Genetic diversity and phylogeny of avian orthoreovirus isolates from Peruvian broiler chickens
Doris Villanueva-Pérez, Luis Tataje-Lavanda, Angela Montalván-Avalos, Gisela Isasi-Rivas, Diego Paredes-Inofuente, Suly Montoya-Ortiz, Manolo Fernández-Sánchez, Eliana Icochea, Manolo Fernández-Díaz

TL;DR
This study examines the genetic diversity of avian orthoreovirus in Peruvian broiler chickens and identifies three main genotypes.
Contribution
The study provides new insights into the genetic classification of avian orthoreovirus isolates in Peru.
Findings
Avian orthoreovirus isolates were grouped into genotypes 1, 2, and 3 based on σC gene analysis.
The findings highlight the need for further research to improve vaccination strategies against the virus.
Abstract
Avian orthoreovirus infections are increasingly common in Peruvian broiler chickens, primarily causing arthritis/tenosynovitis syndrome. In this study, sequencing and phylogenetic analysis of the σC gene revealed that avian orthoreovirus isolates were clustered in genotypes 1, 2, and 3. However, further research is needed to understand its genetic diversity and improve vaccination strategies.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
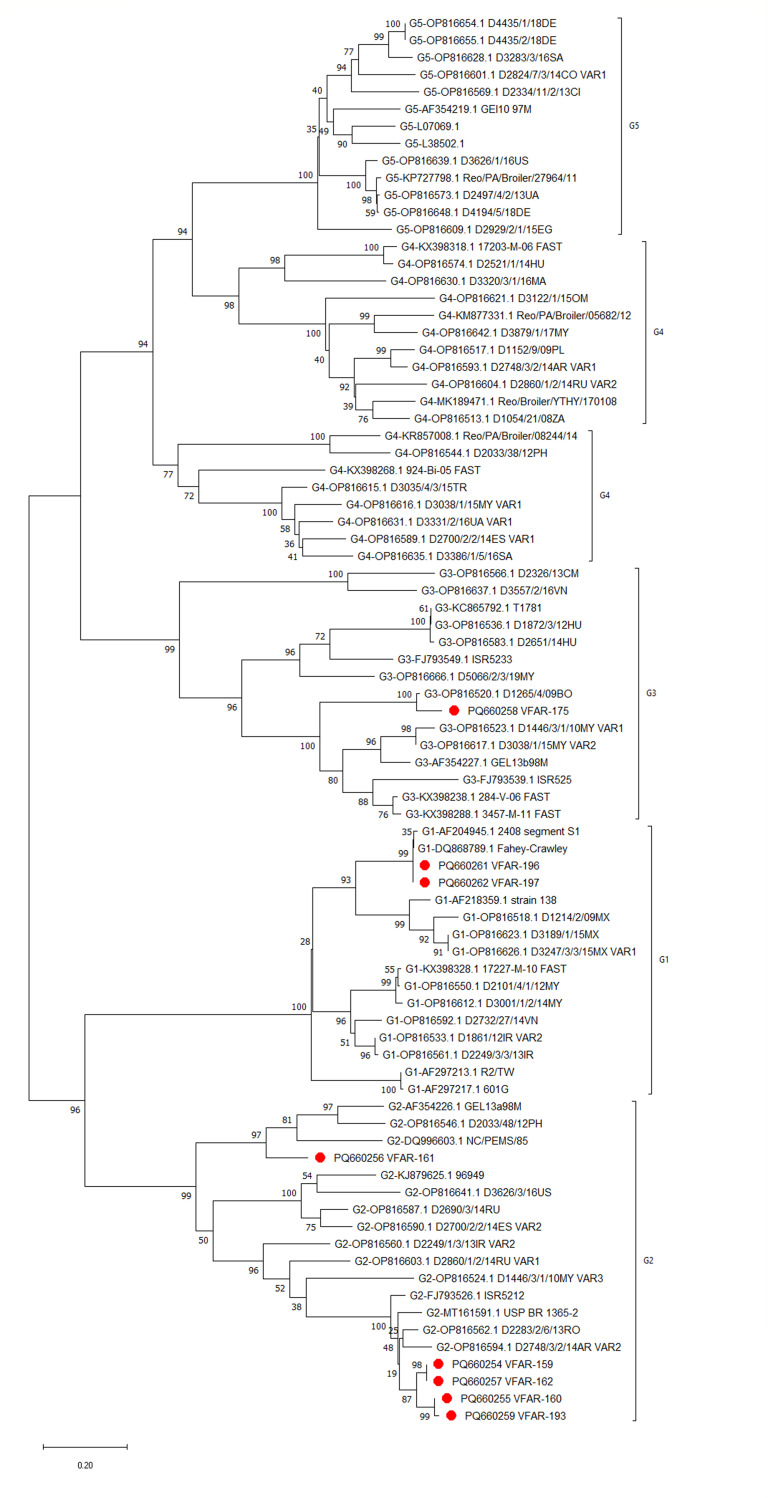 Fig 1
Fig 1| Isolate name (accession number/SRA) | Age of host (days) | Origin | Genotypic cluster | Reference sequence | Consensus length (pb) | σC gene length/% GC | BLASTn coverage/ identity (%) | Closest isolate (accession number/country) |
|---|---|---|---|---|---|---|---|---|
| VFAR-196 ( | 28 | Arequipa | 1 |
| 1101 | 981/48 | 100/99.55 | Reo/ZC0219/SD/2017 ( |
| VFAR-197 ( | 35 | La Libertad | 1 |
| 1101 | 981/48 | 100/99.46 | Reo/ZC0219/SD/2017 ( |
| VFAR-159 ( | 8 | La Libertad | 2 |
| 963 | 963/48.1 | 99/94.9 | USP_BR_362-5 ( |
| VFAR-160 ( | 14 | La Libertad | 2 |
| 963 | 963/48.1 | 99/95.94 | USP_BR_362-5 ( |
| VFAR-161 ( | 14 | La Libertad | 2 |
| 1089 | 981/47.4 | 99/84.23 | USP_BR_362-5 ( |
| VFAR-162 ( | 10 | Arequipa | 2 |
| 963 | 963/48.1 | 99/94.9 | USP_BR_362-5 ( |
| VFAR-193 ( | 21 | Arequipa | 2 |
| 963 | 963/48.2 | 99/95.73 | USP_BR_362-5 ( |
| VFAR-175 ( | 21 | Arequipa | 3 |
| 768 | 768/46 | 100/92.71 | D7 ( |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsViral gastroenteritis research and epidemiology · Animal Virus Infections Studies · Herpesvirus Infections and Treatments
ANNOUNCEMENT
Avian orthoreovirus or avian reovirus (ARV) causes viral arthritis/tenosynovitis in commercial poultry, affecting productivity. ARV belongs to the Orthoreovirus genus, subfamily Spinareovirinae of the Reoviridae family and has a genome of 10 double-stranded RNA segments (1–3). The S1 segment encodes the σC gene, crucial for viral attachment, immune response (3, 4), and genotypic classification (2, 5, 6). To date, ARV genotypes in Peru remain unidentified.
This study sequenced and analyzed the σC gene of ARV isolates from Peruvian broilers. Eight tendon samples were collected in March 2024 from broilers with viral arthritis symptoms in poultry-producing regions of Peru (Table 1). ARV detection was conducted using real-time RT-PCR targeting the S3 gen (7). The PCR-positive samples were minced, homogenized, and centrifuged. The supernatant was filtered and used to infect LMH cells. After 5 days, cells showing >90% cytopathic effects were collected and clarified, and supernatants were conserved at −80°C (4). RNA was extracted from 140 µL supernatant using the QIAamp Viral RNA Mini Kit (Qiagen, Germany), followed by reverse transcription with ProtoScript II Reverse Transcriptase (New England Biolabs, USA) and random hexamers according to the manufacturer’s protocol. Subsequently, the σC gene was amplified using primers P1 and P4 (2) and Q5 High-Fidelity 2× Master Mix (New England Biolabs, USA). PCR products (_~_1,088 bp) were purified using the QIAquick Gel Extraction Kit (Qiagen, Germany), quantified, and prepared for sequencing using the ONT Rapid Barcoding Kit 24 (SQK-RBK114.24).
Sequencing was performed on a MinION Mk1B device (Oxford Nanopore Technologies, UK) using an R10.1 flow cell (N50: 858). Basecalling and demultiplexing were conducted using Dorado Software v7.4.12 (8) with Fast model v4.3.0, 400 bps, generating 270.21 k reads and 155.9 Mb of high-quality bases. Initially, processed reads were aligned to reference sequences representing diverse ARV genotypes using the GalaxyTrakr platform (https://galaxytrakr.org/) (9); all tools were run with default parameters unless otherwise specified. Quality control was assessed using tools such as FastQC (10) (Galaxy Version 0.73+galaxy0), Porechop (11) (Galaxy Version 0.2.4+galaxy0), NanoFilt (12) (Galaxy Version 0.1.0), and MultiQC (13) (Galaxy Version 1.11+galaxy1). Consensus sequences were generated from each alignment and subsequently compared against ARV references (GenBank: AF330703.1, MT161583.1, MN879660.1, ON782394.1, OP816520.1) (Table 1) using Map with BWA-MEM (14) (Galaxy Version 0.7.17.2), Samtools depth (15) (Galaxy Version 1.15.1+galaxy0), and Consensus Sequence (16) (Galaxy Version 0.7.0+galaxy1). The closest matching isolate sequences were identified via BLASTn (17, 18) (Table 1). This approach improved consensus sequence accuracy by minimizing SNPs, indels, and frameshift errors, ensuring a more precise genotype representation. Phylogenetic analysis was performed on eight σC gene sequences using MEGA 11 v 11.0.13 (19). Multiple sequence alignments of nucleotides were generated with MUSCLE (20), and a neighbor-joining phylogenetic tree was constructed with 1,000 bootstrap replicates. Evolutionary distances were computed using the Tajima-Nei method with gamma distribution modeling (shape parameter = 1). The analysis involved 82 nucleotide sequences and 499 aligned positions in the final data set.
The results showed consensus sequence lengths of these σC genes range from 768 bp to 981 bp (Table 1). Phylogenetic analysis revealed three distinct genotypes (1, 2, and 3) circulating among Peruvian broilers (Fig. 1). Genotype 2, closely related to Brazilian isolates, was the most prevalent among ARV isolates, suggesting epidemiological links. The diversity highlights the need for continued surveillance to guide vaccine strategies.
Phylogenetic tree of the σC gene sequences of avian orthoreovirus isolates from Peruvian broiler chickens.
Phylogenetic analysis revealed five distinct clades, with the eight Peruvian ARV isolates clustering into genotypes 1, 2, and 3. Isolates were highlighted with red circles for visual distinction. The tree was constructed using the neighbor-joining method with 1,000 bootstrap replicates, and branch lengths represent evolutionary distances calculated using the Tajima-Nei method. A gamma distribution (shape parameter = 1) was applied to model rate variation among sites. The data set included 82 sequences, with 499 positions after gap and missing data elimination. Bootstrap values are indicated at branch nodes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Jones RC. 2000. Avian reovirus infections. Rev Sci Tech OIE 19:614–625. doi:10.20506/rst.19.2.123710935283 · doi ↗ · pubmed ↗
- 2Kant A, Balk F, Born L, van Roozelaar D, Heijmans J, Gielkens A, ter Huurne A. 2003. Classification of Dutch and German avian reoviruses by sequencing the σC protein. Vet Res 34:203–212. doi:10.1051/vetres:200206712657212 · doi ↗ · pubmed ↗
- 3Goldenberg D, Pasmanik-Chor M, Pirak M, Kass N, Lublin A, Yeheskel A, Heller D, Pitcovski J. 2010. Genetic and antigenic characterization of sigma C protein from avian reovirus. Avian Pathol 39:189–199. doi:10.1080/03079457.2010.48096920544425 · doi ↗ · pubmed ↗
- 4Ayalew LE, Gupta A, Fricke J, Ahmed KA, Popowich S, Lockerbie B, Tikoo SK, Ojkic D, Gomis S. 2017. Phenotypic, genotypic and antigenic characterization of emerging avian reoviruses isolated from clinical cases of arthritis in broilers in Saskatchewan, Canada. Sci Rep 7:3565. doi:10.1038/s 41598-017-02743-828620186 PMC 5472580 · doi ↗ · pubmed ↗
- 5Palomino-Tapia V, Mitevski D, Inglis T, van der Meer F, Abdul-Careem MF. 2018. Molecular characterization of emerging avian reovirus variants isolated from viral arthritis cases in Western Canada 2012-2017 based on partial sigma (σ)C gene. Virology (Auckl) 522:138–146. doi:10.1016/j.virol.2018.06.00630029013 · doi ↗ · pubmed ↗
- 6Sellers HS. 2023. Avian reoviruses from clinical cases of tenosynovitis: an overview of diagnostic approaches and 10-year review of isolations and genetic characterization. Avian Dis 66. doi:10.1637/aviandiseases-D-22-9999036715473 · doi ↗ · pubmed ↗
- 7Sprygin AV, Andreychuk DB, Kolotilov AN, Volkov MS, Runina IA, Mudrak NS, Borisov AV, Irza VN, Drygin VV, Perevozchikova NA. 2010. Development of a duplex real-time Taq Man PCR assay with an internal control for the detection of Mycoplasma gallisepticum and Mycoplasma synoviae in clinical samples from commercial and backyard poultry. Avian Pathol 39:99–109. doi:10.1080/0307945100360462120390544 · doi ↗ · pubmed ↗
- 8Oxford Nanopore Technologies. 2025. Dorado. Available from: https://github.com/nanoporetech/dorado. Retrieved 31 Dec 2024.