Ploidy plasticity drives fungal resistance to azoles used in agriculture and clinics
Kaustuv Sanyal, Aswathy Narayanan

TL;DR
This paper explores how changes in ploidy in Candida tropicalis lead to resistance against antifungal drugs used in agriculture and medicine.
Contribution
It reveals a novel connection between ploidy plasticity and antifungal resistance in Candida tropicalis.
Findings
Ploidy plasticity is linked to the emergence of resistance to azole antifungals.
The study focuses on Candida tropicalis as a model organism for understanding resistance mechanisms.
Abstract
The rapid growth in antimicrobial resistance is of great medical concern. A new study in PLOS Biology unveils the link between ploidy plasticity and the emergence of antifungal resistance in Candida tropicalis. The rapid growth in antimicrobial resistance is of great medical concern. This Primer highlights a new study in PLOS Biology that unveils the link between ploidy plasticity and the emergence of antifungal resistance in Candida tropicalis.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
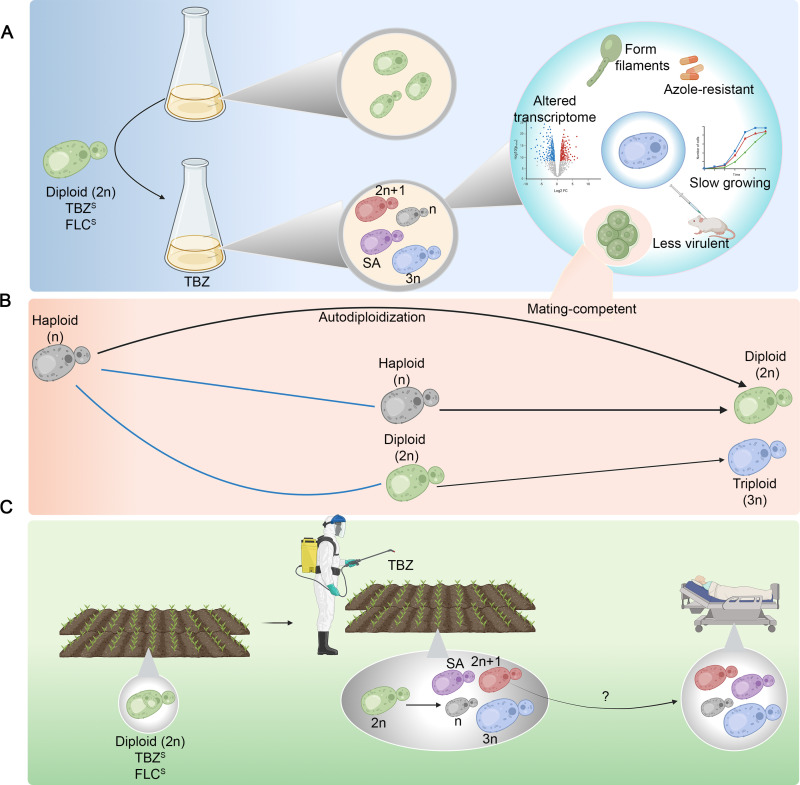 Figure 1
Figure 1- —JC Bose fellowship
- —http://dx.doi.org/10.13039/501100005116Jawaharlal Nehru Centre for Advanced Scientific Research
- —Bose Institute, West Bengal, India
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant tissue culture and regeneration · Plant Pathogens and Fungal Diseases · Nematode management and characterization studies
The fungal kingdom encompasses species with a wide variety of habitats: some thrive as free-living organisms, while others are commensals residing in host systems. A few commensals can become opportunistic pathogens once the host’s immune system is compromised. Among this vast, heterogeneous group of fungal species, there are some that are found in the environment, causing infections in humans and other animals. The impact of fungal pathogens in healthcare and economy has been overlooked for decades, but we are beginning to comprehend the potential effects of anthropogenic activities on the fungal kingdom in the environment [1]. Factors such as the rampant use of high doses of antifungals in poultry and fungicides in agricultural farmlands combined with global warming are thought to be contributing to an alarming increase in drug resistance and the emergence of new pathogens. Azoles are widely used antifungals in clinics; they are also the major component of agricultural fungicides used globally [1]. How do azole-based fungicides used in agriculture lead to the emergence of drug-resistant strains that exhibit cross-resistance to clinical azoles? A recent study in PLOS Biology by Hu and colleagues provides insights into novel mechanisms underlying the acquired resistance in Candida tropicalis.
C. tropicalis, an opportunistic fungal pathogen, belongs to the phylum Ascomycota. C. tropicalis is also isolated from environmental niches—its natural habitats include environmental reservoirs like forest soil, agricultural fields and water bodies [2]. However, it is also classified under high-priority fungal pathogens by the World Health Organization and is a major pathogen in tropical regions, causing superficial and invasive infections. C. tropicalis invasive infections are associated with a mortality rate of 55%–60% and the pathogen exhibits high azole resistance, a growing medical concern [3]. Previous studies indicate that exposure to agricultural azoles in natural environments can facilitate the emergence of clinical azole resistance in C. tropicalis [4,5].
A classical approach to studying the emergence of antifungal drug resistance is experimental evolution in which a drug-susceptible strain is grown in the presence of the drug under controlled laboratory conditions. Hu and colleagues employ an experimental evolution strategy in which azole-susceptible C. tropicalis strains are exposed to incrementally increasing concentrations of tebuconazole (TBZ), an azole-based fungicide used widely in agriculture (Fig 1A). Interestingly, the TBZ-resistant colonies obtained in the experimental evolution regime exhibit cross-resistance to the clinical azoles, too, [6] confirming the possibility of azole-resistant strains emerging in environmental niches.
Candida albicans and C. tropicalis, two abundant fungal pathogens, occur in a diploid (2n) state. Both species are primarily asexual fungi and are known to undergo parasexual mating, in which two diploid cells of opposite mating types fuse to generate tetraploid (4n) cells. The resulting cells undergo concerted chromosome loss, generating cells of various ploidy states [7,8]. However, the most common naturally occurring ploidy state in C. tropicalis is diploid. Interestingly, some TBZ-resistant colonies obtained are haploid (n), a previously unknown ploidy state for this organism, indicating that agricultural azoles can induce ploidy shifts in the C. tropicalis isolates.
Even C. albicans can exist in a mating-competent haploid state that is unstable [9]. Is haploidy a less stable state in C. tropicalis too? The haploid cells obtained by Hu and colleagues could mate with cells of different ploidy states (Fig 1B). Haploid strains of opposite mating types gave rise to diploid cells on mating. Haploid cells could even efficiently mate with diploid progenitors to yield triploid (3n) cells. This versatility, combined with the autodiploidization (n becomes 2n) observed in a fraction of haploid cells, raises the possibility that haploidy is a transition state in C. tropicalis. It is tempting to assume that frequent hybridization processes are facilitated by multiple ploidy states in an environmental niche. Indeed, potential hybrids of C. tropicalis were previously isolated from environmental sources [10]. The study by Hu and colleagues also reveals that haploid cells grow slower and are less virulent in a mouse model of systemic infection, indicating a reduced fitness compared to the diploid strains. The authors report the existence of haploid C. tropicalis clinical isolates by analyzing publicly available whole genome sequencing data revealing these isolates possess low heterozygosity in the genomic DNA, similar to the haploids obtained in the experimental evolution regime.
Besides ploidy plasticity, the authors find copy number variations in azole resistance-related genes like TAC1 and ERG11. TAC1 is a transcriptional regulator of drug-efflux pumps, and ERG genes are involved in the biosynthesis of ergosterol, a major fungal cell membrane component. In C. albicans, a closely related species of C. tropicalis, an isochromosome with the duplicated short arm of chromosome 5 bearing additional copies of ERG11 and TAC1 confers azole resistance [11]. Copy number variations of ERG11 has been implicated previously in azole resistance in multiple fungal pathogens including C. tropicalis [4,12,13]. The study of Hu and colleagues raises the possibility that isochromosome formation could be a general mechanism operating across species, triggered by clinical and agricultural azoles (Fig 1C). How stable are the isochromosomes if at all formed in C. tropicalis? Continuous passages in the absence of the drug can reveal the role of drug pressure in the maintenance of these additional copies of genes conferring drug resistance.
While the fully assembled genome, including haplotypes, of a diploid C. tropicalis strain is available [14], many aspects of the pathobiology of this fungal pathogen remain largely unexplored. The availability of a stable haploid state will facilitate gene function studies, including the importance of recessive alleles. In addition, studying changes in the proteome in a haploid compared to the diploids will help better understand the physiological impact of ploidy shifts. Haploidy and the ability of haploid cells to mate to produce various ploidy states may prove to be the arsenal of C. tropicalis to be one of the most successful drug-resistant fungal pathogens.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Case NT, Gurr SJ, Fisher MC, Blehert DS, Boone C, Casadevall A, et al. Fungal impacts on Earth’s ecosystems. Nature. 2025;638(8049):49–57. doi: 10.1038/s 41586-024-08419-4 39910383 PMC 11970531 · doi ↗ · pubmed ↗
- 2Lima R, Ribeiro FC, Colombo AL, de Almeida JN Jr. The emerging threat antifungal-resistant Candida tropicalis in humans, animals, and environment. Front Fungal Biol. 2022;3:957021. doi: 10.3389/ffunb.2022.957021 37746212 PMC 10512401 · doi ↗ · pubmed ↗
- 3Keighley C, Kim HY, Kidd S, Chen SC-A, Alastruey A, Dao A, et al. Candida tropicalis—a systematic review to inform the World Health Organization of a fungal priority pathogens list. Med Mycol. 2024;62(6):myae 040. doi: 10.1093/mmy/myae 040 38935905 PMC 11210624 · doi ↗ · pubmed ↗
- 4Hu T, Wang S, Bing J, Zheng Q, Du H, Li C, et al. Hotspot mutations and genomic expansion of ERG 11 are major mechanisms of azole resistance in environmental and human commensal isolates of Candida tropicalis. Int J Antimicrob Agents. 2023;62(6):107010. doi: 10.1016/j.ijantimicag.2023.107010 37863341 · doi ↗ · pubmed ↗
- 5Tseng K-Y, Chen Y-Z, Zhou Z-L, Tsai J-N, Tseng M-N, Liu H-L, et al. Detection in orchards of predominant azole-resistant Candida tropicalis genotype causing human Candidemia, Taiwan. Emerg Infect Dis. 2024;30(11):2323–32. doi: 10.3201/eid 3011.240545 39447155 PMC 11521182 · doi ↗ · pubmed ↗
- 6Hu T, Zheng Q, Cao C, Li S, Huang Y, Guan Z. An agricultural triazole induces genomic instability and haploid cell formation in the human fungal pathogen Candida tropicalis. P Lo S Biol. 2025.10.1371/journal.pbio.3003062 PMC 1196087640168394 · doi ↗ · pubmed ↗
- 7Forche A, Alby K, Schaefer D, Johnson AD, Berman J, Bennett RJ. The parasexual cycle in Candida albicans provides an alternative pathway to meiosis for the formation of recombinant strains. P Lo S Biol. 2008;6(5):e 110. doi: 10.1371/journal.pbio.0060110 18462019 PMC 2365976 · doi ↗ · pubmed ↗
- 8Seervai RNH, Jones SK Jr, Hirakawa MP, Porman AM, Bennett RJ. Parasexuality and ploidy change in Candida tropicalis. Eukaryot Cell. 2013;12(12):1629–40. doi: 10.1128/ec.00128-1324123269 PMC 3889571 · doi ↗ · pubmed ↗