The green side of splicing: algal spliceosome shows remarkable structural conservation
Wojciech P Galej

TL;DR
Researchers discovered that the spliceosome in a green alga has a structure very similar to other organisms, showing surprising conservation.
Contribution
The study presents the first Cryo-EM structure of the spliceosome from the green alga Chlamydomonas reinhardtii.
Findings
The algal spliceosome structure is remarkably similar to those in other organisms.
This finding suggests deep evolutionary conservation of the spliceosome across species.
Abstract
Spliceosome structures have been investigated extensively in the past decade, but most of the studies have been restricted to a handful of model organisms. New research by Lu et al, looks outside this field-of-view, reporting on the structure of the spliceosome from a unicellular green alga, Chlamydomonas reinhardtii, showing its extraordinary structural conservation. A new study solves the Cryo-EM structure of a Chlamydomonas reinhardtii spliceosome.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
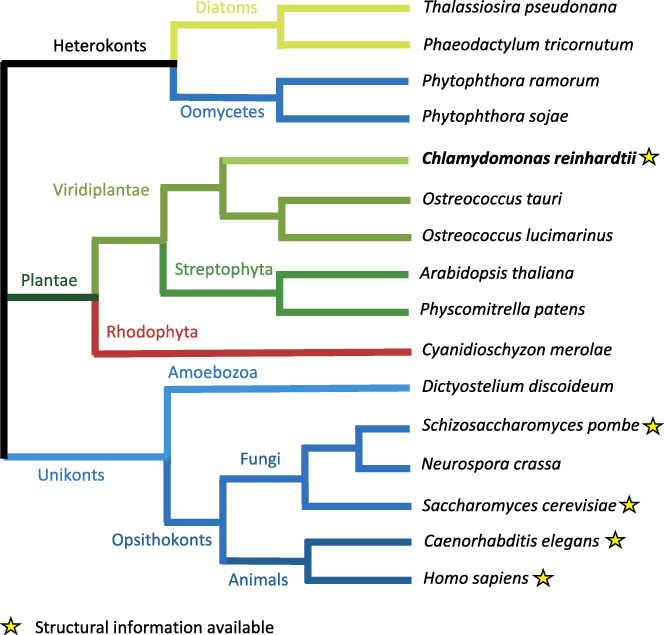 Figure 1
Figure 1 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProtist diversity and phylogeny · RNA Research and Splicing · Microbial Community Ecology and Physiology
The spliceosome is one of the cell’s most complex molecular machines, which catalyses the removal of non-coding introns from precursors of messenger RNAs in a process known as pre-mRNA splicing. Several decades of research have laid the foundations for mechanistic studies of pre-mRNA splicing. However, only when cryoEM structures of the spliceosome became available, the architectural and mechanistic insights into pre-mRNA splicing started to emerge (Kastner et al, 2019; Tholen and Galej, 2022). Despite rapid development of the entire field, structural studies of the spliceosome are mostly restricted to a few model organisms, such as Saccharomyces cerevisiae, Schizosaccharomyces pombe and Homo sapiens, and a handful of model pre-mRNA substrates. Using different species can facilitate trapping transient intermediate states, as recently demonstrated by the study of the post-splicing Intron-Lariat Spliceosome complex from Caenorhabditis elegans (Vorländer et al, 2024).
In the current issue of EMBO Journal, Lu et al (2024), investigate the spliceosome structure in Chlamydomonas reinhardtii, a unicellular green algae. Diverging from land plants over a billion years ago (Fig. 1), Chlamydomonas has a unique position in evolution as it contains many genes that are specific to both plant and animal lineages (Merchant et al, 2007). C. reinhardtii has an intron-rich genome, whose organisation and exon–intron structure resemble that of humans and land plants (Table 1). Exons of C. reinhardtii undergo alternative splicing, although to a much lower extent than in land plants. This makes it an interesting model organism to study alternative splicing regulation in a simplified system.Figure 1. Evolutionary tree of life highlighting (stars) model organisms for which at least one spliceosome structure has been determined experimentally.Adapted and modified with permission from Merchant et al (2007).Table 1. Exon–intron architecture of the genomes of selected model organisms.OrganismGenes with introns [%]Avg. Exons per geneAvg. Intron lengthAvg. Exon lengthChlamydomonas reinhardtii887.4336240Saccharomyces cerevisiae512561500Schizosaccharomyces pombe432107330Arabidopsis thaliana795165304Homo sapiens8593400282Reproduced from Labadorf et al (2010).
In the current study, Lu et al, purified native splicing complexes from C. reinhardtii using affinity tags on the components of the NTC/NTR complex (present in the spliceosome from its activation to disassembly) and analysed them by single-particle cryoEM. Structural analysis revealed that the purified complex is in the C* configuration; that is, it is ready for exon ligation, but the 3’-exon of the pre-mRNA is not yet engaged at the active site. The visualised core of the C. reinhardtii spliceosome is nearly identical to the core of the yeast and human C* complexes (Fica et al, 2017; Bertram et al, 2017), showing extraordinary structural conservation between the RNA catalytic cores and numerous scaffolding proteins.
The authors of the study discuss several subtle “metazoan-like” features of the C. reinhardtii spliceosome, including the presence of U5-40K protein, Aquarius helicase (Schmitzová et al, 2023), and an extended 16 nt-long duplex between the U6 snRNA and 5' splice site, all of which have been seen in the structures of human C* complexes. Another noteworthy observation is that the splicing factor, Prp17, is relatively well-resolved in C. reinhardtii, when compared to reports from other species. This could be potentially due to stabilising interactions between its N-terminal helix and a relatively short 5’ stem-loop of the U6 snRNA, which are unique to C. reinhardtii.
The conserved exon ligation factors (i.e. Prp18, Slu7 and Prp22) are missing in the reported structure, even though they are encoded by the C. reinhardtii genome. This could be caused by a relatively weak binding affinity in this organism or a result of the heterogeneity arising from undefined pre-mRNA species associated with this natively purified sample. If the former is true, it could explain why the authors managed to capture and enrich this particular complex, which has not undergone a full splicing cycle and subsequent disassembly. A thorough proteomic analysis could help address this question in the future. It is also possible that the purified complex is bound to a subset of inefficient and/or slow-splicing pre-mRNA substrates; here, analysis of its RNA composition could provide some crucial insights.
Recent reports of the human C* complexes revealed that numerous additional protein factors (such as Cactin, FAM32A and PRKRIP1) can regulate exon ligation (Zhan et al, 2022; Dybkov et al, 2023). There is no evidence of the presence of any of these additional C* factors in the structure resolved by Lu et al, and whether any have functional homologues in the C. reinhardtii remains to be seen.
Taken together, the work by Lu et al, provides a first into the core of the plant splicing machinery, revealing its remarkable structural conservation and extremely close similarity to evolutionarily distant organisms. At the same time, many questions related to splicing biology in Chlamydomonas, and more broadly, other plants, remain unanswered. For example, are there any plant-specific features and adaptations of the spliceosome? How do they affect its function, and can such features be exploited to advance our mechanistic understanding of pre-mRNA splicing? This and other questions remain to be investigated in the future.