Deguelin promotes longevity and healthspan through C. elegans fmo-4
Angela M Tuckowski, Shijiao Huang, Kelly Chambers, Brandon Buscher, Scott F Leiser

TL;DR
Deguelin, a mitochondrial complex I inhibitor, extends lifespan and healthspan in C. elegans by acting through the FMO-4 enzyme.
Contribution
The study identifies FMO-4 as a key dependency for deguelin's longevity-promoting effects in C. elegans.
Findings
Deguelin increases lifespan in C. elegans.
Deguelin's effects on longevity depend on FMO-4 and other FMO enzymes.
FMO enzymes may work together to promote longevity.
Abstract
There are multiple approaches to longevity interventions in Caenorhabditis elegans , including genetic factors that are necessary or sufficient for lifespan extension and pharmacological agents that modify physiology to extend lifespan. Many pharmacological interventions act through known genetic pathways to promote longevity. Here, we show that the mitochondrial complex I inhibitor, deguelin, promotes lifespan extension and healthspan in an fmo-4- dependent manner. Our results confirm that deguelin increases lifespan and indicate that deguelin induces and requires multiple FMO enzymes to extend lifespan in C. elegans , suggesting these enzymes may promote longevity in a coordinated fashion.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
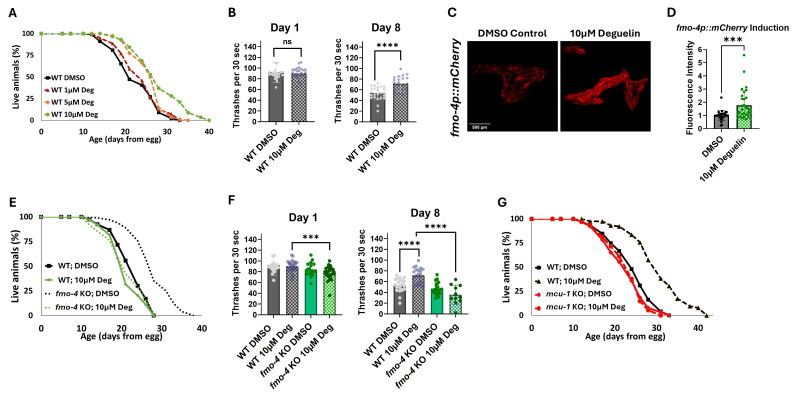 Figure 1
Figure 1|
|
|
|
|
WT or
|
WT |
CGC |
|
|
|
CGC |
|
|
|
CGC |
|
|
|
Suny Bioscience |
|
|
|
|
|
Deguelin |
Sigma |
D0817-5MG |
|
DMSO |
Fisher Scientific |
BP213-100 |
|
Sodium Azide |
Sigma |
S2002-5G |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBirth, Development, and Health · Genetics, Aging, and Longevity in Model Organisms · Nuclear Structure and Function
Description
Aging is a complex biological process that affects all living organisms, leading to a decline in physiological functions and an increased susceptibility to diseases ^1^ . Studying the aging process is crucial because it allows for a better understanding of how to prevent or delay the onset of multiple age-related chronic diseases simultaneously. The nematode * Caenorhabditis elegans * ( * C. elegans * ) has emerged as a powerful model organism for studying aging due to its short lifespan, well-characterized genetics, and conserved aging pathways ^2^ . Recent research has focused on identifying longevity interventions in * C. elegans * , including genetic factors and pharmacological agents that can extend lifespan and improve healthspan, potentially offering insights into interventions that could promote healthy aging in humans.
A gene family of particular interest are the flavin-containing monooxygenase ( fmo ) genes. From this family, * fmo-2 * and * fmo-4 * are both sufficient and necessary for lifespan extension in * C. elegans * ^3,4^ . These genes play crucial roles in altering cellular metabolism, including one carbon metabolism ^5^ and calcium regulation between the endoplasmic reticulum and mitochondria ^4^ , contributing to the longevity phenotype. Pharmacologically, compounds like deguelin, a natural rotenoid isolated from plants of the
- Leguminosae* family, have gained attention for their potential to extend lifespan through the manipulation or inhibition of cellular processes ^6-8^ . For instance, deguelin treatment has been successfully tested as an anti-tumor agent in human cells lines, inhibits mitochondrial complex I, inhibits mammalian target of rapamycin complex I (mTORC1), and extends lifespan in wild-type (WT)
C. elegans * ^6-8^ .
Our lab previously demonstrated that these interventions can act in the same genetic pathway to extend lifespan. Specifically, we have shown that deguelin requires * fmo-2 * to promote longevity in * C. elegans , * suggesting that * fmo-2 * acts downstream of deguelin-mediated lifespan extension ^6^ . Furthermore, we established that * fmo-2 * also requires * fmo-4 * for its overexpression to extend lifespan ^4^ . Given these interconnected relationships, here we investigate whether deguelin and * fmo-4 * genetically interact in the context of longevity. Interestingly, our recent publication shows that * fmo-4 * extends lifespan downstream of the inhibition of mTORC1 ( * rsks-1 * RNA interference) and has genetic ties to mitochondrial metabolism ^4^ . Considering these similarities between * fmo-4 * and deguelin, we hypothesized that deguelin may extend lifespan and promote healthspan through the induction of * fmo-4 * , and we sought to investigate the downstream effectors involved in this process.
To address this hypothesis, we first examined the effects of deguelin on WT * C. elegans * lifespan. Our lab published that deguelin, in addition to multiple other compounds, extends lifespan of WT worms ^6^ , and so we aimed to validate these data using a range of concentrations. Our results confirm that deguelin extends WT lifespan in a dose-dependent manner ( ** Figure 1A ** ). To further investigate the quality of life during this extended lifespan, we assessed the healthspan of deguelin-treated worms. Our data show that deguelin not only extends lifespan but also promotes healthspan in middle-aged (day 8 of adulthood) WT worms ( ** Figure 1B ** ), indicating that the compound may improve overall health and functionality during aging.
To explore the molecular mechanisms underlying deguelin's effects, we focused on the potential involvement of * fmo-4 * . We recently found that * fmo-4 * is required for mTOR pathway gene rsks-1- RNAi-mediated longevity, and that it extends lifespan by regulating calcium signaling between the ER and mitochondria ^4^ . This is interesting because deguelin is an inhibitor of both mTORC1 and mitochondrial complex I ^7^ , providing more evidence for a potential interaction between the two longevity interventions. Using a transcriptional * fmo-4 ::mCherry * reporter, we observed that deguelin treatment induces the expression of * fmo-4 * in * C. elegans * by ~2-fold. Fluorescent images and quantification reveal this significant increase in * fmo-4 * expression in deguelin-treated worms compared to the DMSO-treated control worms ( ** Figure 1C- D ** ). This induction suggests that * fmo-4 * is regulated by deguelin and may play a crucial role in mediating the longevity-promoting effects of deguelin. To confirm the requirement of * fmo-4 * in deguelin-mediated lifespan extension, we performed a lifespan assay using * fmo-4 * knockout (KO) worms. Our results demonstrate that the lifespan-extending effect of deguelin is abolished in * fmo-4 * KO worms ( ** Figure 1E ** ), indicating that * fmo-4 * is indeed necessary for and downstream of deguelin to extend lifespan in * C. elegans * . Similarly, we found that * fmo-4 * is also required for deguelin to promote healthspan in day 8 adult worms ( ** Figure 1F ** ), further supporting the critical role of * fmo-4 * in mediating the beneficial effects of deguelin on aging.
To gain insight into the downstream mechanisms of deguelin-mediated longevity, we investigated the involvement of * mcu-1 * , a downstream effector of fmo-4- mediated longevity ^4,9^ . * mcu-1 * is a mitochondrial calcium uniporter that has been implicated in aging, stress response, and calcium regulation ^4,9^ . Since * fmo-4 * is required for the longevity and healthspan effect seen with deguelin treatment, and since * mcu-1 * acts downstream of * fmo-4 ^4^ , * we hypothesized that * mcu-1 * would also be required for the benefits of deguelin treatment. Our data show that * mcu-1 * is required for deguelin-mediated longevity ( ** Figure 1G ** ), suggesting that the effects of deguelin on lifespan extension may involve modulation of mitochondrial calcium homeostasis through the * fmo-4 / mcu-1 * pathway.
While our findings provide compelling evidence for the role of deguelin in promoting longevity and healthspan in * C. elegans * through the induction of * fmo-4 * , there are several limitations to consider. First, the exact mechanism by which deguelin induces * fmo-4 * expression remains to be elucidated. It is possible that deguelin, * fmo-2 , * and * fmo-4 * are working in the same genetic pathway to promote these health benefits but more work needs to be done to confirm this involvement. Additionally, further research is needed to determine whether the effects of deguelin on lifespan and healthspan are conserved in other organisms, including mammals. Finally, potential off-target effects of deguelin and long-term consequences of its administration should be carefully evaluated in future studies.
In summary, our data demonstrate that deguelin extends lifespan and promotes healthspan in * C. elegans * in an * fmo-4 * -dependent manner. We find that deguelin induces the expression of * fmo-4 * and that both * fmo-4 * and its downstream effector * mcu-1 * are required for deguelin-mediated longevity. These findings contribute to our understanding of the molecular mechanisms underlying lifespan extension and highlight the potential of deguelin as a pro-longevity compound. Future directions for this research include investigating the upstream regulators of * fmo-4 * induction by deguelin, exploring the conservation of this pathway in higher organisms, and evaluating the potential of deguelin or related compounds as interventions to promote healthy aging in humans.
Methods
Strains and Maintenance
Standard * C. elegans * cultivation procedures were used as previously described ^3^ . Worm strains were maintained on solid nematode growth medium (NGM) using *E. coli * OP50 throughout life. Worms were transferred using a platinum wire. All worm strains were maintained at 20°C.
Lifespan Assays
Gravid adult worms were placed on NGM plates seeded with *E. coli * OP50 for three hours. Then the adults were removed and eggs were allowed to hatch and develop to day 1 adulthood at 20°C. Adult worms were transferred to NGM plates containing 25 µg/mL carbenicillin, floxuridine (FUdR), and either dimethylsulfoxide (DMSO), 1 µM deguelin (Sigma, D0817), 5 µM deguelin, or 10 µM deguelin. Additionally, these plates were seeded with 200 µL of paraformaldehyde (PFA) killed *E. coli * OP50 at a concentration of 3x. Approximately 70 worms were transferred to fresh plates on day 1, day 2, day 4, and day 6 of adulthood. Two plates per strain per condition were tested per replicate experiment. Experimental animals were scored every 2-3 days and considered dead when they did not move in response to prodding under a dissection microscope. Worms that crawled off the plate were not considered, but ruptured worms were considered as previously described ^3^ . Three replicates were performed for each lifespan assay.
Thrashing Assay
Worms were synchronized by placing 10 gravid adult worms on NGM plates seeded with *E. coli * OP50 and allowing them to lay eggs for 2 hours at 20°C. The gravid adults were removed and the eggs were allowed to hatch and develop at 20°C until larval stage 2 (L2). At this stage, the L2 worms were transferred to NGM plates containing either DMSO or 10 µM deguelin, seeded with paraformaldehyde killed *E. coli * OP50 . On day 1 adulthood, worms were placed in a drop of M9 solution, as previously described. The body bends were counted at maximum rate for 30 seconds. Thrashing was assayed on day 1 and day 8 of adulthood. The worms that were not used for the day 1 assay were transferred to fresh plates containing either DMSO or deguelin two times until they were ready to be assayed. Three replicates were performed. Data were analyzed in GraphPad Prism using unpaired two-tailed t tests with Welch's correction.
** * fmo-4 * Induction on Deguelin **
Gravid * fmo-4 p::mCherry * transcriptional reporter adult animals were placed on NGM plates seeded with *E. coli * OP50 . After 3 hours, adults were removed and the eggs were allowed to develop at 20°C until they reached larval stage 4 (L4). Then 30 of the L4 worms were transferred to NGM plates containing either DMSO or 10 µM deguelin and seeded with paraformaldehyde (PFA) killed *E. coli * OP50 . The worms were incubated for 24 hours at 20°C. Then ~20 worms per condition were picked off these plates and added to unseeded NGM plates, anesthetized in 0.5 M sodium azide (Sigma), and imaged at 6.3x magnification with the LASx software and Leica scope using the mCherry fluorescence channel. Three replicates were performed. Each worm was measured for fluorescence in ImageJ. Data were analyzed in GraphPad Prism using t tests.
Statistical Analyses
Log-rank test was used to derive p-value for lifespan assays using p < 0.05 cut-off threshold compared to DMSO or wild-type controls. Cox regression was also used to assess interactions between genotype and condition for lifespans using p < 0.01 cut-off threshold compared to controls. **Supplemental Data 1 ** provide the results of the Log-rank test and Cox regression analyses, which were run in RStudio.
Reagents
**: **
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Guo J Huang X Dou L Yan M Shen T Tang W Li J 20221216 Aging and aging-related diseases: from molecular mechanisms to interventions and treatments.Signal Transduct Target Ther 712095-990739139110.1038/s 41392-022-01251-036522308 PMC 9755275 · doi ↗ · pubmed ↗
- 2Kenyon CJ 2010325 The genetics of ageing.Nature 46472880028-083650451210.1038/nature 0898020336132 · doi ↗ · pubmed ↗
- 3Leiser SF Miller H Rossner R Fletcher M Leonard A Primitivo M Rintala N Ramos FJ Miller DL Kaeberlein M 20151119 Cell nonautonomous activation of flavin-containing monooxygenase promotes longevity and health span.Science 35062660036-80751375137810.1126/science.aac 925726586189 PMC 4801033 · doi ↗ · pubmed ↗
- 4Tuckowski Angela M Beydoun Safa Kitto Elizabeth S Bhat Ajay Howington Marshall B Sridhar Aditya Bhandari Mira Chambers Kelly Leiser Scott F 2025214 fmo-4 promotes longevity and stress resistance via ER to mitochondria calcium regulation in C. eleganse Life 132050-084X 10.7554/elife.99971 PMC 1182848439951337 · doi ↗ · pubmed ↗
- 5Choi HS Bhat A Howington MB Schaller ML Cox RL Huang S Beydoun S Miller HA Tuckowski AM Mecano J Dean ES Jensen L Beard DA Evans CR Leiser SF 202322 FMO rewires metabolism to promote longevity through tryptophan and one carbon metabolism in C. elegans.Nat Commun 14156256210.1038/s 41467-023-36181-036732543 PMC 9894935 · doi ↗ · pubmed ↗
- 6Huang S Cox RL Tuckowski A Beydoun S Bhat A Howington MB Sarker M Miller H Ruwe E Wang E Li X Gardea EA De Nicola D Peterson W Carrier JM Miller RA Sutphin GL Leiser SF 2024524 Fmo induction as a tool to screen for pro-longevity drugs.Geroscience 4652509-27154689470610.1007/s 11357-024-01207-y 38787463 PMC 11335711 · doi ↗ · pubmed ↗
- 7Carpenter EL Chagani S Nelson D Cassidy PB Laws M Ganguli-Indra G Indra AK 2019618 Mitochondrial complex I inhibitor deguelin induces metabolic reprogramming and sensitizes vemurafenib-resistant BRAF(V 600E) mutation bearing metastatic melanoma cells.Mol Carcinog 5890899-19871680169010.1002/mc.2306831211467 PMC 6692247 · doi ↗ · pubmed ↗
- 8Xu H Li X Ding W Zeng X Kong H Wang H Xie W 2015225 Deguelin induces the apoptosis of lung cancer cells through regulating a ROS driven Akt pathway.Cancer Cell Int 151475-2867252510.1186/s 12935-015-0166-425741219 PMC 4349657 · doi ↗ · pubmed ↗