Influence of Sesamia nonagrioides and Fusarium verticillioides on carbon elemental and isotopic composition in maize stem piths
Elisa Fernández-Descalzo, Lorena Álvarez-Iglesias, Ana Butrón, Serafín J. González-Prieto

TL;DR
This study examines how a stem borer insect and a fungus affect carbon composition in maize, revealing insights into plant defense mechanisms.
Contribution
The study is the first to explore the combined impact of Sesamia nonagrioides and Fusarium verticillioides on carbon metabolism in maize genotypes.
Findings
Insect infestation reduces δ¹³C by 0.6‰, partially offset by prior fungal infection.
Fungal infection alone reduces δ¹³C by 0.3‰ in maize.
Carbon isotopic signatures provide insights into maize defense mechanisms.
Abstract
The stem borer Sesamia nonagrioides and the fungus Fusarium verticillioides are frequently present in maize. However, their interaction with this crop and their effects on its physiology remain poorly understood. This study explores the combined impact of these two organisms on carbon composition and δ¹³C content in the stem pith. Eight genetically distinct maize lines (A239, A509, A630, A637, EP42, EP77, EP125, PB130) were subjected to four treatments: an untreated control (UC), infection by F. verticillioides (FV), attack by S. nonagrioides (SN), and infection by F. verticillioides and infestation by S. nonagrioides (FS). Results show moderate variation in δ¹³C values between UC genotypes (-12.6 to -13.1‰, with the exception of line EP77 at -13.8‰), with minor differences related to their inbred resistance to the insect and fungus. Insect infestation results in a noticeable reduction…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
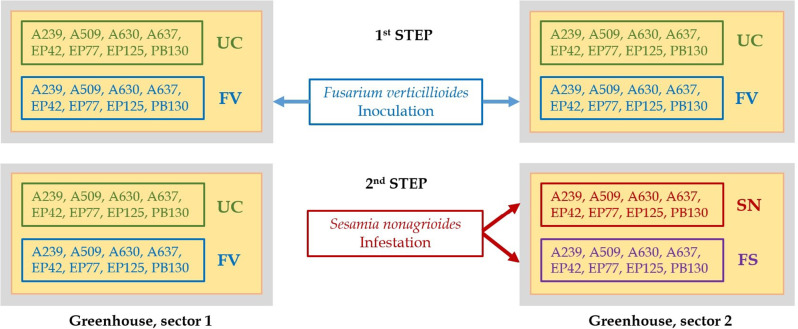 Figure 1
Figure 1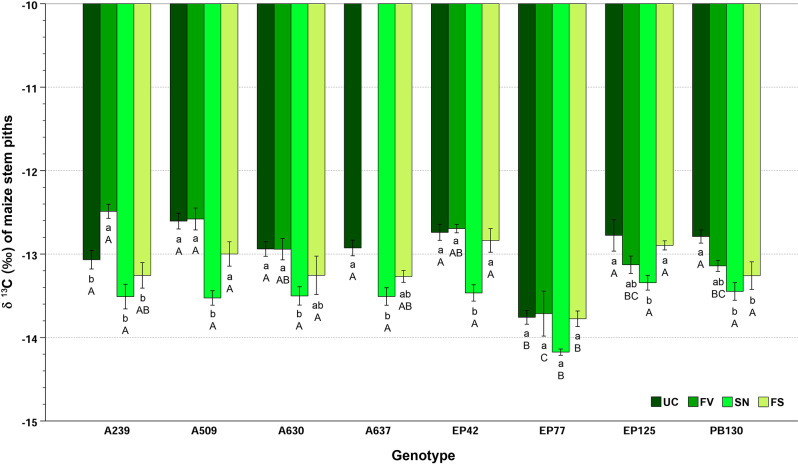 Figure 2
Figure 2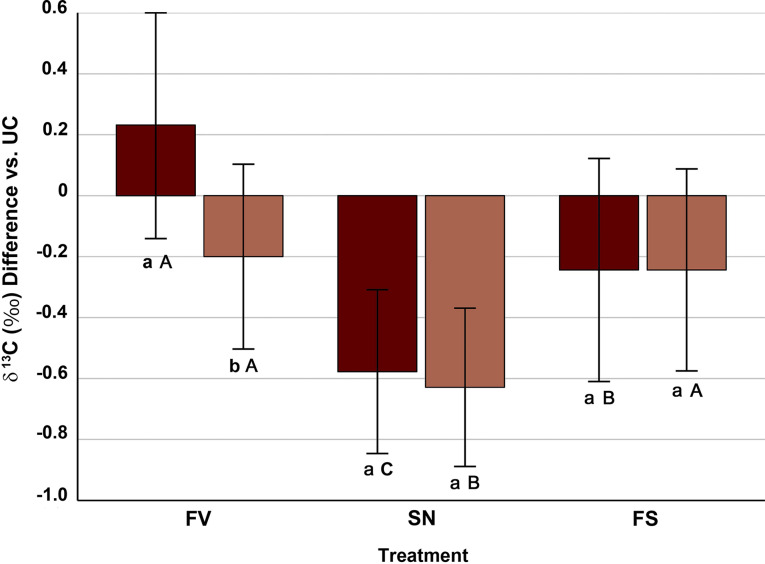 Figure 3
Figure 3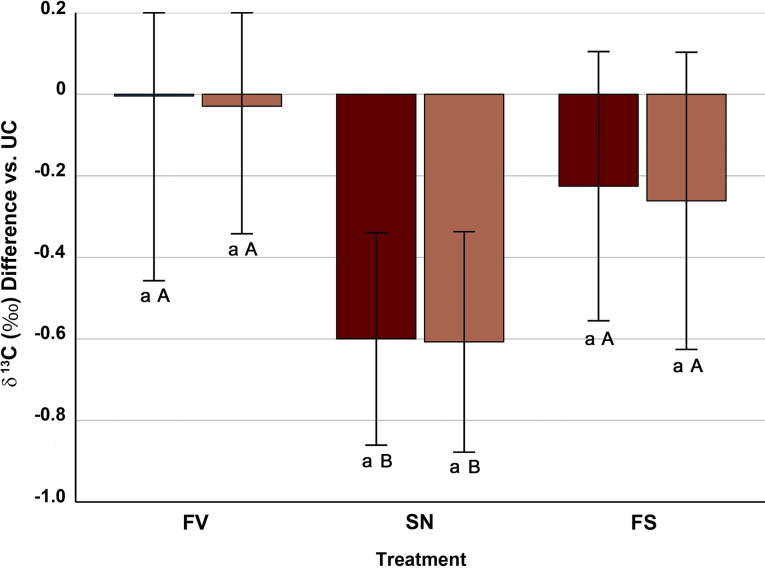 Figure 4
Figure 4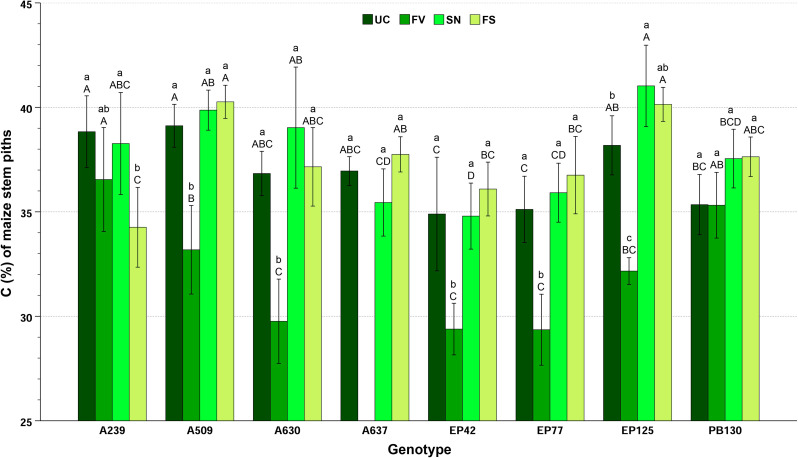 Figure 5
Figure 5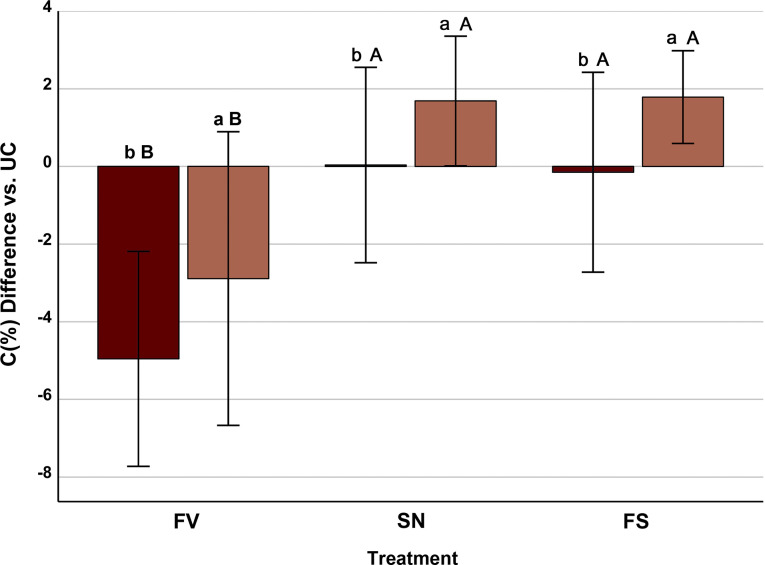 Figure 6
Figure 6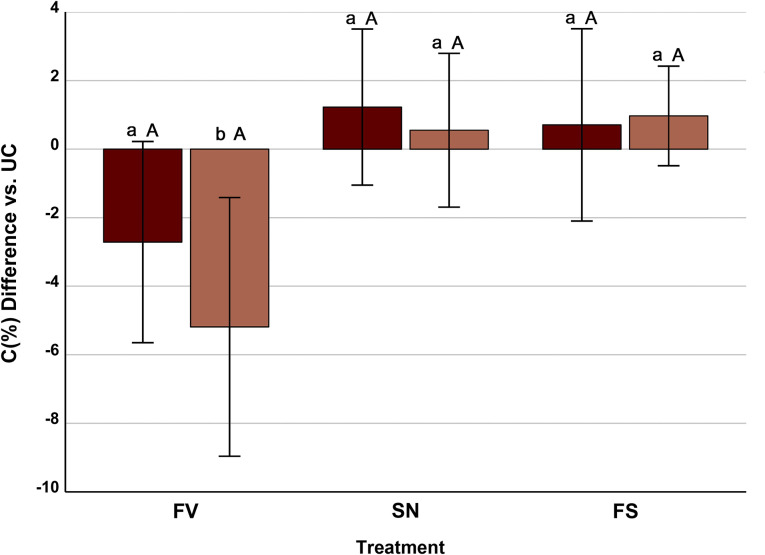 Figure 7
Figure 7- —Consejo Superior de Investigaciones Cientificas (CSIC)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCassava research and cyanide · Isotope Analysis in Ecology · Food composition and properties
Introduction
With its production growing three times faster than that of wheat or rice in the 2000–2022 period, maize (Zea mays L.) is currently the second most produced crop worldwide after sugarcane [1].
Lepidoptera caterpillars that feed on maize stalk (‘stem borers’) are one of the most damaging insect pests of the crop [2]; among them, several Sesamia species are of particular concern around the world: S. calamistis Hampson, 1910, in Africa [3], S. inferens (Walker, 1856) in Asia [4] and S. nonagrioides (Lefèbvre, 1827) -thereafter Sesamia- in the Mediterranean area [5].
The fungus Fusarium verticillioides (Sacc.) Niremberg, 1976 -thereafter Fusarium- is often found inside maize kernels in temperate areas around the world [6] and poses a threat not only to crop yield, but also to animal and human health [7]. However, this fungus is also able of asymptomatic colonization as an endophyte and it can enhance host fitness at seedling stage by promoting growth and inducing defence-associated responses such as lignin deposition, and, at mature stage, it may also contribute to increased biomass [8–10].
The maize response to Sesamia could be mediated by the presence of Fusarium as seed inoculation with the fungus upon infestation with the insect significantly altered defence metabolism in resistant inbreds [11]. In sugarcane, Mahlanza et al. [12] showed that infection by Fusarium species, especially in lower internodes, affected resistance to a borer pest. Fusarium could also be entomopathogenic against Sesamia as it has been proved for other fungal endophytes [13]. In maize, the entomopathogenic performance of Fusarium has been studied with contradictory results: while Fusarium infection of the roots impacted negatively on western corn rootworm [14], higher pest densities were found in the stem and ear of field grown maize in Fusarium infected plants [15].
Carbon isotopes are considered a powerful tool for assessing time-integrated plant responses to environmental conditions. From the two naturally occurring carbon isotopes ^12^C and ^13^C, plants are able to discriminate against the heavier ^13^C isotope during gas exchange, as Rubisco preferentially fixes ^12^CO_2_ [16]. This isotope discrimination known as Δ ^13^C results in a lower ^13^C concentration in plants compared to the atmosphere. The carbon isotope signature (or composition) of a plant tissue (δ ^13^C) is a consequence of this discrimination during CO_2_ fixation. Theory has shown that δ ^13^C is strongly related to C_i_/C_a_, the leaf intercellular to atmospheric [CO_2_] ratio, as this ratio depends on both CO_2_ supply to the leaf intercellular spaces through stomata and the CO_2_ demand driven by the photosynthetic capacity of mesophyll. Thus, δ ^13^C is linked to A/E, the ratio of photosynthesis (A) to transpiration (E), and serves as an indicator of gas exchange traits such as carbon fixation, stomatal conductance, and transpiration [17, 18].
Both Δ ^13^C and δ ^13^C vary in response to a wide range of environmental factors, including abiotic and biotic stress. Additionally, variations exist among plant genotypes and between photosynthetic pathways [19]. Although both C3 and C4 plants are depleted in ^13^C compared to atmospheric CO_2_, their average δ ^13^C values range from − 20 to -35‰ for C3 plants and from − 11 to -15‰ for C4 plants [18]. In C3 plants, variations in Δ ^13^C and δ ^13^C are consistently linked to environmental conditions. Under drought, these traits have been used as indirect selection criteria for breeding water use efficient crop varieties [20, 21].
There are few studies on how biotic stresses affect plant δ ^13^C [22, 23]. Most available studies focus on boring insects, which induce isotopic enrichment in their hosts plants. This effect is likely due to water transport disruption, a process that mimics water stress [22, 23]. However, no studies have investigated the effects of Sesamia, or other stem borer pests, on δ ^13^C in maize. Similarly, no published reports were found on the impact of Fusarium infection, alone or in combination with these pests.
This study aimed to evaluate changes in maize composition (%C, δ ^13^C) induced by Sesamia infestation and to assess potential synergistic or antagonistic effects of simultaneous Fusarium colonization. Eight genotypes with contrasting responses to either biotic agent were selected for this purpose. The following hypotheses were tested: (a) Sesamia infestation will induce maize δ ^13^C changes similar to those caused by water stress; (b) variations in plant δ ^13^C will differ among inbreds and may be partially explained by their level of resistance to Sesamia and/or Fusarium; and (c) Fusarium seed infection may influence maize plants responses to subsequent infestation.
Materials and methods
Carbon analysis equipment and standards
Elemental and isotopic analyses of carbon were carried out using an elemental analyser (EA, Carlo Erba CNS 1508, Milano, Italy) coupled to an isotope ratio mass spectrometer (IRMS, Finnigan Mat, delta C, Bremen, Germany). Measurement accuracy was ensured through the use of elemental (LECO 502–694, 10.8 ± 0.26% C, Saint Joseph, MI, USA) and isotopic standards (IAEA-CH-6, sucrose, -10.4 ± 0.2‰; IAEA-CH-7, polyethylene, -31.8 ± 0.2‰; IAEA Vienna). Samples were weighted using a Sartorius MC5 micro-balance (Göttingen, Germany, accuracy ± 1 µg).
Experimental protocol
Eight maize inbreds, either resistant (R+) or susceptible (R-) to Fusarium ear rot and Sesamia (thereafter Fusarium^(R+)^, Fusarium^(R−)^, Sesamia^(R+)^ and Sesamia^(R−^) were selected (Table 1).
Table 1. Inbreds lines used and their resistance levels to Sesamia nonagrioides and Fusarium verticillioides: R+, resistant; R-, susceptible [24–26]InbredsSesamia nonagrioides infestationFusarium verticillioides infectionPB130R+R-EP77R+R+A509R+R+EP125R+R-EP42R-R-A637R-R+A630R-R+A239R-R-
A total of 80 seeds per inbred were disinfected both internally and externally. Half of them were inoculated with Fusarium before being sown in 16 pots (2–3 seeds/pot), while the other half (non-inoculated) were sown in another 16 pots. The 32 pots were placed in a greenhouse. Once the plants were established, only one per pot was kept. For infestation, 8 inoculated and 8 non-inoculated plants per genotype were moved to an isolated greenhouse section and exposed to two Sesamia larvae per plant, generating four treatments: UC (untreated control, no Fusarium, no Sesamia), FV (Fusarium-inoculated), SN (Sesamia-infested), and FS (Fusarium-inoculated and *Sesamia-*infested) (Fig. 1).
Fig. 1. Schematic of the two-steps experimental protocol applied to the eight genotypes studied (A239, A509, A630, A637, EP42, EP77, EP125 and PB130) for obtaining the four planned treatments: UC, untreated control, no Fusarium, no Sesamia; FV, infected with Fusarium verticillioides; SN, infested with Sesamia nonagrioides; FS, infected with F. verticillioides and infested with S. nonagrioides
Initially, six replicates per inbred-treatment were used, but due to germination issues only five replicates of SN and FS were available for inbreds A509, EP42 and A239. Inbred A637 showed poor germination in Fusarium-inoculated seeds, leading to the exclusion of FV treatment for this inbred. One month later, at the reproductive stage (with well-developed and lignified tissues), stem pith samples were collected from the first internode below the cob and lyophilized. Plants showing Sesamia damage were excluded from the control group, while non-damaged plants were excluded from the infested treatments (SN and FS).
Elemental and isotopic analyses of maize
Maize subsamples (1.4 to 1.6 mg) were weighed into tin capsules (4 mm Ø x 6 mm), and analysed for their elemental (%C) and isotopic (natural abundance, δ ^13^C) composition. Isotopic composition was calculated using the following formula:
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${\delta ^{13}}C\, = \,\left( {{{{{({{{}^{13}C} \over {{}^{12}C}})}_{sample}}} \over {{{({{{}^{13}C} \over {{}^{12}C}})}_{PDB}}}} - 1} \right)\, \times \,{1000^{}}$$\end{document}Isotopic analysis followed several methodological principles, including some recommended by TD Jardine and RA Cunjak [27]. To ensure homogeneity of signal amplitudes between samples and standards, we adjusted their mass. Similarly, the signal from the internal reference injected at each analysis was harmonized by adjusting the flow rate from a CO_2_ pressure bottle calibrated against IAEA standards. Analytical accuracy and precision were verified using standards inserted after each series of samples. When wear on the EA combustion and reduction furnaces caused a deviation in accuracy of 0.3–0.4‰, they were replaced. If necessary, the isotopic values of samples were corrected according to the deviation between the measured and certified values of the IAEA standards used as reference in the analytical sequence. Finally, plants whose δ ^13^C deviated by more than 0.5‰ from the average of their respective treatments were subjected to a new analysis.
Data analysis
An exploratory analysis of %C and δ ^13^C values was first carried out to identify any outliers or anomalies likely to affect the results. The normality of the data distribution was checked using Shapiro-Wilk’s W test, and the homogeneity of variance between groups was assessed by Levene’s test. If these assumptions were not met, the data were transformed using Tukey’s power scale. The data were then analysed using three rounds of two-way ANOVA, with treatment as the first factor, and either genotype, Sesamia resistance, or Fusarium resistance as the second factor (for %C and δ ^13^C). Significant differences between the group means were determined using the Bonferroni test for multiple comparisons, with a significance level set at p < 0.05. The proportion of the variance explained by each factor or interaction was estimated using the partial eta-squared coefficient (η_p_^2^). All statistical analyses were performed using the IBM-SPSS 28.0 software.
Results
13C isotopic signature of stem piths
Two-way ANOVA revealed a significant influence of genotype (F = 21.63; p < 0.001), treatment (F = 45.30; p < 0.001) and their interaction (F = 1.95; p = 0.013) on the ^13^C isotopic signature in maize stem pith. These factors explained 51.8%, 49.1% and 21.7% of the total variance, respectively. Regarding the genotype effect, A509 had the highest (less negative) mean δ ^13^C value at -12.93‰, significantly different from most other lines, with the exception of EP42, EP125, and A239. Conversely, EP77 displayed the most negative value (-13.86‰), significantly lower than the other genotypes, while the remaining lines presented intermediate values without marked differences except for EP42, A637, and EP77. Regarding the treatment effect, the δ ^13^C isotopic signature followed a decreasing trend in the following order: FV ≈ UC (-12.95‰) > FS (-13.20‰) > SN (-13.56‰). The effects of the inbred and treatment factors were qualified by their significant interaction. In all treatments, EP77 displayed the most negative δ ^13^C with significant differences compared to the other lines, that did not differ among them except in a few pairwise comparisons for the FV treatment (A239 ≈ A509 > PB130 ≈ EP125; uppercase letters in Fig. 2). Specifically, A239 showed the lowest value in SN condition, but stood out for a significantly higher isotopic signature in FV condition than in the other treatments (Fig. 2). For the other genotypes, the lowest δ ^13^C value was recorded under SN condition, but the magnitude of significant differences with other treatments varied according to the line: A509 and EP42 (SN < UC, FV, FS), A630 (SN < UC, FV), EP125 (SN < UC, FS), A637 and PB130 (SN < UC), while for EP77 no significant differences were observed (Fig. 2).
Fig. 2. Mean ± standard error of δ ^13^C in maize stem piths for each genotype-treatment combination. Lowercase letters show differences (p < 0.05) between treatments within the same genotype; uppercase letters show differences (p < 0.05) between inbreds within the same treatment. Key of treatments: UC (untreated control, no Fusarium verticillioides, no Sesamia nonagrioides), FV (Fusarium-inoculated), SN (Sesamia-infested), and FS (Fusarium-inoculated and *Sesamia-*infested)
The two-way ANOVA with resistance to Sesamia and treatment as main sources of variation showed that both factors (F = 8.40; p = 0.004 and F = 41.41; p < 0.001, respectively) and its interaction (F = 5.85; p < 0.001) affect significantly the differences in ^13^C isotopic signature of maize stem pith of FV, SN and FS treatments with regard to the double control UC (6.3%, 39.7% and 8.5% of the variance explained, respectively), SN treatment leading to a more intense decrease of δ ^13^C than FS and FV (Fig. 3).
Fig. 3. Mean ± standard error of the differences in δ ^13^C of maize stem piths between the infection/infestation treatments (FV, Fusarium verticillioides-inoculated; SN, *Sesamia nonagrioides-*infested; FS, Fusarium-inoculated and *Sesamia-*infested) and the untreated control (UC, no Fusarium, no Sesamia). Within a given treatment, different lowercase letters show differences (p < 0.05) between inbreds resistant (light boxes) and susceptible (dark boxes) to S. nonagrioides. Within an inbred group, different uppercase letters show differences (p < 0.05) between treatments
The two-way ANOVA with resistance to Fusarium and treatment as main sources of variation showed that only the latter (F = 32.68; p < 0.001) had a significant effect on ^13^C isotopic response to Fusarium, Sesamia and the double treatment FS (p < 0.001; 34.2% of the variance explained). Compared with UC, the FV treatment did not modify consistently the δ ^13^C, whereas the FS and, especially, the SN treatments led to a decrease of δ ^13^C, which was similar in Fusarium^(R−)^ and Fusarium^(R+)^ (Fig. 4).
Fig. 4. Mean ± standard error of the differences in δ ^13^C of maize stem piths between the infection/infestation treatments (FV, Fusarium verticillioides-inoculated; SN, *Sesamia nonagrioides-*infested; FS, Fusarium-inoculated and *Sesamia-*infested) and the untreated control (UC, no Fusarium, no Sesamia). Within a given treatment, different lowercase letters show differences (p < 0.05) between inbreds resistant (light boxes) and susceptible (dark boxes) to F. verticillioides. Within an inbred group, different uppercase letters show differences (p < 0.05) between treatments
C content of stem piths
The C content of maize stem pith was significantly influenced by the inbred (F = 18.78; p < 0.001) and treatment factors (F = 89.14; p < 0.001) and their interaction (F = 6.02; p < 0.001) that explained 47.9%, 65.2% and 45.7% of the total variance, respectively. EP125, A509 and A239 presented the highest C contents (37–39%) and EP42 and EP77 the lowest (34–35%), but the significant differences among inbreds were scarce: EP125 > A637, PB130, A630, EP77, EP42; A509 > A630, EP77, EP42; A239 > EP77, EP42; and EP42 < than the rest, except EP77. Treatment FV (around 32% C) significantly differed from the other treatments (37–38% C). Although the inbred x treatment interaction was significant, the effect of the treatment was similar for all inbreds, except for A239 (Fig. 5).
Fig. 5. Mean ± standard error of C content (%) in maize stem piths for each genotype-treatment combination. Lowercase letters show differences (p < 0.05) between treatments within the same genotype; uppercase letters show differences (p < 0.05) between inbreds within the same treatment. Key of treatments: UC (untreated control, no Fusarium verticillioides, no Sesamia nonagrioides), FV (Fusarium-inoculated), SN (Sesamia-infested), and FS (Fusarium-inoculated and *Sesamia-*infested)
The two-way ANOVA considering resistance to Sesamia and treatment as main sources of variation showed that both factors (F = 14.74; p < 0.001; and F = 37.80; p < 0.001) affect the responses in C content of FV, SN and FS treatments (10.5 and 37.5% of the variance explained, respectively). In all treatments (FV, SN and FS), the C content was higher for Sesamia^(R+)^ than for Sesamia^(R−)^ inbreds (Fig. 6).
Fig. 6. Mean ± standard error of the differences in C content (%) of maize stem piths between the infection/infestation treatments (FV, Fusarium verticillioides-inoculated; SN, *Sesamia nonagrioides-*infested; FS, Fusarium-inoculated and *Sesamia-*infested) and the untreated control (UC, no Fusarium, no Sesamia). Within a given treatment, different lowercase letters show differences (p < 0.05) between inbreds resistant (light boxes) and susceptible (dark boxes) to S. nonagrioides. Within an inbred group, different uppercase letters show differences (p < 0.05) between treatments
The two-way ANOVA with resistance to Fusarium and treatment showed that only the interaction between both factors (F = 4.05; p = 0.022; 10.8% of the variance explained) affect the differences among responses in C content to the FV, SN and FS treatments, the only significant difference among responses being that between susceptible and resistant genotypes to FV treatment (Fig. 7).
Fig. 7. Mean ± standard error of the differences in C content (%) of maize stem piths between the infection/infestation treatments (FV, Fusarium verticillioides-inoculated; SN, *Sesamia nonagrioides-*infested; FS, Fusarium-inoculated and *Sesamia-*infested) and the untreated control (UC, no Fusarium, no Sesamia). Within a given treatment, different lowercase letters show differences (p < 0.05) between inbreds resistant (light boxes) and susceptible (dark boxes) to F. verticillioides. Within an inbred group, different uppercase letters show differences (p < 0.05) between treatments
Discussion
It must be highlighted that the SN treatment always had the most negative δ ^13^C values, being significant the differences in 6 out 8 genotypes irrespectively of their resistance to Sesamia. Compared to the mean δ ^13^C of UC plants (-12.95‰), SN plants showed a decrease around − 0.6‰. This result is opposed to expectations of decreased ∆ ^13^C and increased δ ^13^C based on results obtained in C3 plants attacked by borer insects [22, 23]. However, one of the main consequences of maize stem boring by Sesamia is the water stress caused by larval galleries [28]. The active tunnelling behaviour of corn borers feeding on the stalk pith also damages the surrounding vascular tissues. This results in a partially disrupted transport of water and essential nutrients throughout the plant, causing drought-like symptoms in adult maize plants. Because of that, maize δ ^13^C changes caused by Sesamia attack could be similar to those caused by drought stress. Thus, we would expect an increase in Δ ^13^C and a decrease in δ ^13^C, as in the few reports on the relationship between drought stress and isotopic discrimination in maize [21, 29, 30]. These results, opposed to the relationships found in C3 plants, are attributed to the particularities of the CO_2_ fixation process in C_4_ plants like maize [21].
Under seed inoculation with Fusarium alone (no infestation), the δ ^13^C increased in Sesamia^(R−)^ inbreds and decreased in Sesamia^(R+)^, with the resistant inbreds EP77, EP125 and PB130 showing the lowest levels for δ ^13^C. Maize infection by Fusarium has been associated to increased or decreased stomatal conductance, and discrepancies among studies could be attributed to the existence of genotype-dependent responses [31, 32]. Therefore, we hypothesize that the less negative δ ^13^C of most Sesamia^(R−)^ inbreds under systemic colonization with Fusarium could be associated to enhanced tolerance to xylem disruption mediated by lowering stomatal conductance; meanwhile, other inbreds would increase stomatal conductance.
Seed infection by Fusarium significantly reduced the percentage of carbon in the stem pith and there are at least two possible explanations, not mutually exclusive: (a) the fungal biomass itself because the C/N ratio of Fusarium sp. is almost five times lower (28 ± 7 vs. 146 ± 1) than that of maize stalks [33, 34]; and (b) decreased maize cell wall lignification, as the C content of lignin is lower than that of cellulose [35]. However, although lignin is ^13^C depleted compared to other plant constituents [36, 37], the δ ^13^C in in stem piths of FV and UC plants were similar. Besides, no significant difference was found between the Klason lignin content in the stem pith of plants developed from Fusarium-inoculated and Fusarium-free seeds [11]. Therefore, the possible role of reduction of cell wall lignification on the observed carbon changes should be disregarded.
Under infestation with Sesamia, the presence of Fusarium in the seed reduced δ ^13^C half compared to reduction caused by infestation alone. This result suggests that seed infection by Fusarium would interfere in the photosynthesis response of maize to damage by stem borers and could, simultaneously, interfere in the defensive response [38]. Nitrogen content depletion and C/N enrichment are indicators of induced defence against insect attack [39–43]. Agreeing with the expectations, after Sesamia infestation these indicators varied less in plants primed by seed inoculation than in non-inoculated ones [11]. Carbon content increments in response to Sesamia treatments (SN and FS) were always higher in resistant than susceptible genotypes. This result could be consequence of C-expensive defence enrichment in some resistant inbreds, such as A509 and EP125 that presented the highest levels for C content after both treatments [44].
As shown before, resistant inbreds presented the highest levels for C content after infestation (A509 and EP125) and/or the lowest δ ^13^C after FV treatment (EP77, EP125 and PB130). Therefore, neither single parameter could be used as proxy of resistance to Sesamia because each one would be related to a different defence mechanism and the inbreds used in the current study did not share the same mechanism of defence [45, 46].
Conclusions
Maize stem infestation by Sesamia led to a substantial stem pith δ ^13^C decrease of 0.6‰, causing drought-like symptoms. This reduction was partially counterbalanced in plants primed by Fusarium infection, in which the defensive response seemed diminished compared to Fusarium-free-seed plants. Unfortunately, neither the C content nor the ^13^C isotopic fingerprint could be used as proxy of resistance to Sesamia in breeding programs. Nevertheless, the ^13^C isotopic and C content signatures provide relevant information about maize defence mechanisms.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Malvar R, Butrón A, Ordás B, Santiago R. Causes of natural resistance to stem borers in maize. In: Crop Protection Research Advances. Edited by Burton EN, Williams PV. New York, USA: Nova Science Publishers, Inc. 2008. 57–100.
- 2Santiago R, Cao A, Malvar RA, Butrón A. Genomics of maize resistance to Fusarium ear rot and Fumonisin contamination. Toxins (Basel) 2020;12(7).10.3390/toxins 12070431 PMC 740499532629954 · doi ↗ · pubmed ↗
- 3Tiru Z, Mandal P, Chakraborty AP, Pal A, Sadhukhan S. Fusarium disease of maize and its management through sustainable approach. In: Fusarium - An overview of the Genus. Edited by Mirmajlessi SM. London, UK: Intech Open. 2021.
- 4Mc Kellar RC, Wolfe AP, Muehlenbachs K, Tappert R, Engel MS, Cheng T, Sánchez-Azofeifa GA. Insect outbreaks produce distinctive carbon isotope signatures in defensive resins and fossiliferous ambers. Proceedings of the Royal Society B: Biological Sciences. 2011;278(1722):3219–3224.10.1098/rspb.2011.0276 PMC 316902921429925 · doi ↗ · pubmed ↗
- 5Rodriguez VM, Padilla G, Malvar RA, Kallenbach M, Santiago R, Butrón A. Maize stem response to long-term attack by Sesamia nonagrioides. Front Plant Sci. 2018;9.10.3389/fpls.2018.00522 PMC 592596929740463 · doi ↗ · pubmed ↗
- 6Woźniak M, Ratajczak I, Wojcieszak D, Waśkiewicz A, Szentner K, Przybył J, Borysiak S, Goliński P. Chemical and structural characterization of maize Stover fractions in aspect of its possible applications. Mater (Basel). 2021;14(6).10.3390/ma 14061527 PMC 800400833804750 · doi ↗ · pubmed ↗
- 7Fotelli MN, Lyrou FG, Avtzis DN, Maurer D, Rennenberg H, Spyroglou G, Polle A, Radoglou K. Effective defense of Aleppo pine against the giant scale Marchalina Hellenica through ecophysiological and metabolic changes. Front Plant Sci. 2020;11.10.3389/fpls.2020.581693 PMC 775841033362812 · doi ↗ · pubmed ↗