Interactome of the oncogenic ΔNp73α isoform in human papillomavirus 38 E6/E7-transformed keratinocytes
Luc Negroni, Valerio Taverniti, Massimo Tommasino, Katia Zanier

TL;DR
This study identifies proteins that interact with ΔNp73α, an oncogenic form of p73, in HPV38-transformed skin cells linked to non-melanoma skin cancer.
Contribution
The paper presents the first interactome of ΔNp73α in HPV38 E6/E7-transformed keratinocytes.
Findings
ΔNp73α interacts with multiple proteins in both organelle and cytoplasmic compartments.
The interactome includes proteins involved in cell survival and proliferation pathways.
Findings highlight potential therapeutic targets in HPV-related skin cancer.
Abstract
ΔNp73α is a major oncogenic isoform of tumor suppressor p73. Here, we report ΔNp73α protein-binding partners in the organelle and cytoplasmic compartments of a cellular model consisting of human keratinocytes transformed by the E6 and E7 oncoproteins of the β-HPV38 virus associated with non-melanoma skin cancer.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
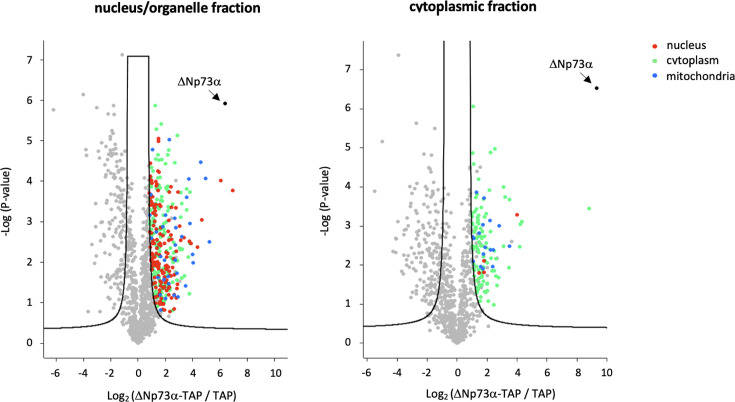 Fig 1
Fig 1| Log2 | -Log | Accession | Gene name | Log2 | -Log | Accession | Gene name |
|---|---|---|---|---|---|---|---|
| Nuclear/organelle fraction | |||||||
| 6.93 | 3.77 |
| ANXA4 | 2.42 | 1.66 |
| RCN2 |
|
|
|
|
| 2.4 | 3.87 |
| ERCC5 |
| 6.09 | 4.02 |
| TP63 | 2.38 | 2.13 |
| PAWR |
| 5.26 | 2.51 |
| ACSL1 | 2.38 | 2.35 |
| TNK1 |
| 4.97 | 4.06 |
| MRPS22 | 2.33 | 1.36 |
| PPP2CA |
| 4.66 | 3.05 |
| LUC7L3 | 2.33 | 1.68 |
| JMY |
| 4.58 | 4.48 |
| MRPS35 | 2.32 | 1.31 |
| ATP6V1H |
| 4.37 | 2.38 |
| ANXA11 | 2.32 | 0.79 |
| MED15 |
| 4.03 | 1.98 |
| MRPS31 | 2.31 | 3.74 |
| SNW1 |
| 3.99 | 2.19 |
| MRPS17 | 2.3 | 3.34 |
| FRYL |
| 3.87 | 2.46 |
| ARNT | 2.3 | 1.4 |
| DDX20 |
| 3.82 | 2.47 |
| MYO1E | 2.28 | 5.03 |
| MTHFD1L |
| 3.82 | 2.95 |
| MRPS23 | 2.28 | 1.78 |
| ELOA |
| 3.8 | 3.74 |
| SVIL | 2.28 | 2.61 |
| MACF1 |
| 3.78 | 2.51 |
| PQBP1 | 2.27 | 1.36 |
| PIK3C2A |
| 3.77 | 1.22 |
| EXOC4 | 2.26 | 4.77 |
| KRT4 |
| 3.74 | 4.06 |
| MRPS27 | 2.26 | 1.2 |
| DCAF6 |
| 3.7 | 2.14 |
| DDX60 | 2.26 | 2.54 |
| P4HA2 |
| 3.57 | 3.61 |
| MRPS9 | 2.26 | 1.46 |
| MRPS6 |
| 3.57 | 3.3 |
| PARP14 | 2.25 | 1.47 |
| WDR12 |
| 3.56 | 3.81 |
| SIPA1L1 | 2.24 | 1.47 |
| PKP2 |
| 3.5 | 1.42 |
| VPS13C | 2.24 | 3.03 |
| SF3B4 |
| 3.41 | 1.67 |
| DYNLRB1 | 2.23 | 1.65 |
| IDH3B |
| 3.35 | 2.6 |
| RPS15 | 2.22 | 1.7 |
| MMTAG2 |
| 3.27 | 2.32 |
| EIF4E2 | 2.21 | 1.88 |
| RRP1 |
| 3.22 | 3.45 |
| TMOD3 | 2.21 | 3.37 |
| OSBPL9 |
| 3.22 | 1.61 |
| SH2D4A | 2.19 | 2.76 |
| VPS51 |
| 3.21 | 2.64 |
| MRPS5 | 2.19 | 3.26 |
| TRIO |
| 3.14 | 2.31 |
| CDK18 | 2.19 | 3.77 |
| STRIP1 |
| 3.14 | 2.55 |
| RPS6KA4 | 2.19 | 2.22 |
| RPAP2 |
| 3.12 | 3.12 |
| ANXA5 | 2.18 | 4.65 |
| DNAAF5 |
| 3.12 | 3.72 |
| TRIM4 | 2.18 | 3.88 |
| HOOK2 |
| 3.06 | 2.35 |
| FYCO1 | 2.17 | 1.4 |
| EIF2B5 |
| 3.05 | 2.89 |
| TUBGCP2 | 2.16 | 1.67 |
| MAP3K5 |
| 3.04 | 1.53 |
| CCDC85C | 2.12 | 1.78 |
| LRRC41 |
| 2.95 | 1.15 |
| MRPS16 | 2.11 | 3.81 |
| CAMK1 |
| 2.93 | 3.73 |
| PPP6C | 2.1 | 1.92 |
| RPRD1B |
| 2.89 | 5.13 |
| EPPK1 | 2.1 | 2.29 |
| MYLK |
| 2.88 | 1.46 |
| GSK3B | 2.1 | 1.83 |
| PSMC4 |
| 2.87 | 2.04 |
| HSDL2 | Cytoplasmic fraction | |||
| 2.87 | 1.36 |
| MICAL3 |
|
|
|
|
| 2.85 | 1.73 |
| HADHB |
|
|
|
|
| 2.84 | 1.56 |
| TRADD |
|
|
|
|
| 2.83 | 2.27 |
| RPA3 |
|
|
|
|
| 2.8 | 1.12 |
| CPSF4 |
|
|
|
|
| 2.8 | 1.75 |
| PHKG2 |
|
|
|
|
| 2.79 | 2.86 |
| MRPS34 |
|
|
|
|
| 2.78 | 4.02 |
| KIAA1671 |
|
|
|
|
| 2.77 | 3.34 |
| CAMKK1 |
|
|
|
|
| 2.75 | 2.12 |
| TARBP1 |
|
|
|
|
| 2.74 | 2.43 |
| DAP3 |
|
|
|
|
| 2.73 | 2.05 |
| TIMM44 |
|
|
|
|
| 2.73 | 1.77 |
| ZNHIT2 |
|
|
|
|
| 2.69 | 2.26 |
| RSU1 |
|
|
|
|
| 2.69 | 3.09 |
| ALDH4A1 |
|
|
|
|
| 2.69 | 2.04 |
| ABCF2 |
|
|
|
|
| 2.69 | 4.46 |
| LGALS3BP |
|
|
|
|
| 2.69 | 4.45 |
| ALOX15B |
|
|
|
|
| 2.67 | 1.69 |
| NOC4L |
|
|
|
|
| 2.65 | 0.84 |
| NCOA3 |
|
|
|
|
| 2.64 | 1.19 |
| TUFM |
|
|
|
|
| 2.64 | 1.19 |
| MYBBP1A |
|
|
|
|
| 2.62 | 2.09 |
| DIMT1 |
|
|
|
|
| 2.61 | 2.23 |
| BAG2 |
|
|
|
|
| 2.6 | 2.5 |
| NAP1L1 |
|
|
|
|
| 2.58 | 2 |
| ATXN2 |
|
|
|
|
| 2.54 | 3.44 |
| LRCH3 |
|
|
|
|
| 2.53 | 1.51 |
| MTMR1 |
|
|
|
|
| 2.52 | 0.81 |
| PHKA2 |
|
|
|
|
| 2.51 | 1.87 |
| GTF2I |
|
|
|
|
| 2.5 | 2.08 |
| ARHGAP21 |
|
|
|
|
| 2.47 | 1.12 |
| EIF4ENIF1 |
|
|
|
|
| 2.46 | 3.36 |
| NUP133 |
|
|
|
|
| 2.46 | 2.57 |
| HERC4 | ||||
| 2.46 | 3.09 |
| RALGAPB | ||||
| 2.43 | 2.68 |
| RNMT | ||||
- —Institut National de la Santé et de la Recherche Médicalehttp://dx.doi.org/10.13039/501100001677
- —Agence Nationale de la Recherchehttp://dx.doi.org/10.13039/501100001665
- —Fondation pour la Recherche Médicalehttp://dx.doi.org/10.13039/501100002915
- —Institut National Du Cancerhttp://dx.doi.org/10.13039/501100006364
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCancer-related Molecular Pathways · Virus-based gene therapy research · interferon and immune responses
ANNOUNCEMENT
The tumor suppressor p53 family comprises three transcription factors (TFs), p53, p63, and p73, which, despite sharing high sequence identity, display specific functions and regulation. All three TFs are enriched in the nucleus, where they act on transcription and DNA repair, whereas their levels are regulated in the cytoplasm by ubiquitin-dependent or ubiquitin-independent degradation. In addition, mitochondrial localization has been reported for p53 and p73 (1). The genes coding for the three proteins (TP53, TP63, and TP73) can also be expressed as multiple isoforms (2). The highly carcinogenic ΔNp73α isoform investigated here is a dominant-negative inhibitor of full-length p53 and p73 (TAp53 and TAp73α) (3). It is generated by transcription from the internal P2 promoter of TP73 and, consequently, lacks the N-terminal transactivation domain (2, 4, 5). Expression of viral oncoproteins can also result in ΔNp73α accumulation (6, 7), including the E6 and E7 oncoproteins from cutaneous beta (β) human papillomavirus (HPV) types (8–10) associated with non-melanoma skin cancer (11).
We have performed affinity purification-mass spectrometry (AP-MS) to characterize the ΔNp73α interactome using human keratinocytes immortalized by the β-HPV38 E6 and E7 oncoproteins as a model of cellular transformation (named 38HK hereafter) (8). To this end, we generated two stable cell lines, expressing either ΔNp73α fused to the N-terminus of the tandem affinity purification (TAP) tag (ΔNp73α-TAP) or TAP alone (negative control) (12). To discriminate between cytoplasmic versus nuclear or other organelle interactants, 38HK ΔNp73α-TAP and TAP 38HK cells were fractionated as described in our previous report (12). Briefly, 38HK cells were lysed mechanically in hypotonic buffer and centrifuged. The supernatant, corresponding to the cytoplasmic extract, was recovered. The pellet, containing nuclei and other organelles, was resuspended in buffer containing NaCl (250 mM) and glycerol (20%), incubated on ice, and centrifuged. The supernatant from this second centrifugation corresponds to the nuclear/organelle extract. Then, extracts were processed by tandem affinity purification (13) following the protocol described in Reference (12). Protein complexes were eluted from calmodulin beads and analyzed by SDS-PAGE and Western blotting or silver staining. Protein levels were quantified by the Bradford assay.
Eluates from calmodulin beads were precipitated with trichloroacetic acid, and the pellet was washed with acetone and dissolved with 2 M urea. Samples were digested with 100 ng of trypsin at 37°C for 10 h. The resulting peptide mixtures were directly analyzed by MS with a nanoLC U3000 in-line coupled with an Orbitrap Elite mass spectrometer using a nano-electrospray source (Thermo Scientific). MS measurements were performed on one biological sample per condition. Technical variability was estimated from three MS acquisitions for each sample (technical replicates). Peptides were separated using a C18 column (75 µm × 25 cm) with a 35 min linear gradient from 5% to 50% buffer B (A: 0.1% trifluoroacetic acid in H_2_O/B: 80% ACN, 0.08% trifluoroacetic acid in H_2_O). The mass spectrometer was operated in data-dependent mode with survey scans from m/z 300–1650 acquired in the Orbitrap at a resolution of 120,000 at m/z 400. Proteins were identified using Proteome Discoverer 2.5 software (Thermo Scientific), with false discovery rate (FDR) < 1% and at least one unique peptide. Label-free quantification was based on eXtracted Ion Chromatography (XIC) using the Minora node from Proteome Discoverer. Precursor and fragment mass tolerance were set at 7 ppm and 0.5 Da, respectively. Oxidation (M) was set as variable modification and carbamidomethylation (C) as fixed modification. The human fasta database with 20,419 sequences was obtained from Uniprot (https://www.uniprot.org/proteomes/UP000005640). Statistical analyses were performed using Perseus 1.6.1.5 according to the processing workflow described by the authors (14). The repeated t-test used for volcano plot analyses was based on standard parameters of Perseus (FDR 5%, S0 = 1).
The lists of the top interactants of ΔNp73α (Log2FC > 2.1) in the nucleus/organelle and cytoplasmic fractions are reported in Table 1. The statistics from Perseus is presented in a volcano plot with each protein colored according to Gene Ontology Cellular Component (Fig. 1). The nuclear/organelle and cytoplasmic fractions are strongly enriched with protein partners with reported nuclear and cytoplasmic localization, respectively, thereby validating our biochemical approach. While more abundant in the nuclear/cytoplasmic fractions, mitochondrial binders are also present in the cytoplasmic fraction.
Volcano plot analysis of ΔNp73α partners in the nucleus/organelle (left) and cytoplasm (right) fractions of 38HK cells. The black continuous line indicates the limit of significance (FDR 5%, S0 = 1). Proteins situated on the right-hand side of this line (positive Log2 value) are significant interactants. These proteins are color coded according to the localization retrieved from the Gene Ontology Resource (https://geneontology.org). Red: nucleus; green: cytoplasm; blue: mitochondria; gray: localization not reported; black: ΔNp73α bait.
In our previous study (12), we have characterized the interaction of ΔNp73α with the E2F4/p130 transcriptional repressor complex, which is detected in the nuclear/organelle fraction of these analyses. Results show that E2F4 binds to ΔNp73α but not to the full-length TAp73α protein, suggesting that the interactomes of the oncogenic isoform and of the tumor suppressor protein may be different.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sayan AE, Sayan BS, Gogvadze V, Dinsdale D, Nyman U, Hansen TM, Zhivotovsky B, Cohen GM, Knight RA, Melino G. 2008. P 73 and caspase-cleaved p 73 fragments localize to mitochondria and augment TRAIL-induced apoptosis. Oncogene 27:4363–4372. doi:10.1038/onc.2008.6418362891 · doi ↗ · pubmed ↗
- 2Murray-Zmijewski F, Lane DP, Bourdon J-C. 2006. P 53/p 63/p 73 isoforms: an orchestra of isoforms to harmonise cell differentiation and response to stress. Cell Death Differ 13:962–972. doi:10.1038/sj.cdd.440191416601753 · doi ↗ · pubmed ↗
- 3Di C, Yang L, Zhang H, Ma X, Zhang X, Sun C, Li H, Xu S, An L, Li X, Bai Z. 2013. Mechanisms, function and clinical applications of D Np 73. Cell Cycle 12:1861–1867. doi:10.4161/cc.2496723708520 PMC 3735700 · doi ↗ · pubmed ↗
- 4Lee CW, La Thangue NB. 1999. Promoter specificity and stability control of the p 53-related protein p 73. Oncogene 18:4171–4181. doi:10.1038/sj.onc.120279310435630 · doi ↗ · pubmed ↗
- 5Fontemaggi G, Kela I, Amariglio N, Rechavi G, Krishnamurthy J, Strano S, Sacchi A, Givol D, Blandino G. 2002. Identification of direct p 73 target genes combining DNA microarray and chromatin immunoprecipitation analyses. J Biol Chem 277:43359–43368. doi:10.1074/jbc.M 20557320012213815 · doi ↗ · pubmed ↗
- 6Accardi R, Fathallah I, Gruffat H, Mariggiò G, Le Calvez-Kelm F, Voegele C, Bartosch B, Hernandez-Vargas H, Mc Kay J, Sylla BS, Manet E, Tommasino M. 2013. Epstein - Barr virus transforming protein LMP-1 alters B cells gene expression by promoting accumulation of the oncoprotein ΔNp 73α. P Lo S Pathog 9:e 1003186. doi:10.1371/journal.ppat.100318623516355 PMC 3597522 · doi ↗ · pubmed ↗
- 7Allart S, Martin H, Detraves C, Terrasson J, Caput D, Davrinche C. 2002. Human cytomegalovirus induces drug resistance and alteration of programmed cell death by accumulation of delta N-p 73alpha. J Biol Chem 277:29063–29068. doi:10.1074/jbc.M 20197420012034725 · doi ↗ · pubmed ↗
- 8Accardi R, Dong W, Smet A, Cui R, Hautefeuille A, Gabet A-S, Sylla BS, Gissmann L, Hainaut P, Tommasino M. 2006. Skin human papillomavirus type 38 alters p 53 functions by accumulation of delta Np 73. EMBO Rep 7:334–340. doi:10.1038/sj.embor.740061516397624 PMC 1456898 · doi ↗ · pubmed ↗