Characterization of temperature-sensitive alleles of anillin-like Mid1 and polo kinase Plo1 in Schizosaccharomyces pombe
Joshua S. Park, Lesley A. Turner, Kathleen L. Gould, Alaina H. Willet

TL;DR
This study examines temperature-sensitive mutations in Mid1 and Plo1 proteins in fission yeast to better understand their roles in cell division.
Contribution
The study identifies specific mutations in Mid1 and Plo1 and highlights key domains critical for cytokinesis.
Findings
The Mid1 C2 domain is essential for proper cytokinesis.
The Plo1 kinase domain's C-terminal lobe is crucial for cell division.
Temperature-sensitive alleles reveal growth and morphological defects in mutant strains.
Abstract
The Schizosaccharomyces pombe anillin-like Mid1 is important for the correct positioning of the cell division site. A key regulator of Mid1 is the polo kinase Plo1 which is important for several mitotic and cytokinetic events including spindle formation and division site placement. Here, we defined the mutations within a set of temperature-sensitive mid1 and plo1 alleles and compared the growth and morphological defects of the strains. This work expands the repertoire of mid1 and plo1 mutants for studying cytokinesis and highlights the requirement of the Mid1 C2 domain and the Plo1 kinase domain C-terminal lobe as particularly important for cytokinesis.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
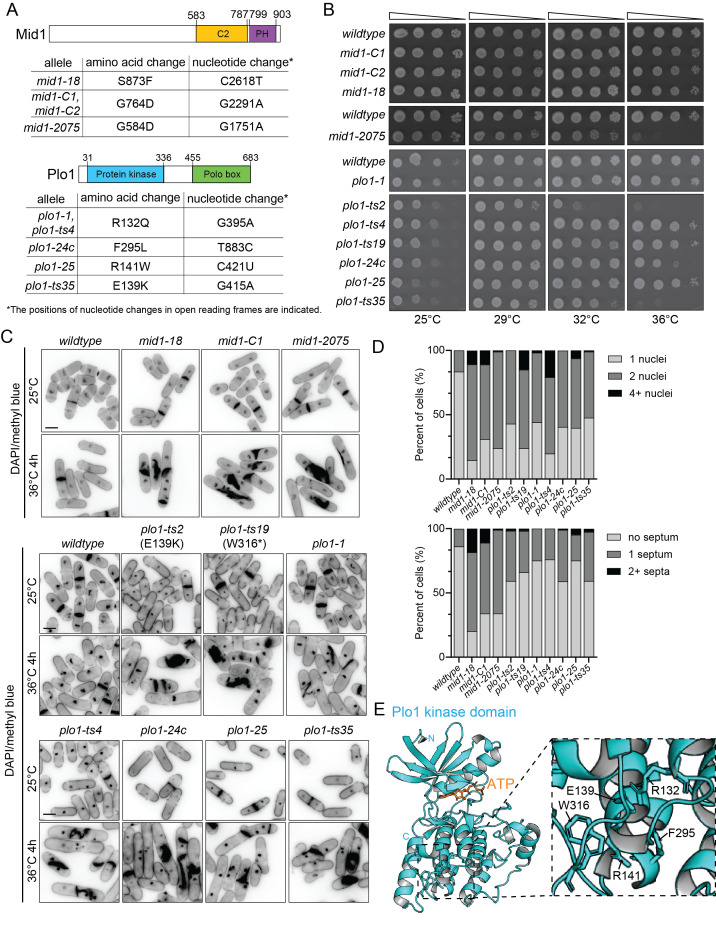 Figure 1
Figure 1|
|
|
|
|
KGY246 |
|
Lab stock |
|
KGY1001 |
|
|
|
KGY846 |
|
|
|
KGY1460 |
|
|
|
KGY16150-2 |
|
|
|
|
|
|
|
KGY16148-2 |
|
|
|
KGY16149-2 |
|
|
|
KGY1058 |
|
Balasubramanian et al., 1998 |
|
KGY1059 |
|
Balasubramanian et al., 1998 |
|
|
|
Balasubramanian et al., 1998 |
|
KGY442-2 |
|
This study |
|
KGY436-2 |
|
This study |
|
KGY3717 |
|
This study |
- —National Institute of General Medical Sciences (United States)https://ror.org/04q48ey07
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsATP Synthase and ATPases Research · Microtubule and mitosis dynamics · Protein Tyrosine Phosphatases
Description
The fission yeast Schizosaccharomyces pombe utilizes an actin- and myosin-based cytokinetic ring (CR) to accomplish cytokinesis (Cheffings et al., 2016; Glotzer, 2017; Mangione & Gould, 2019). The *S. pombe * CR is built from medial cytokinetic nodes established by the anillin-like Mid1 protein (reviewed in Rincon & Paoletti, 2012). These nodes condense into a coherent ring structure that will eventually constrict concomitant with septum ingression (reviewed in Willet et al., 2015). The precise spatial and temporal ordering of mitotic and cytokinetic events is critical for the faithful segregation of the genetic material into daughter cells. One mechanism by which mitosis and cytokinesis are coordinated is through the action of mitotic kinases that phosphorylate substrates in a cell cycle-dependent manner. One such enzyme is the S. pombe polo kinase Plo1 which operates downstream of Cdk1 to promote successful mitosis and cytokinesis (Ohkura et al., 1995; Tanaka et al., 2001). One of Plo1 's substrates is Mid1 and Plo1-dependent phosphorylation of Mid1 promotes the nuclear export of Mid1 (Almonacid et al., 2011; Bähler et al., 1998; Ohkura et al., 1995). This ensures that Mid1 can localize to the medial cytokinetic nodes at mitotic entry to promote CR formation (Almonacid et al., 2009, 2011). Here, we examine * mid1 * and * plo1 * temperature-sensitive alleles obtained from various genetic screens, some previously uncharacterized and many lacking information as to the sites of mutations (Bähler et al., 1998; Balasubramanian et al., 1998; MacIver et al., 2003).
To better understand the nature of the * mid1 * and * plo1 * alleles, the respective open reading frames were amplified from nine strains ( mid1-18, mid1-C1, *mid1-C2, mid1-2075, plo1-1, plo1-ts4, plo1-24c, plo1-25 * and plo1-ts35 ) in which the relevant mutations have not been reported (Rutherford et al., 2024) and the PCR products sequenced. In each case, a single point mutation was detected ( Figure 1A ). mid1-18 encodes a serine to phenylalanine substitution at position 873 (S873F), both mid1-C1 and mid1-C2 encode a glycine to aspartic acid substitution at position 764 (G764D) and mid1-2075 encodes a glycine to aspartic acid substitution at position 584 ( Figure 1A ). All of the mutated amino acids reside in structured regions of the protein ( Figure 1A ). Another previously characterized * mid1 * allele, mid1-366 , encodes a different mutation within the C2 domain (G718D) (Chang et al., 1996; Sun et al., 2015). Thus, our results bring the total number of distinct * mid1 * temperature-sensitive alleles to four.
Two * plo1 * alleles ( *plo1-1 * and *plo1-ts4) * encode the same mutation causing an arginine to glutamine change at residue 132 (R132Q). plo1-24c encodes a mutation resulting in a change at residue 295 from phenylalanine to leucine, plo1-25 encodes an arginine to tryptophan substitution at residue 141 (R141W) and *plo1-ts35 * encodes a glutamic acid to lysine change at position 139 (E139K) ( Figure 1A ). E139K is the same mutation previously identified in the plo1-ts2 allele (MacIver et al., 2003). Thus, in addition to two previously sequenced temperature-sensitive * plo1 * alleles ( *plo1-ts2 * and
- plo1-ts19* ), there are three additional distinct temperature-sensitive
plo1 * alleles.
To examine the * mid1 * and * plo1 * mutant cohorts further, we compared the temperature-sensitivity of each strain with a growth assay. *mid1-18, mid1-C1 and mid1-C2 * grew similar to wildtype at all temperatures but mid1-2075 showed almost no growth at 36°C ( Figure 1B ). Two * plo1 * alleles, plo1-ts2 and plo1-ts35, had almost no growth at 36°C and reduced growth at 32°C, and all other * plo1 * alleles were slightly impaired in growth compared to wildtype at 36°C ( Figure 1B ). These results are consistent with previously published growth assays in which some of the mutants were examined (MacIver et al., 2003; Petersen & Hagan, 2005; Rachfall et al., 2014).
Finally, to visualize and compare the cell phenotypes, we examined each mutant by staining for nuclei and septa after the cells were grown at 25°C and then shifted to 36°C for 4 hours. We found that all temperature sensitive alleles looked similar to wildtype at 25°C, but at 36°C the cells were multinucleated with abnormal septa present ( Figure 1C- D). Our data are consistent with previous descriptions of the subset of previously studied alleles (Bähler et al., 1998; Balasubramanian et al., 1998; Bhutta et al., 2014; Huang et al., 2008; MacIver et al., 2003; Wachtler et al., 2006).
All * mid1 * temperature-sensitive alleles displayed phenotypes similar to *mid1∆ * (Chang et al., 1996; Sohrmann et al., 1996), suggesting that they are loss-of-function alleles. mid1-18 has a mutation in the PH domain while mid1-C1 , *mid1-C2, mid1-2075 * and mid1-366 contain mutations in the C2 domain ( Figure 1A ). The C2 domain appears to directly bind the plasma membrane and is required for Mid1 function (Lee & Wu, 2012; Sun et al., 2015). Cells with a mutant Mid1 lacking the entire PH domain have only mild cell division site positioning defects, thus the function of the PH domain is not fully clear (Lee & Wu, 2012; Paoletti & Chang, 2000). It has been suggested that the PH domain may be important for protein stability and/or play a role in membrane binding in collaboration with the C2 domain, like human anillin (Hall et al., 2024; Liu et al., 2012; Sun et al., 2015). Taken together, we speculate that the mid1-18 allele produces a less stable protein at high temperatures while the other alleles disrupt C2 domain function.
Plo1 has two essential domains, the N-terminal kinase domain and the C-terminal polo box domain (PBD) (Reynolds & Ohkura, 2003) ( Figure 1A ). While the kinase domain phosphorylates protein substrates, the PBD binds both substrates and protein partners to direct subcellular localization (Park et al., 2010). Interestingly, all mutations encoded by the * plo1 * temperature sensitive alleles map within the kinase domain and none are within the PBD ( Figure 1A ). The one unique allele previously sequenced is plo1-ts19 which contains an early stop codon at tryptophan 316 (MacIver et al., 2003). The W316* mutation truncates an ɑ-helix within the kinase domain and eliminates an unstructured region and the PBD (MacIver et al., 2003) which is a surprising result given the reported requirement for the PBD for Plo1 function (Reynolds & Ohkura, 2003).
To follow up these observations, we used AlphaFold3 to model the Plo1 kinase domain with an ATP molecule (Abramson et al., 2024), and mapped the mutated residues encoded by the * plo1 * alleles onto the predicted structure. We found that all of the mutated residues clustered in the C-terminal lobe of the kinase domain ( Figure 1E ). Deciphering if ATP binding, substrate binding or other protein partner binding is defective in these * plo1 * alleles will be exciting direction for future studies. Further, how cells tolerate a lack of the Plo1 PBD will be an interesting avenue of study.
Methods
Yeast methods
S. pombe strains were grown in yeast extract (YE) and standard S. pombe mating, sporulation, and tetrad dissection techniques were used to construct new strains (Moreno et al., 1991).
Molecular biology methods
The * mid1 * open reading frames from *mid1-C1 * and *mid1-C18 * cells were amplified by generating a PCR product with an oligonucleotide 74 bp upstream of the start site (GTTGTACTTCAGGGTGCTTA) and an oligonucleotide 380 bp downstream of the stop codon (AGGTTCTCCATCTCATGGCT) (Integrated DNA technologies). The * mid1 * open reading frame was amplified from *mid1-C2 * cells using two sets of overlapping oligonucleotides. The first PCR product was generated with the above-mentioned oligonucleotide 74 bp upstream of the start site and an oligonucleotide 1350 bp into the open reading frame (GTTGCATTGATGGGTGACGT). The second PCR product was produced with an oligonucleotide 1200 bp within the open reading frame (GTATGGTCATGGATCTGTAACG) and the above-mentioned oligonucleotide 380 bp downstream of the stop codon.
The * plo1 * alleles were amplified by generating two PCR products. One PCR reaction used an oligonucleotide 40 bp upstream of the start site (GCAACCACTTTGTTTACCCTCA) and an oligonucleotide 1100 bp into the open reading frame of * plo1 * (TGGACTTAAAACACTTGGTAATATTCG) (Integrated DNA technologies). The second PCR reaction used an oligonucleotide 900 bp into the open reading frame (TCCAGATGAAATTTTACATTCAATGCCT) with an oligonucleotide 20 bp downstream of the stop codon (GCATAGTAACTTAACGCCCAAGTA).
The PCR products were sequenced by Plasmidsaurus using Oxford Nanopore Technology with custom analysis and annotation.
Microscopy and image analysis
Strains for fixed-cell imaging experiments were grown at 25°C in YE and then shifted to 36°C for 3 hours. Cells were fixed with 70% ethanol for DAPI and methyl blue (MB) staining as described previously (Roberts-Galbraith et al., 2009). Images were acquired using a Zeiss Axio Observer inverted epifluorescence microscope with Zeiss 63× oil (1.46 NA) objective and captured using Zeiss ZEN 3.0 (Blue edition) software. A singular medial Z slice was obtained. All images were further processed using ImageJ (Schindelin et al., 2012).
AlphaFold3 structural prediction
Protein structure predictions were generated with the AlphaFold3 server (Abramson et al., 2024) and visualized using the PyMOL molecular graphics system (version 3.0, Schrodinger, LLC).
Reagents
The strains used in this study and their genotypes are listed below.
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abramson Josh Adler Jonas Dunger Jack Evans Richard Green Tim Pritzel Alexander Ronneberger Olaf Willmore Lindsay Ballard Andrew J. Bambrick Joshua Bodenstein Sebastian W. Evans David A. Hung Chia-Chun O’Neill Michael Reiman David Tunyasuvunakool Kathryn Wu Zachary ŽemgulytėAkvilė Arvaniti Eirini Beattie Charles Bertolli Ottavia Bridgland Alex Cherepanov Alexey Congreve Miles Cowen-Rivers Alexander I. Cowie Andrew Figurnov Michael Fuchs Fabian B. Gladman Hannah Jain Rishub Khan Yousuf A. Low Caroline M. R. Perlin Kuba Potapenk · doi ↗ · pubmed ↗
- 2Almonacid Maria Celton-Morizur Séverine Jakubowski Jennifer L. Dingli Florent Loew Damarys Mayeux Adeline Chen Jun-Song Gould Kathleen L. Clifford Dawn M. Paoletti Anne 201131 Temporal Control of Contractile Ring Assembly by Plo 1 Regulation of Myosin II Recruitment by Mid 1/Anillin Current Biology 2160960-982247347910.1016/j.cub.2011.02.00321376600 PMC 3088474 · doi ↗ · pubmed ↗
- 3Almonacid Maria Moseley James B. Janvore Julie Mayeux Adeline Fraisier Vincent Nurse Paul Paoletti Anne 200961 Spatial Control of Cytokinesis by Cdr 2 Kinase and Mid 1/Anillin Nuclear Export Current Biology 19110960-982296196610.1016/j.cub.2009.04.02419427212 · doi ↗ · pubmed ↗
- 4Bähler Jürg Steever Alexander B. Wheatley Sally Wang Yu-li Pringle John R. Gould Kathleen L. Mc Collum Dannel 19981214 Role of Polo Kinase and Mid 1p in Determining the Site of Cell Division in Fission Yeast The Journal of Cell Biology 14360021-95251603161610.1083/jcb.143.6.16039852154 PMC 2132972 · doi ↗ · pubmed ↗
- 5Balasubramanian Mohan K Mc Collum Dannel Chang Louise Wong Kelvin C Y Naqvi Naweed I He Xiangwei Sazer Shelley Gould Kathleen L 199871 Isolation and Characterization of New Fission Yeast Cytokinesis Mutants Genetics 14931943-26311265127510.1093/genetics/149.3.12659649519 PMC 1460233 · doi ↗ · pubmed ↗
- 6Bhutta Musab Mc Inerny Christopher Gould Gwyn 20141125 ESCRT Function in Cytokinesis: Location, Dynamics and Regulation by Mitotic Kinases International Journal of Molecular Sciences 15121422-0067217232173910.3390/ijms 15122172325429432 PMC 4284674 · doi ↗ · pubmed ↗
- 7Chang Fred Woollard Alison Nurse Paul 199611 Isolation and characterization of fission yeast mutants defective in the assembly and placement of the contractile actin ring Journal of Cell Science 10910021-953313114210.1242/jcs.109.1.1318834798 · doi ↗ · pubmed ↗
- 8Cheffings Thomas H. Burroughs Nigel J. Balasubramanian Mohan K. 201681 Actomyosin Ring Formation and Tension Generation in Eukaryotic Cytokinesis Current Biology 26150960-9822 R 719R 73710.1016/j.cub.2016.06.07127505246 · doi ↗ · pubmed ↗