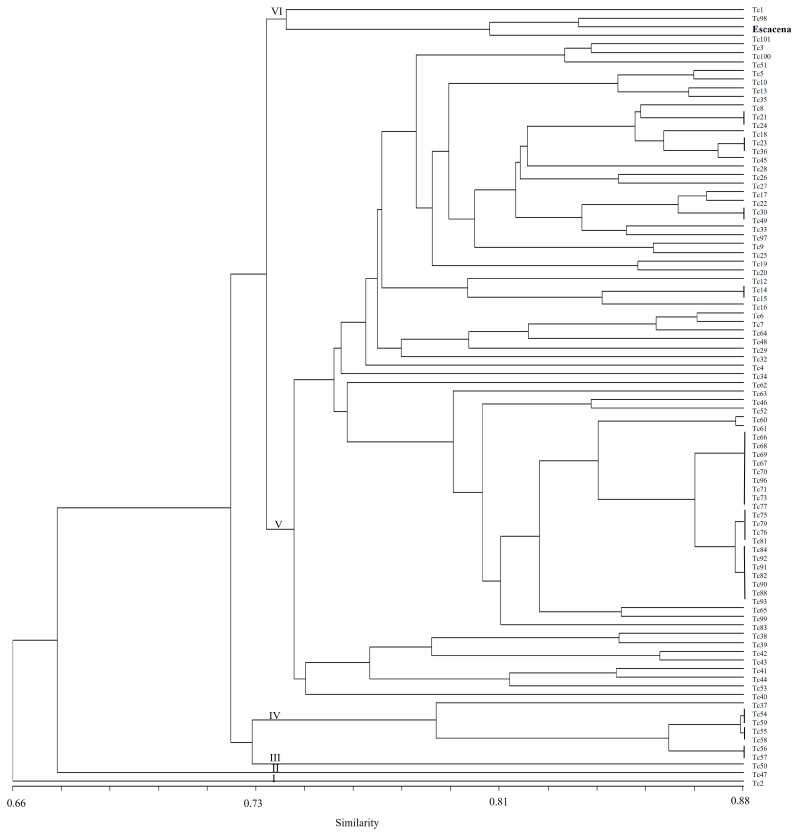 Figure 4
Figure 4 Figure 5
Figure 5- —Spanish Research Agency (Ministry of Science, Innovation and Universities)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsWheat and Barley Genetics and Pathology · Genetics and Plant Breeding · Genetic Mapping and Diversity in Plants and Animals
1. Introduction
Club wheat (Triticum compactum [Triticum aestivum ssp. compactum (Host) Mac Key, 2n = 6x = 42, A^u^A^u^BBDD]) is a subspecies of common wheat (T. aestivum ssp. aestivum, 2n = 6x = 42, A^u^A^u^BBDD) that is distributed throughout the Old World [1]. The main areas for cultivation of club wheat are the U.S. Pacific Northwest and some areas of the Middle East (Afghanistan, Iran, Pakistan), Asia (Armenia, Turkey), Europe (Austria, Switzerland) and Australia [2,3]. It is a free-threshing wheat with rounded grains that differs from common wheat in certain morphological characteristics, including its compact spike due to shorter rachis segments. In addition, it has smaller grains than common wheat, but this smaller size is compensated for by a higher seed number [3]. The dominant C locus mapped on the short arm of chromosome 2D determines the compact spike [3,4] and also has a pleiotropic effect on many agronomic traits such as spike length, seed size and seed number, influencing crop yield [3,5]. Club wheat is commercially important for producing flours suitable for making cakes and pastries. Some of its quality characteristics, such as low gluten strength, low water absorption, low batter viscosity and low protein, result in high-quality biscuits and cakes [6,7]. Screening for genes related to agronomic traits in club wheat germplasm has shown variability for high molecular weight glutenin [8,9], waxy proteins [10] and stripe rust resistance [11]. It has also been suggested that club wheat cultivars are better adapted to some agroclimatic regions and may be more competitive in dryland areas than common wheat [2,3,12].
The development of new wheat varieties better adapted to water-limited environments is one of the main challenges for plant breeders. The use of the genetic resources available for wheat is key for obtaining new genotypes adapted to climate change. Species in the primary wheat gene pool, such as T. compactum, could be good candidates as gene sources for new allelic variants to enable the diversification of phenological characteristics of common wheat cultivars. The transition to flowering in cereals is mainly determined by temperature and photoperiod. In hexaploid wheat, the vernalization-induced flowering pathway is mainly regulated by the MADS-box transcription factor VERNALIZATION1 (VRN1) loci [13,14,15]. Allelic variants at the VRN-A1, VRN-B1 and VRN-D1 homeoloci map to the long arm of chromosomes 5A, 5B and 5D [16,17,18], respectively, and they are one of the main resources of genetic variation in vernalization requirements in wheat. The dominant VRN1 alleles conferring a spring growth habit result from insertions and/or deletions in the promoter and intron 1 regions [15,19,20,21]. PHOTOPERIOD (PPD1), a member of the pseudo-response regulator (PRR) family, is a key gene determining the sensitivity to day length in wheat [22,23]. PPD1 loci map to the short arm of chromosomes 2A, 2B and 2D, and among them, PPD-D1 is considered the most important photoperiod regulator in wheat [23,24]. The semi-dominant Ppd-D1a allele, characterized by a large deletion upstream of the coding region, leads to photoperiod insensitivity [25,26].
Studies on the diversity of phenology-related genes in T. compactum are scarce. The characterization of wheat germplasm for genes controlling heading date variations will provide a valuable tool for breeders in developing wheat varieties that can adapt to various environmental conditions. This study aimed to use functional molecular markers to evaluate the allelic variation in four major genes controlling phenology (VRN-A1, VRN-B1, VRN-D1 and PPD-D1) in 89 accessions of hexaploid wheat T. compactum and to study the effect of allele combination on heading date. The genetic diversity in the club collection was also estimated.
2. Results
2.1. Allelic Variation at the VRN1 and PPD-D1 Genes in the T. compactum Collection
Amplification of genomic DNA using specific primers to detect allelic variations in the promoter region of the VRN-A1 gene allowed the identification of two dominant alleles (Vrn-A1a and Vrn-A1b) and one recessive allele (vrn-A1). PCR products of 965 + 876 bp in length corresponded to the Vrn-A1a allele, while those 734 bp in length corresponded to Vrn-A1b (714 bp) or vrn-A1 (734 bp). To differentiate between these latter alleles, the PCR product was digested with the MspI restriction endonuclease, and the fragments were separated using polyacrylamide gels. We obtained two restriction patterns, one 119 bp long corresponding to the dominant Vrn-A1b allele and the other, 138 bp long, corresponding to the recessive vrn-A1 allele. Overall, 18 (20.2%) of the club wheat accessions carried the foldback element insertion characteristic of the Vrn-A1a allele, 12 (13.5%) the 119 bp restriction fragment characteristic of the Vrn-A1b allele and 59 (66.3%) the 138 bp fragment corresponding to the recessive vrn-A1 allele (Figure 1a,b).
To study allelic variation at the VRN-B1 gene, multiplex PCR was performed using three primers (Table S1). In principle, these primers can detect two dominant (Vrn-B1a and Vrn-B1b) alleles and one recessive (vrn-B1) allele [19]. Screening with these primers found two alleles in the club wheat collection studied: the dominant Vrn-B1a allele, which was identified in 31 (34.8%) accessions, and the recessive vrn-B1 allele, which was found in 58 (65.2%) (Figure 1c). The dominant Vrn-B1b allele was not detected in the T. compactum germplasm evaluated in this study.
To identify allelic variation at the VRN-D1 gene, multiplex PCR screening was carried out using three primers (Table S1). With these primers, two dominant (Vrn-D1a and Vrn-D1s) alleles and one recessive (vrn-D1) allele can be detected [19,27]. Two alleles were identified in the club wheat collection: the dominant Vrn-D1a allele in 11 (12.4%) accessions and the recessive vrn-D1 allele in 78 (87.6%) (Figure 1d). No T. compactum genotypes were found to carry the Vrn-D1s allele. In total, 52 (58.4%) accessions carried at least one of the dominant vernalization alleles, and 37 (41.6%) accessions carried recessive alleles at all three VRN1 vernalization loci.
Finally, two PCR products (Figure 1e) were obtained from the club wheat collection with the PPD-D1-specific primers: the dominant Ppd-D1a photoperiod-insensitive and the recessive Ppd-D1b photoperiod-sensitive alleles were present in four (4.5%) and 85 (95.5%) accessions, respectively. Figure 2 shows the allele frequencies of the major effect genes, VRN-A1, VRN-B1, VRN-D1 and PPD-D1, in the T. compactum germplasm studied. The allelic variants of vernalization and photoperiod sensitivity genes in each genotype of the club wheat collection studied are listed in Table S2.
2.2. Effect of VRN1 and PPD-D1 Allele Combinations on T. compactum Heading Date
Heading date was assessed by growing the club wheat collection under a long photoperiod without vernalization in both the greenhouse and the field. The data gathered was normally distributed under both growth conditions (Figure S1). Out of the 89 T. compactum genotypes tested, 37 (41.6%) were classified as having a winter growth habit. These accessions did not flower under either greenhouse or field conditions, and all of them carried recessive (vrn-A1-vrn-B1-vrn-D1) vernalization alleles at all three VRN1 loci. Table 1 shows the relationship of heading date with vernalization and photoperiod genotypes in the T. compactum germplasm under both greenhouse and field conditions.
A total of 10 phenology gene profiles (haplotypes) of four major genes (VRN-A1, VRN-B1, VRN-D1 and PPD-D1) were observed in the T. compactum germplasm collection (Table 1). Only one accession was found to carry the triple dominant Vrn-A1a-Vrn-B1a-Vrn-D1a (1.1%) allele combination at all three loci (haplotype 1). This accession also carried the semi-dominant Ppd-D1a allele for photoperiod insensitivity. We also observed combinations of two dominant alleles, namely, Vrn-A1a-Vrn-B1a (6.7%), Vrn-A1b-Vrn-B1a (10.1%) and Vrn-B1a-Vrn-D1a (3.4%). A single dominant allele was observed for Vrn-A1a (12.3%), Vrn-A1b (3.4%), Vrn-B1a (13.5%) and Vrn-D1a (7.9%). All these two-dominant and one-dominant-allele combinations carried the recessive Ppd-D1b allele for photoperiod sensitivity. The wild-type allele Ppd-D1b, causing photoperiod sensitivity, was present in 95.5% of the accessions, whereas only four accessions carried the semi-dominant photoperiod-insensitive Ppd-D1a allele (4.5%).
The analysis of variance revealed significant differences (p < 0.05) in mean heading date among the different spring allele combinations under both greenhouse and field conditions (Table 1). Under greenhouse conditions, the LSD test found no significant differences in the mean heading date among haplotype 1 (Vrn-A1a-Vrn-B1a-Vrn-D1a-Ppd-D1a), haplotype 2 (Vrn-A1a-Vrn-B1a-vrn-D1-Ppd-D1b), haplotype 3 (Vrn-A1b-Vrn-B1a-vrn-D1-Ppd-D1b), haplotype 4 (vrn-A1-Vrn-B1a-Vrn-D1a-Ppd1b), haplotype 8 (vrn-A1-vrn-B1-Vrn-D1a-Ppd1b) and the spring common wheat cultivar Escacena (Vrn-A1a-vrn-B1-Vrn-D1a-Ppd-D1a) used as control. The LSD test also showed that the club wheat genotypes carrying haplotype 6 (Vrn-A1b-vrn-B1-vrn-D1-Ppd-D1b) headed significantly later than those harboring triple or double dominant allele combinations at the VRN1 loci, but no significant differences were found in the mean heading date between haplotypes 6, 5 (Vrn-A1a-vrn-B1-vrn-D1-Ppd-D1b) and 7 (vrn-A1-Vvrn-B1a-vrn-D1-Ppd-D1b), all of them carrying one dominant allele at the VRN1 loci. From field data, the LSD test showed that the T. compactum accession with haplotype 1 carrying all the dominant alleles at both the VRN1 and PPD-D1 loci (Vrn-A1a-Vrn-B1a-Vrn-D1a-Ppd-D1a) and the spring common wheat cultivar Escacena used as a control headed significantly earlier than all the other spring allele combinations. Furthermore, although haplotype 5 (Vrn-A1a-vrn-B1-vrn-D1-Ppd-D1b) carrying a single dominant Vrn-A1a allele headed later than haplotype 4 (vrn-A1-Vrn-B1a-Vrn-D1a-Ppd1b) carrying two dominant alleles and haplotypes 6, 7 and 8 carrying one dominant allele (Vrn-A1b, Vrn-B1a or Vrn-D1a, respectively), the differences were not statistically significant under field conditions.
2.3. Geographical Distribution of VRN1 and PPD-D1 Alleles and Allele Combinations
The highest frequencies in the Vrn-A1a allele were found in countries from Southeastern Europe (42.9%), whereas Vrn-A1b was mainly detected (66.7%) in germplasm from Australia (Table 2). Our results also showed that the Vrn-A1a allele was not present in accessions from Africa or Southeast/Central Asia, whereas the Vrn-A1b allele was not detected in accessions from Southeastern, Western or Central Europe. The most frequent allele at the Vrn-A1 locus was the recessive vrn-A1 allele (66.3%), and it was detected in all geographical areas except Australia. The dominant Vrn-B1a allele was present in germplasm from all regions studied, and it showed the highest frequency in accessions from Africa (100%) and Australia (88.9%). The Vrn-D1a allele was most frequent in Southeast Asia (69.2%), though it was also detected in genotypes from the USA. Our studies also showed a low frequency (4.5%) of the Ppd-D1a allele in the club wheat germplasm evaluated in this study. This allele was only detected in two accessions from the USA, one from Spain and one from Switzerland. By contrast, the photoperiod-sensitive Ppd-D1b seems to be much more common and was present at high frequencies in accessions from all geographical regions tested.
Figure 3 shows the distribution of the ten haplotypes by geographical origin. Haplotype 1 (Vrn-A1a-Vrn-B1a-Vrn-D1a-Ppd-D1a) carrying the dominant alleles for the VRN-A1, VRN-B1, VRN-D1 and PPD-D1 genes was detected in the cultivar Calorwa from the USA. Haplotypes 3 and 6 were mainly distributed in Australia, Southeast/Central Asia and Southeastern Europe. Haplotype 4 (vrn-A1-Vrn-B1a-Vrn-D1a-Ppd-D1b) only occurred in Southeast/Central Asia, specifically in accessions from Kazakhstan and Kyrgyzstan. Haplotype 7 (vrn-A1-Vrn-B1a-vrn-D1-Ppd-D1b) was the most common of the spring genotypes, and it was also the most universally distributed across all continents, except Australia. Haplotype 8 was mainly found in Southeast/Central Asia. Finally, winter genotypes (haplotypes 9 and 10) were common in America and Southern Europe, Western/Central Europe and Southeastern Europe, while they were absent in Africa, Australia and Southeast/Central Asia.
2.4. Simple Sequence Repeat Marker and Cluster Analysis
A total of 183 alleles were detected across all 89 club wheat accessions analyzed in this study using 21 simple sequence repeat (SSR) markers (Table 3; Table S3). The allele sizes ranged from 54 (Xgwm44) to 280 bp (Xgwm397). The mean number of alleles per marker was 3.2, ranging from 1 to 8.9. All the SSR markers, except Xgwm44 and Xgwm391, were considered informative markers (PIC > 0.5). The mean PIC value estimated was 0.84. The phylogenetic relationship among the T. compactum accessions was investigated by cluster analysis.
The dendrogram indicates six main groups (Figure 4). The first group is separated from all the others and contains one accession (Tc2), a landrace from Kyrgyzstan. Groups II and III also contain just one accession each (Tc47 and Tc50, respectively), both from the USA. Group IV consists of seven accessions (T37, T54, T55, T56, T57, T58 and T59), all of them landraces from Pakistan. Group V was divided into two subgroups: Group V-1 is composed of eight accessions, five of them from Australia (Tc41, Tc42, Tc43, Tc,44; Tc53), two from Africa (Tc39, Tc40) and one from Pakistan (Tc38), and Group V-2 is a wider cluster containing 68 accessions from all eco-geographical regions studied except Africa. Finally, Group VI comprises three accessions (Tc1, Tc98 and Tc101), one collected in Mexico, one in (the then) Czechoslovakia and one in Georgia. The common wheat cultivar Escacena was also included in this cluster.
Clusters I, IV, V-1 and VI contained accessions with a spring growth habit (haplotypes 1 to 8) and clusters II and III accessions with a winter growth habit (haplotypes 9 and 10), while cluster V-2 contained both spring and winter growth habit accessions.
3. Discussion
The use of diagnostic DNA markers has allowed us to establish the allelic composition of VRN1 and PPD-D1 genes affecting the vernalization requirement and photoperiod response, respectively, in a T. compactum germplasm collection representing a wide range of eco-geographical regions. Within the screened germplasm, the highest allele frequency (87.6%) was recorded for the recessive vrn-D1 allele, followed by the recessive vrn-A1 (66.3%) and vrn-B1 (65.2%) alleles. The photoperiod-sensitive Ppd-D1b allele was detected in 85 (95.5%) accessions and the insensitive Ppd-D1a allele in four (4.5%). The Ppd-D1a allele was found in accessions from Spain, Switzerland and the USA. In common wheat, the Ppd-D1a allele has been mostly found in Central and Southern European [26] and Asian [28,29,30] varieties, whereas the photoperiod-sensitive Ppd-D1b allele was most common among wheat varieties from the USA and Canada [28].
Among the genotypes with at least one dominant VRN1 allele, the frequencies of the different alleles varied across geographical regions (Table 2, Figure 3). The Vrn-A1a allele has been found to be the most frequent dominant allele in common spring wheat cultivars in many parts of the world [15,21,31,32]. In this study, the Vrn-B1a allele was the most frequent dominant allele in spring T. compactum accessions evaluated, with a frequency of 34.8%, followed by Vrn-A1a (20.2%), Vrn-A1b (13.5%) and Vrn-D1a (12.4%). In common wheat, the Vrn-A1b allele was found in commercial cultivars from Southern Europe [33] and landraces from Turkey [34]; in the T. compactum germplasm collection evaluated in this study, this allele occurred in all regions tested except Southeastern Europe and Western/Central Europe, being most frequent in Australia. The Vrn-A1b allele was also found in some genotypes of club wheat from Afghanistan [35].
We did not find the dominant Vrn-B1b allele in any of the T. compactum accessions studied, consistent with previous reports [35,36]. Dragovich et al. [35] found that 66.7% of T. compactum accessions contained haplotype 7 (vrn-A1-Vrn-B1a-vrn-D1-Ppd-D1b); they considered this haplotype to be species-specific and also suggested that this allelic composition was characteristic of Asian club wheats. In accordance with their results [35], in the collection we studied, haplotype 7 was the most frequent (13.5%) among the T. compactum accessions with a spring growth habit. On the other hand, we also found that haplotype 7 was the most widely distributed, occurring in all regions tested except Australia. In this study, we found that haplotype 4 (vrn-A1-Vrn-B1a-Vrn-D1a-Ppd-D1b) was the only allelic combination found in Southeast/Central Asia (Figure 3), although it was also present in genotypes from Africa, Bulgaria, the UK and the USA.
Three alleles (Vrn-D1a, Vrn-D1s and vrn-D1) were detected at the VRN-D1 gene in a multiplex reaction with the three primers used (Table S1). In the T. compactum germplasm analyzed here, the dominant Vrn-D1a allele and the recessive vrn-D1 allele were identified. The Vrn-D1s allele, caused by DNA transposon insertion in intron 1, was identified in T. aestivum ssp. spelta [27]. The same authors also found this allele in one accession of T. compactum and concluded that it is not specific to hexaploid T. spelta. In this study, we did not find the Vrn-D1s allele in any of the accessions evaluated. On the other hand, this allele was the only dominant allelic variant at the VRN-D1 locus found in a Spanish spelt collection, supporting the hypothesis that Vrn-D1s is specific to T. spelta [37].
Based on the heading date characteristics of the club wheat accessions assessed, 52 (58.4%) of the genotypes had a spring seasonal growth habit and 37 (41.6%) a winter growth habit. Accordingly, analysis with gene-specific molecular markers confirmed the presence of the recessive alleles at all three VRN1 loci in winter genotypes, whereas at least one dominant VRN1 allele was present in spring genotypes. The LSD test revealed significant differences in mean heading date among the spring T. compactum genotypes, showing that the allele combinations at VRN1 influenced seed head emergence under both greenhouse and field conditions, as previously reported in common wheat [38,39].
The mean heading date did not differ significantly between the local spring wheat cultivar Escacena (Vrn-A1a-vrn-B1-Vrn-D1a-Ppd-D1a) and the T. compactum cultivar Calorwa carrying the dominant (Vrn-A1a-Vrn-B1a-Vrn-D1a-Ppd-D1a) combination for both VRN1 and PPD-D1 alleles (haplotype 1) under greenhouse or field conditions. All haplotypes headed significantly later than Calorwa under field conditions; however, this cultivar was the only T. compactum accession carrying the dominant alleles for both VRN1 and PPD-D1 genes. Therefore, we cannot rule out the possibility that the significant LSD data are not statistically representative due to the small sample size. In wheat, alleles other than those studied here have been found to influence heading date [27,40,41,42] as well as copy number variations in VRN1 alleles [43,44,45], and hence, we cannot rule out such alleles affecting the phenology of T. compactum accessions.
Genetic diversity in the T. compactum genotypes was assessed by capillary electrophoresis using 21 genomic SSR markers covering the A, B and D genomes. All the primers used in this study were designed for common wheat, and all of them successfully amplified products in T. compactum. This result is consistent with that previously found by Zhang et al. [46], who investigated the transferability of sequence-tagged SSR markers from common wheat to 18 related grass species and found full transferability (100%) for T. compactum, supporting the genetic proximity of these subspecies. The dendrogram obtained in this study did not separate the T. compactum accessions from the common wheat cultivar Escacena. Specifically, this cultivar clustered in the same group as accessions Tc1, Tc98 and Tc101, providing evidence for the genetic proximity of club wheat and common wheat germplasm [46].
A total of 183 alleles were detected from 21 SSR markers among the 89 T. compactum genotypes. The Xgwm565 molecular marker was the most informative SSR marker, with a PIC value of 0.92, identifying 21 alleles (Table 3). The mean number of different alleles was 8.7, with a mean number of alleles per locus of 3.2. The mean number of alleles per locus obtained in different germplasm collections of common wheat varies from 2.5 [47,48] and 3.1 [46] to 5.05 [49], 5.4 [50] or 6.5 [51] alleles per locus. The mean PIC value calculated in T. compactum in this study was 0.84, which is higher than the values of 0.4 [46], 0.43 [50] and 0.69 [48] obtained with SSRs in common wheat in previous studies. Both the PIC value and the mean number of alleles per locus revealed a high level of genetic diversity in the T. compactum germplasm collection. The T. compactum accessions included in this study were from 24 different countries. This broad geographic distribution of the club wheat accessions in the collection may explain the high diversity observed. This diversity is also seen in spike and seed morphology (Figure 5).
On the other hand, we found accessions with identical SSR patterns that are clustered together in the dendrogram, and they also share both haplotype and geographical origin. For instance, Tc56 and Tc57 are both from Pakistan, and Tc14 and Tc15 are from Kazakhstan. These findings suggest that certain accessions might be duplicated and that SSR studies can aid in detecting duplicate accessions.
4. Materials and Methods
4.1. Plant Material
Eighty-nine accessions of T. compactum obtained from the National Small Grains Collection (Aberdeen, ID, USA) were employed in this study (Table S2). The collection included landraces, cultivars and breeding materials from 23 countries across the world. Spring wheat cultivars, namely, Chinese Spring, Cadet, Escacena and Mara, were used as controls.
4.2. Phenotyping
Greenhouse and field experiments were carried out in the 2023–2024 growing season in Córdoba, in southern Spain. For the greenhouse experiments, two germinated seeds for each accession were sown individually in plastic pots and grown at 20–25 °C under long-day conditions (18 h day) without vernalization. The heading date was measured in days from emergence until 50% of the spike had emerged from the flag leaf on the main stem, and the mean was calculated from the two plants grown per accession.
For evaluating the ear emergence time of club wheat accessions in the field, the seeds were sown on 1 March 2024, that is, under long-day conditions, in a randomized complete block design with two replications without vernalization. Each experimental unit was a single-row plot (50 cm long) containing at least 20 plants, with a 35 cm space between adjacent plots. The heading date was measured in 50% of the plants in the plot, and as in the greenhouse experiments, by counting the number of days from emergence until 50% of the spike had emerged from the flag leaf.
4.3. Genotyping
PCR Amplification of Allele-Specific Markers
Total genomic DNA was isolated from frozen young (four-leaf stage) plants using the cetyltrimethylammonium bromide method [52]. To prepare DNA samples for each accession, equal amounts of leaves from three individual plants of each accession were bulked. The samples were stored at −20 °C until amplification through PCR. The concentration of DNA in each sample was estimated using a NanoDrop 1000 Spectrophotometer (Thermo Scientific, Waltham, MA, USA).
Genotypes were characterized for the VRN1 (VRN-A1, VRN-B1 and VRN-D1) and PPD-D1 genetic loci using a set of functional molecular markers. Dominant (Vrn-A1a and Vrn-A1b) and recessive (vrn-A1) alleles of the VRN-A1 locus were detected using the primers VRN1AF and VRN1-INTR1R, as described by Yan et al. [15]. To distinguish between Vrn-A1b and vrn-A1, the PCR product was digested with restriction endonuclease MspI as in Shcherban et al. [33], and DNA fragments were separated on polyacrylamide gels. Allelic variants of the VRN-B1 gene were identified using three gene-specific primers (Intr1/B/F, Intr1/B/R3 and Intr1/B/R4), a combination linked to intron 1 in a multiplex PCR reaction, which allows detection of Vrn-B1a, Vrn-B1b and vrn-B1 [12,19]. Primers Intr1 and Intr1/B/R3 were used to differentiate between Vrn-B1a and Vrn-B1c [33].
To detect VRN-D1 intron 1 alleles, a multiplex PCR was performed using three primers (Intr1/D/F, Intr1/D/R3 and Intr1/D/R4). This PCR makes it possible to distinguish between Vrn-D1a, Vrn-D1s and vrn-D1 allelic variants [19,27]. Further, three specific primers (Ppd-D1-F1, Ppd-D1-R1 and Ppd-D1-R2) were used in a multiplex PCR assay to identify both the photoperiod-insensitive allele Ppd-D1a and photoperiod-sensitive allele Ppd-D1b of the PPD-D1 gene, respectively [25]. The primer sequences used in this study and annealing temperatures for each primer combination are listed in Table S1. PCR was performed with 40 ng of template DNA in a 25 µL volume reaction mixture containing 5 µL of 1× PCR buffer, 0.4 µM of each primer, 1.5 mM of MgCl_2_, 0.4 mM of dNTPs and 0.25U of Taq DNA polymerase (Promega, Madison, WI, USA). The PCR conditions were as follows: 4 min at 94 °C, followed by 35 cycles of denaturation at 94 °C for 45 s, annealing for 45 s, extension at 72 °C for 1 min and then a final extension at 72 °C for 5 min. Amplified products were resolved on 2.0% agarose gels or separated by polyacrylamide gels (10% w/v, C: 2.67%) and stained with Safe View Nucleic Acid Stain (NBS Biologicals, Huntingdon, UK) incorporated in the gel. Amplicon lengths were determined using Kodak Digital Science 1D software (version 2.0).
4.4. Simple Sequence Repeat Analysis
To evaluate the genetic variation in 89 accessions of T. compactum, we selected 21 primers developed in wheat by Röder et al. [53]. The commercial common wheat variety Escacena was included in the analysis. The PCR amplification (final volume: 10 µL) consisted of 20 ng of genomic DNA, 2 mM of MgCl_2_, 0.8 mM of dNTP, 5× polymerase buffer, 0.125 µM of forward primer, 0.25 µM of reverse primer, 0.25 µM of fluorochromes (FAM or 6-HEX) and 1 U of Taq polymerase (Promega). The forward primers were synthesized with a 19 bp long M13 tail (5′-CACGACGTTGTAAAACGAC-3′). The cycling protocol was: 1 min at 94 °C, 35 cycles at 94 °C for 1 min, annealing at 53 °C for 1 min and polymerization at 72 °C for 1.5 min, followed by a final extension at 72 °C for 10 min. The amplification products were separated using an automated capillary sequencer (ABI 3130 Genetic Analyzer, Applied Biosystems/HITACHI, Madrid, Spain) in the Genomics Unit of the Central Research Support Service at the University of Córdoba. The size of the amplified bands was determined based on an internal DNA standard (400HDROX) with GeneScan software (version 3.x), and the results were interpreted using Genotyper software (Version 3.7), both from Applied Biosystems. The number of different alleles and polymorphic information content (PIC) were also determined [54]. The Dice coefficient was calculated as a measure of similarity between accessions. Cluster analysis was performed using the unweighted pair group method with arithmetic means, a hierarchic agglomerative method equivalent to the average linkage between groups method. A dendrogram was constructed using the Numerical Taxonomy and Multivariate Analysis System software (NTSYS Version 2.0, Applied Biostatistics, Setauket, NJ, USA).
4.5. Statistical Analysis
To evaluate the statistical significance of differences in heading date between different allele combinations for VRN1 and PPD-D1 loci in the club wheat collection, an analysis of the variance was conducted, and the means were compared with the least significant difference (LSD) test (p < 0.05) using the Statistix software package, version 8.0. The local common wheat cultivar Escacena was included as a control.
5. Conclusions
The knowledge of diversity in major phenology genes may be useful in T. compactum breeding programs as a source of agronomically valuable traits. Furthermore, the club wheat germplasm evaluated in this study could provide new genetic resources needed for developing wheat varieties adapted to new agroecological conditions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Filatenko A.A. Hammer K. Wheat landraces from Oman: A botanical analysis Emir. J. Food Agric.20142611913610.9755/ejfa.v 26i 2.16760 · doi ↗
- 2Gul A. Allan R.E. Relation of the club gene with yield and yield components of near-isogenic wheat lines Crop Sci.19721229730110.2135/cropsci 1972.0011183 X 001200030013 x · doi ↗
- 3Johnson E.B. Nalam V.J. Zemetra R.S. Riera-Lizarau O. Mapping the compactum locus in wheat (Triticum aestivum L.) and its relationship to other spike morphology genes of the Triticeae Euphytica 200816319320110.1007/s 10681-007-9628-7 · doi ↗
- 4Kajla A. Schoen A. Paulson C. Yadav I.S. Neelam K. Riera-Lizarazu O. Leonard J. Gill B.S. Venglat P. Datla R. Physical mapping of the wheat genes in low-recombination regions: Radiation hybrid mapping of the C-locus Theor. Appl Genet.20232113615910.1007/s 00122-023-04403-037344686 · doi ↗ · pubmed ↗
- 5Wen M. Su J. Jiao C. Zhang X. Xu T. Wang T. Liu X. Wang Z. Sun L. Yuan C. Pleiotropic Effect of the compactum gene and its combined effects with other loci for spike and grain-related traits in wheat Plants 202211183710.3390/plants 1114183735890471 PMC 9316965 · doi ↗ · pubmed ↗
- 6Rayfuse L.M. Jones S.S. Variation at Glu-1 loci in club wheats Plant Breed.1993111899810.1111/j.1439-0523.1993.tb 00613.x · doi ↗
- 7Jernigan K.L. Morris C.F. Zemetra R. Chen J. Garland-Campbell K. Carter A.H. Genetic analysis of soft white wheat end-use quality traits in a club by common wheat cross J. Cereal Sci.20177614815610.1016/j.jcs.2017.06.005 · doi ↗
- 8Xu L.L. Li W. Wei Y.M. Zheng Y.L. Genetic diversity of HMW glutenin subunits in diploid, tetraploid and hexaploid Triticum species Genet. Resour. Crop Evol.20095637739110.1007/s 10722-008-9373-3 · doi ↗