Complete genome sequence of Pseudomonas sp. PP3, a dehalogenase-producing bacterium, confirms the unusual mobile genetic element DEH
Gordon Webster, Amy J. Baldwin, Edward Cunningham-Oakes, Alex J. Mullins, Rachel Dodds, Katja E. Hill, Li Ling Lee, Mark J. Leggett, Andrew W. Topping, Andrew J. Weightman

TL;DR
The genome of Pseudomonas sp. PP3, a bacterium that breaks down harmful chemicals, was fully sequenced and shows it is closely related to P. reinekei.
Contribution
The paper confirms the presence of the unusual mobile genetic element DEH in the genome of Pseudomonas sp. PP3.
Findings
The genome of Pseudomonas sp. PP3 is 6.42 Mb in size.
The genome is most closely related to P. reinekei.
The bacterium produces dehalogenase, which is important for bioremediation.
Abstract
Pseudomonas sp. PP3, originally isolated from contaminated soil enriched in a chemostat culture on 2,2-dichloropropionic acid, has a 6.42 Mb genome, most closely related to P. reinekei. This well-characterized organism continues to provide key insights into adaptive dehalogenase-mediated bioremediation of halogenated organic pollutants.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
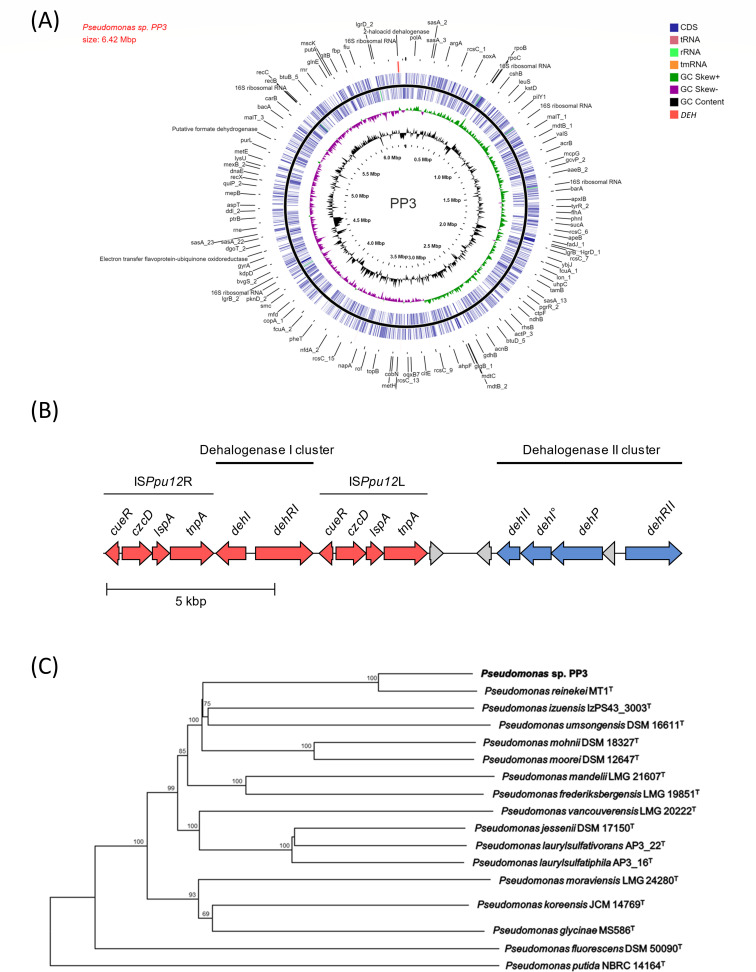 Fig 1
Fig 1| Genome | Genome size (Mbp) | G + C content (%) | ||
|---|---|---|---|---|
| Pairwise ANI (%) | Pairwise DDH (%) | |||
| 6.42 | 59.2 | 100 | 100 | |
| 6.25 | 59.2 | 95.6 | 63.3 | |
| 6.86 | 59.6 | 88.5 | 33.6 | |
| 6.70 | 59.7 | 88.2 | 33.0 | |
| 6.59 | 59.6 | 88.3 | 33.5 | |
| 6.55 | 59.7 | 88.5 | 34.0 | |
| 6.16 | 62.3 | 84.3 | 22.4 | |
| 6.39 | 60.2 | 85.6 | 24.7 | |
- —Biotechnology and Biological Sciences Research Councilhttp://dx.doi.org/10.13039/501100000268
- —UK Research and Innovationhttp://dx.doi.org/10.13039/100014013
- —Biotechnology and Biological Sciences Research Councilhttp://dx.doi.org/10.13039/501100000268
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial bioremediation and biosurfactants · Microbial Community Ecology and Physiology · Enzyme Production and Characterization
ANNOUNCEMENT
Halo-organic compounds are used as herbicides, pesticides, preservatives, solvents, and other applications. The persistence and toxicity of many of these compounds raise serious environmental concerns regarding their use (1). Biodegradation of such compounds, including haloalkanoic acids, by Pseudomonas species is well known (2).
Pseudomonas sp. strain PP3 was isolated from a soil microbial community in a chemostat culture on the herbicide 2,2-dichloropropionic acid as the sole carbon and energy source as described (3–5). For long-term storage, the strain was kept in 40% (vol/vol) glycerol stocks at −80°C and for routine laboratory culture grown aerobically in standard basal salts medium [SBS; (4)] containing 5 mM halogenated substrate at 30°C, shaking at 150 rpm.
For genome sequencing, PP3 was grown in SBS with 10 mM 2-chloropropionic acid for 24 h. Cells were pelleted by centrifugation (4,000 rpm, 10 min, ALC-PK120 centrifuge), and genomic DNA was extracted using the Wizard genomic DNA purification kit (Promega) according to the manufacturer’s protocol. Following QC, DNA was sheared and size-selected (~10 kb) using Covaris g-TUBE and sequenced by Novogene (UK). Sequencing libraries were prepared with the SMRTbell template prep kit 1.0 (PacBio) and NEBNext DNA library prep kit for Illumina. Genome sequencing was performed on a PacBio Sequel SMRT Cell 1M and an Illumina NovaSeq 6000 (paired-end, 2 × 150 bp). In total, 208,470 high-quality PacBio subreads (average length = 5,218 bp; N50 = 7,983 bp) and 19,253,220 Illumina raw reads were obtained and quality checked with FastQC v0.11.8. The genome (620× coverage) was assembled de novo from the PacBio subreads into one contig using Flye v2.8 (6) and polished with both the PacBio reads using Arrow (via pbmm2 v1.4.0, GCpp v2.0.0 tools; https://github.com/PacificBiosciences), and Illumina reads with Pilon v1.23 (7) using default settings. The assembled genome was reoriented with Circlator v1.5.5 (8) at the dnaA gene start position.
Genome size and other metrics for the assembly are as follows: 6,421,237 bp and 59.17%G + C, similar to other Pseudomonas species (Table 1; 9), 5,745 coding DNA sequences (CDS), 19 rRNAs, 71 tRNAs, and four ncRNAs identified using PGAP v6.7 (10) and CGView (11) (Fig. 1A). The PP3 genome contains two 2-haloalkanoate dehalogenase genes (representing dehI and dehII gene families) previously described (12), and one silent dehI gene. The DEH mobile genetic element in PP3 (13) is confirmed to contain a dehI family gene (12) and its regulatory gene, dehRI, flanked by two almost identical insertion sequences (ISPpu12). The DEH element is located close to a separate putative dehalogenase operon (Fig. 1B). Several other ORFs encoding putative dehalogenases and enzymes associated with halo-organic catabolism are also evident in the genome.
Visual representation and phylogenetic classification of the Pseudomonas sp. PP3 genome and DEH mobile element. (A) Pseudomonas sp. PP3 chromosome map of 5,744 predicted proteins. From outer circle to the center: select view of gene identification; dehalogenase gene clusters (within and around DEH); all CDSs (blue) and RNAs (crimson/green/orange) genes on forward strand; all CDSs (blue) and RNAs (crimson/green/orange) genes on reverse strand; GC skew (positive GC skew values are plotted in green, and negative values are in purple); GC content (black); scale bar. The map was generated using the Circular Genome Viewer (CGView). (B) Close-up of the dehalogenase gene clusters (dehI) and the upstream dehII region highlighted in (A), generated using Clinker v0.0.27 (14). The gene designations for the dehalogenase I cluster within the DEH element are as follows: dehI, dehalogenase I family gene (12); dehRI, σ54-dependent activator; tnpA, putative ISL3-family transposase; lspA, putative lipoprotein signal peptidase; czcD, putative heavy-metal-associated efflux transporter; cueR, putative heavy-metal-associated responsive transcriptional regulator. The positions of the ISPpu12 insertion sequence (independently mobile) flanking regions are also indicated (13). The gene designations for the dehalogenase II cluster are as follows: dehII, dehalogenase II family gene; dehI°, cryptic dehalogenase I family gene; dehP, putative permease transporter; dehRII, σ54-dependent activator. (C) Phylogenetic classification of Pseudomonas sp. PP3. The genomes of the 14 closest related type strains along with type strains P. putida and P. fluorescens were used for phylogenetic analysis as described by TYGS (15). The tree was inferred with FastME 2.1.6.1 (16) using Genome BLAST Distance Phylogeny (GBDP) distances calculated from genome sequences. The branch lengths are scaled in terms of GBDP distance formula d5. The numbers above branches are GBDP pseudo-bootstrap support values > 60% from 100 replications, with an average branch support of 86.1%. The tree was rooted at the midpoint.
Average nucleotide identity (ANI) analysis with pyani v0.2.10 (18) and genome comparisons using the Type (Strain) Genome Server [TYGS; (15)] inferred that PP3 groups phylogenomically with P. reinekei MT1^T^ (Table 1; Fig. 1C), with an ANI value >95% proposed as the same species (17). However, dDDH analysis produced a value below the 70% species threshold (17) compared with P. reinekei and other Pseudomonas genomes (Table 1). The ANI and dDDH values together suggest that PP3 may represent a new species closely related to P. reinekei.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chaudhry GR, Chapalamadugu S. 1991. Biodegradation of halogenated organic compounds. Microbiol Rev 55:59–79. doi:10.1128/mr.55.1.59-79.19912030673 PMC 372801 · doi ↗ · pubmed ↗
- 2Pieper DH, Reineke W. 2004. Degradation of chloroaromatics by Pseudomona(d)s, p 509–574. In Ramos JL (ed), Pseudomonas. Vol. 3. Biosynthesis of macromolecules and molecular metabolism. Springer US, Boston, MA.
- 3Senior E, Bull AT, Slater JH. 1976. Enzyme evolution in a microbial community growing on the herbicide Dalapon. Nature 263:476–479. doi:10.1038/263476 a 0972691 · doi ↗ · pubmed ↗
- 4Slater JH, Lovatt D, Weightman AJ, Senior E, Bull AT. 1979. The growth of Pseudomonas putida on chlorinated aliphatic acids and its dehalogenase activity. J Gen Microbiol 114:125–136. doi:10.1099/00221287-114-1-125 · doi ↗
- 5Weightman AJ, Slater JH, Bull AT. 1979. The partial purification of two dehalogenases from Pseudomonas putida PP 3. FEMS Microbiol Lett 6:231–234. doi:10.1016/0378-1097(79)90067-3 · doi ↗
- 6Kolmogorov M, Yuan J, Lin Y, Pevzner PA. 2019. Assembly of long, error-prone reads using repeat graphs. Nat Biotechnol 37:540–546. doi:10.1038/s 41587-019-0072-830936562 · doi ↗ · pubmed ↗
- 7Walker BJ, Abeel T, Shea T, Priest M, Abouelliel A, Sakthikumar S, Cuomo CA, Zeng Q, Wortman J, Young SK, Earl AM. 2014. Pilon: an integrated tool for comprehensive microbial variant detection and genome assembly improvement. P Lo S One 9:e 112963. doi:10.1371/journal.pone.011296325409509 PMC 4237348 · doi ↗ · pubmed ↗
- 8Hunt M, Silva ND, Otto TD, Parkhill J, Keane JA, Harris SR. 2015. Circlator: automated circularization of genome assemblies using long sequencing reads. Genome Biol 16:294. doi:10.1186/s 13059-015-0849-026714481 PMC 4699355 · doi ↗ · pubmed ↗