In vitro models to study viral-induced asthma exacerbation: a short review for a key issue
Rémi Pereira De Oliveira, Clément Droillard, Gilles Devouassoux, Manuel Rosa-Calatrava

TL;DR
This review discusses in vitro models for studying how viruses worsen asthma, highlighting their strengths and limitations.
Contribution
The paper provides a focused review on in vitro models for viral-induced asthma exacerbation, emphasizing their suitability for specific aspects of the disease.
Findings
Asthma involves complex interactions between multiple cell types and inflammatory pathways.
No single in vitro model can fully represent the entire asthma physiopathology.
Different in vitro models are useful for studying specific aspects of virus-related asthma.
Abstract
Asthma is a heterogenous inflammatory bronchial disease involving complex mechanisms, several inflammatory pathways, and multiples cell-type networks. Bronchial inflammation associated to asthma is consecutive to multiple aggressions on epithelium, such as microbiologic, pollutant, and antigenic agents, which are responsible for both T2 and non-T2 inflammatory responses and further airway remodeling. Because asthma physiopathology involves multiple crosstalk between several cell types from different origins (epithelial, mesenchymal, and immune cells) and numerous cellular effectors, no single and/or representative in vitro model is suitable to study the overall of this disease. In this short review, we present and discuss the advantages and limitations of different in vitro models to decipher different aspects of virus-related asthma physiopathology and exacerbation.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
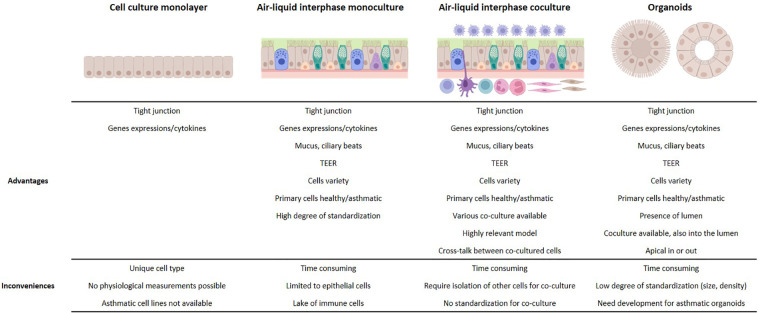 Figure 1
Figure 1| Cell lines | Can be infected by | References |
|---|---|---|
| CaLu-3 | RSV, HRV, SARS-CoV, Influenza | ( |
| 16HBE | RSV, HRV, SARS-CoV-2, Influenza, HMPV | ( |
| PBEC | RSV, HRV, SARS-CoV-2, Influenza, HMPV | ( |
| 3D co-cultures models | Cell origins | Conclusions | References |
|---|---|---|---|
| PBEC and fibroblast | Healthy donor cells | Activation of smooth muscle through TGF- | ( |
| BEAS-2B or PBEC and airway smooth muscle cells | Healthy donor for smooth muscle cells | Proliferation of smooth muscle and stimulation of pro-inflammatory genes expression | ( |
| 16HBE and HUVECs cells | Cell lines | Proliferation of endothelial cells and expression of E-selectin and ICAM-I | ( |
| PBEC and macrophages | Healthy donor cells | Inflammation and viral clearance | ( |
| PBEC and macrophages and dendritic cells | PBECs from healthy, asthmatic, COPD donors | Inflammation | ( |
| PBEC and fibroblast and eosinophils | Healthy donor cells | Epithelium remodeling and inflammation | ( |
| PBEC and PBMC/lymphocytes | Healthy donor cells | Inflammation, exacerbation and viral clearance | ( |
| PBEC and neutrophils | Healthy donor cells | Neutrophils infiltration induced by HRV infection | ( |
| Organoids and macrophages | Healthy donor cells | Increase of pro-inflammatory proteins with macrophages | ( |
| Organoids and resident T lymphocytes | Healthy donor cells | T lymphocytes activation after SARS-CoV-2 infection | ( |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAsthma and respiratory diseases · IL-33, ST2, and ILC Pathways · Immune Cell Function and Interaction
Introduction
1
Asthma affects more than 300 million individuals worldwide (1) and is characterized by airway obstruction, bronchial hyperresponsiveness, and bronchial inflammation. Asthma is schematically separated into two main immune and inflammatory-related phenotypes. The most prevalent is the T2-high phenotype, which involves allergic and/or eosinophilic inflammation, involving mainly Th2 lymphocytes, basophils, eosinophils, mast cells, and type 2 innate lymphoid cells (ILC2). By contrast, the T2-low inflammatory phenotype is driven by Th17 lymphocytes and/or neutrophils and is characterized by the absence of allergy, eosinophilic inflammation, and other T2 determinants, such as high FeNO levels, nasal polyposis, or atopic dermatitis (2). In some cases, when both neutrophil and eosinophil infiltration in the airways is low, the condition is called a mixed or pauci-granulocytic phenotype (3). A major burden of the disease is the occurrence of acute clinical events, known as asthma exacerbations, which are associated with an increase in bronchial inflammation in response to various environmental stimuli (4–6). Among these stimuli, allergens, pollutants, bacteria, and viruses are largely implicated and responsible for initiating the inflammatory cascade. They activate three main epithelial alarmins: thymic stromal lymphopoietin (TSLP), IL-33, and IL-25, which activate dendritic cells, ILC2, macrophages, T cells, and mast cells (7, 8). Respiratory viral infections are a leading cause of asthma exacerbation, with human rhinovirus (HRV) being the most commonly implicated, followed by respiratory syncytial virus (RSV) (9) and, to a lesser extent, human metapneumovirus (hMPV) and influenza viruses (9, 10). The virus-related mechanisms of asthma exacerbation are not yet fully understood and the development of relevant in vitro models to study virus/asthma functional relationships remains a significant challenge. In this short review, we summarize and discuss the advantages and limitations of the commonly used in vitro models for studying asthma and its exacerbation by viral infections.
Monoculture cell line models
2
The simplest models involve human bronchial cell lines cultured in medium or at the air–liquid interface (ALI). Some of these cell lines, such as CaLu-3 and 16HBE, express tight junction (TJ) proteins, which are key determinants of epithelium permeability and integrity (11, 12). Disrupted TJs are observed in the bronchial epithelium of asthmatic patients, significantly affecting respiratory airway barrier function (13). Studies using allergens, nanoparticles, or cigarette smoke extract have shown an increase in transepithelial permeability through the disruption of TJs on CaLu-3 (14, 15) and 16HBE (16, 17). Therefore, measuring TJ integrity and exploring functional interactions with respiratory viruses in permissive epithelial cells (Table 1) should be considered, especially as HRV, RSV, hMPV, influenza, and coronaviruses are known to target specific TJ proteins (18–21). However, although airway epithelial cell lines are more accessible than primary cells and can be cultured in ALI models to study host responses to viral infections (22), they are less physiologically relevant due to the lack of cell diversity (i.e., only ciliated cells are present, with no secretory or basal cells) (23, 24) and the absence of cell lines from asthmatic patients.
Primary cell-based ALI human airway models
3
Primary bronchial epithelial cells (PBECs) are more relevant models, as they are directly recovered from patient biopsies, including those with asthma or other pulmonary diseases such as chronic obstructive pulmonary disease (COPD). PBECs can be cultured in submerged mono-cultures or in an air–liquid interface as a pseudostratified human airway epithelium (HAE) that includes several types of differentiated cells (ciliated, club, goblet, and basal) (37). These models allow for the investigation of a broader range of functional and physiological characteristics, including morphology, transepithelial electrical resistance, tight junction integrity, mucus secretion, cilia clearance, innate immune response (38, 39), sensitivity to allergens or respiratory viruses (Table 1), and transcriptomic signatures (40). As such, PBECs are widely used to study asthma physiopathology and exacerbation (41, 42). For example, the T2 associated IL-13 cytokine has been extensively studied for its ability to modulate gene expression profiles (43) and alter mucin production in primary cell-based ALI models (44), which are features of asthma physiopathology.
Moreover, HAE models are well-suited for investigating asthma exacerbation related to viral infection, as asthmatic patients exhibit physiological impairments that can be measured in HAE. These impairments include alterations in tight junctions and epithelium integrity (20, 21, 45), modifications in mucus production, mucociliary function (46–49), and aberrant epithelial tissue repair and remodeling through epithelial-mesenchymal transition (50–52).
Furthermore, PBECs can also react to viral infections through the innate immune response, which is an important factor in asthma exacerbation (8, 53, 54), as respiratory viruses are known to suppress antiviral responses and enhance inflammation in epithelial cells (55–57). For example, Wark et al. demonstrated an impairment of interferon beta (IFN-β) in ALI cultures from asthmatic patient PBECs compared to healthy controls during HRV infection (36). Consistently, the addition of IFN-β restored the expression of antiviral genes against HRV in asthmatic PBECs, thereby reducing the duration of inflammation (58). In addition, viral double-stranded RNA induces a stronger expression of the alarmin TSLP, an important enhancer of asthma exacerbation through the induction of the T2 inflammatory pathway, in asthmatic PBEC-based ALI compared to healthy controls (59). These results are similar to those obtained with HAE models infected by different respiratory viral infections, such as RSV (60), HRV (61), and SARS-CoV-2 (62).
ALI models co-cultivated with primary cells of different origins
4
Asthma is a complex lung disease that involves not only epithelial cells but also immune cells implicated in inflammation, viral clearance, and remodeling processes. In addition, mesenchymal cells, such as fibroblasts, endothelial cells, and smooth muscle cells, contribute to tissue inflammation and physiopathology (63). In this context, co-cultured primary cells of different origins can be very useful for investigating cellular crosstalk and their respective contributions to asthma development and exacerbation (Table 2). The ability of respiratory viruses to impair host immune responses, thereby increasing the risk of asthma exacerbation (64–67), underscores the need for further investigation into the functional relationships between epithelial, mesenchymal, and immune cells within integrated in vitro asthma models. In this regard, Transwell® cell separation system used for ALI cultures (68) allows for the co-cultivation of epithelial cells in the open air with other types of primary cells, whether adherent or not, on the lower surface of the Transwell®. Furthermore, co-culture models with direct contact between primary cells can be highly beneficial for mimicking in vivo crosstalk.
Co-culture of ALI airway models with mesenchymal cells
4.1
ALI co-culture models of healthy airway epithelium with fibroblast cells adhering to the lower surface of Transwell® have highlighted that chemical injury to the epithelium leads to the secretion of TGF-β2 by fibroblasts, which in turn triggers the expression of several asthma markers, such as Tenascin-C and α-smooth muscle actin (α-SMA) (69), as observed in the bronchus of asthmatic patients (81). In a similar model, Reeves et al. reported that fibroblasts secrete more hyaluronan when co-cultured with asthmatic ALI airway epithelium compared to healthy tissue, thereby promoting inflammation by increasing leukocyte binding. In this study, bronchial epithelial cells were co-cultured with human lung fibroblasts with Fibroblast Growth Media (FGM-2) and PneumaCult ALI for 96 h (82). Consistently, fibroblasts infected by RSV also secreted more hyaluronan, promoting mast cell binding and protease production, which are both known to contribute to asthma inflammation (83).
Co-culture models of epithelial cells with healthy smooth muscle cells in various culture media have been developed to study cell proliferation and pro-inflammatory responses (70), in line with their role in asthma physiopathology through bronchoconstriction and airway tissue remodeling (84). In addition, the monocyte migration chemokine CCL5 was reported to be significantly more expressed in asthmatic smooth muscle cells than in healthy ones when the ALI model was infected by HRV. This aligns with the increased risk of asthma exacerbation observed through monocyte migration in infected asthma patients (85). On the other hand, co-culturing asthmatic smooth muscle cells with bronchial epithelial cells increases their susceptibility to HRV infection compared to co-cultures with healthy smooth muscle cells, with an increased expression of CCL20 by smooth muscle cells altering the antiviral PKR/eIF2 pathway within epithelial cells (86).
Endothelial cells are also important for tissue inflammation by binding factors on the surface membrane that enhance leukocyte infiltration (87). A co-culture model involving epithelial 16HBE and endothelial HUVECs, stimulated with virus mimicking double-stranded RNA, showed an increased expression of leukocyte-recruiting adhesion molecules (E-selectin and ICAM-I) on endothelial cells. In this study, Blume et al. co-cultured human bronchial epithelial cells in minimum essential medium (MEM) with Glutamax and serum, and HUVEC cells in M199 medium supplemented with L-Glutamine seeded at the basal side, for up to 8 days (71). Similar results were obtained when healthy ALI cultures were infected with SARS-CoV-2 (88).
Finally, ALI co-culture models are not only suitable for studying crosstalk between epithelial and mesenchymal cells involved in bronchial inflammation and immune cell recruitment, but they are also essential for characterizing the epithelium’s sensibility to viral infections. This makes them highly interesting for deciphering virus-related mechanisms that lead to asthma exacerbation.
Co-culture of ALI airway models and immune cells
4.2
ALI airway models co-cultured with additional immune cell components are even more suitable for studying the mechanisms underlying asthma physiopathology. After initial epithelial cell injuries, immunes cells are recruited, activated, and amplify bronchial inflammatory cascades that can lead to asthma exacerbation and tissue remodeling. In this context, M1- or M2-type polarized macrophages contribute to determining the asthma phenotype (89) and exacerbation in cases where viral clearance is delayed (90).
Using Transwell® co-culture, Ronaghan et al. co-cultured human bronchial epithelial cells in ALI with macrophages in PneumaCult ALI medium or ImmunoCult-SF Macrophage Medium (IMM) for 72 h (72). They reported that M1 macrophage polarization reduced RSV infection in healthy PBECs ALI cultures, but not M2 polarization (72). These results are consistent with the predominance of M2 macrophages in asthmatic patients (91), which are associated with decreased in vivo interferon secretion and the promotion of T2 cytokines. This could help explain why asthmatics are more susceptible to RSV infection and related asthma exacerbation (92).
Other studies have developed more complex models by co-cultivating three different cell types. Paplinska-Goryca et al. used PBECs from healthy, asthmatic, or COPD patients in co-culture with macrophages on apical side and dendritic cells in the lower surface of Transwell®. This model highlighted the importance of immune cells and the origin of PBECs in the expression of alarmins when epithelium was exposed to double-stranded RNA stimulation (93). Similarly, Choe et al. showed the importance of eosinophils in airway epithelium remodeling and inflammation through a co-culture model of epithelial and fibroblast cells with eosinophils. Briefly, eosinophils, isolated from human venous blood, were seeded on top of the epithelium with RPMI 1640 medium. After 2 h of incubation and medium removal, eosinophils were stimulated with calcium ionophore for 48 h of co-culture (74). This co-culture model provides information about lung epithelium remodeling mechanisms in asthmatics (51).
Co-culturing peripheral blood mononuclear cells (PBMCs) at the lower surface of the Transwell® allows for the inclusion of a broader range of immune cells, enabling the study of crosstalk between epithelial cells and lymphocytes or natural killer (NK) cells. Several studies have focused on the T1/T2 balance and inflammatory cytokines, such as IP-10 and IFN-γ, during influenza and HRV infections of healthy nasal epithelium co-cultivated with PBMC in RPMI supplemented with serum for 48 h (75, 76). Qin et al. reported that RSV infection of healthy bronchial ALI cultures promotes an increase in Th2 cell populations from co-cultivated PBMC (77). Although NK cell polarization is known to be important for viral clearance and inflammation control through the secretion of IFN-γ or IL-4 and by killing eosinophils and neutrophils (94–96), their role in asthma physiopathology and exacerbation is not fully understood. To explore this further, co-culture models of human nasal epithelial cells with PBMC have been used to highlight NK cell activation after influenza infection (76). Future co-culture models with HAE and purified NK cells are needed to further study their role in asthma exacerbation during viral infections.
Organoids
5
Organoids are three-dimensional structures derived from stem cells or induced pluripotent stem cells, which harbor some of the structural and functional characteristics of organs, including the bronchus, to study airway development and physiopathology (97–100). These organoids are spherical, with a lumen at the sphere’s center or epithelial cells exposed at the surface (101), allowing them to be permissive to respiratory viruses like RSV (102, 103), SARS-CoV-2 (102, 104, 105), HMPV (106), influenza (104), or human parainfluenza 3 virus (107). Studies using healthy organoids have focused on aspects of asthma pathology, including goblet cell metaplasia (108) and remodeling (109).
Organoids co-cultured with immune cells are also being developed to mimic functional cell–cell interactions. Various methods have been explored: (i) seeding immune cells and lung stem cells together, (ii) co-cultivation of organoids with isolated immune cells, (iii) injecting immune cells into the organoid lumens, and (iv) expanding cultures from tissue samples that include both epithelial and resident immunes cells (110). Seo et al. co-cultivated macrophages with organoids and stimulated them with lipopolysaccharide (LPS) to model alveolar inflammation. They showed an increase in pro-inflammatory protein expression in the presence of macrophages, supporting the relevance of such organoid models (79). Organoids with resident immune cells have also been used to study SARS-CoV-2 infection and the corresponding T lymphocyte response (80), making these models promising for studying the crosstalk between resident immune cells and lung tissue. However, the development of organoid models using cells from asthmatic patients is currently lacking whereas they would be useful to complement the ALI models to further decipher viral mediated mechanisms associated with this pathology.
Lung-on-chip
6
Microfluidic systems, known as lung-on-chip models, are more complex systems that enable the movement of cells and media on the basal side, or the exposure of stimuli by injecting air onto the apical side of the airway (111, 112). For example, a lung-on-chip system with a co-culture of lung epithelium and circulating neutrophils in the basal medium was used to explore the mechanism behind asthma exacerbation induced by HRV infection. Briefly, the epithelium was cultivated in ALI on the surface of the chip membrane. On the other side, endothelial cells were cultivated in the vascular channel of the chip. After HRV inoculation at the apical side of the epithelium, neutrophils isolated from human peripheral blood were added to the chip’s reservoirs and perfused in RPMI 1640 (78). Studies have shown that neutrophil infiltration into the epithelium, a factor of neutrophilic asthma exacerbation, was induced by HRV infection and significantly increased with IL-13 stimulation (78). Another innovative system, known as “breathing lung-on-cheap,” mimics breathing and has been used to evaluate the effects of toxic particles potentially involved in the development and/or exacerbation of asthma (113). This model could be interesting when studying the impact of respiratory volume on asthmatic epithelium infection and asthma exacerbation. However, further development is needed to incorporate cells from patients.
Discussion
7
In vitro 3D airway models offer significant advantages compared to monolayer cell line-based in vitro models (Figure 1). Airway epithelial functions, including relevant inflammatory and antiviral responses, are reconstituted with a large variety of primary cells from healthy individuals of different ages and/or from patients with various bronchial diseases, such as asthma or COPD. This makes ALI and organoid models valuable and complementary tools for studying the complex mechanisms involved in asthma and its exacerbations. However, further development of and characterization of co-culture models involving mesenchymal and immune cells are needed to better understand key aspects of asthma physiopathology, such as inflammation, immune cell polarization, and tissue remodeling, leading to the development of new therapeutic approaches.
Advantages and limitations of in vitro models for the study of asthma exacerbation.
In conclusion, ALI co-culture models, organoids, and lung-on-chip systems are highly effective tools for studying asthma and its exacerbation from viral infections. With a diverse range of models available, each offering unique strengths and limitations, the choice of the most suitable in vitro model will depend on its relevance and ability to complement others in revealing various aspects of virus-induced asthma pathophysiology and exacerbation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Brusselle GG Koppelman GH. Biologic therapies for severe asthma. N Engl J Med. (2022) 386(2):157–71. 10.1056/NEJ Mra 203250635020986 · doi ↗ · pubmed ↗
- 2Habib N Pasha MA Tang DD. Current understanding of asthma pathogenesis and biomarkers. Cells. (2022) 11(17):2764. 10.3390/cells 1117276436078171 PMC 9454904 · doi ↗ · pubmed ↗
- 3Liu T Woodruff PG Zhou X. Advances in non-type 2 severe asthma: from molecular insights to novel treatment strategies. Eur Respir J. (2024) 64(2):2300826. 10.1183/13993003.00826-202338697650 PMC 11325267 · doi ↗ · pubmed ↗
- 4Aghasafari P George U Pidaparti R. A review of inflammatory mechanism in airway diseases. Inflamm Res. (2019) 68(1):59–74. 10.1007/s 00011-018-1191-230306206 · doi ↗ · pubmed ↗
- 5Calvén J Ax ERådinger M. The airway epithelium—a central player in asthma pathogenesis. IJMS. (2020) 21(23):8907. 10.3390/ijms 2123890733255348 PMC 7727704 · doi ↗ · pubmed ↗
- 6Wang Y Bai C Li K Adler KB Wang X. Role of airway epithelial cells in development of asthma and allergic rhinitis. Respir Med. (2008) 102(7):949–55. 10.1016/j.rmed.2008.01.01718339528 · doi ↗ · pubmed ↗
- 7Gauvreau GM Sehmi R Ambrose CS Griffiths JM. Thymic stromal lymphopoietin: its role and potential as a therapeutic target in asthma. Expert Opin Ther Targets. (2020) 24(8):777–92. 10.1080/14728222.2020.178324232567399 · doi ↗ · pubmed ↗
- 8Jartti TBønnelykke K Elenius V Feleszko W. Role of viruses in asthma. Semin Immunopathol. (2020) 42(1):61–74. 10.1007/s 00281-020-00781-531989228 PMC 7066101 · doi ↗ · pubmed ↗