Characterization and comparison of Schizosaccharomyces pombe cdc15 temperature-sensitive mutants
Lesley A. Turner, Aleksandar Vjestica, Alaina H. Willet, Snezhana Oliferenko, Kathleen L. Gould

TL;DR
This paper studies temperature-sensitive mutants of the Cdc15 protein in fission yeast to better understand its role in cell division.
Contribution
A new cdc15 allele is identified that reveals Cdc15's role in recruiting the early secretory pathway to the division site.
Findings
Multiple cdc15 temperature-sensitive mutants were characterized and compared.
A new cdc15 allele was identified that affects secretory pathway recruitment.
The mutants provide new tools for studying cytokinesis in S. pombe.
Abstract
The F-BAR protein Cdc15 is essential for cytokinesis in the fission yeast Schizosaccharomyces pombe , playing a key scaffolding role and connecting the actomyosin-based cytokinetic ring to the plasma membrane. Here, we compared cdc15 temperature-sensitive mutants isolated in multiple genetic screens. We determined the mutations within each cdc15 mutant allele and analyzed their growth at different temperatures. Additionally, we report a new cdc15 allele that highlights the requirement for Cdc15 in the recruitment of the early secretory pathway to the cellular division site. The new mutants described here expand the toolkit for studying cytokinesis in S. pombe .
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
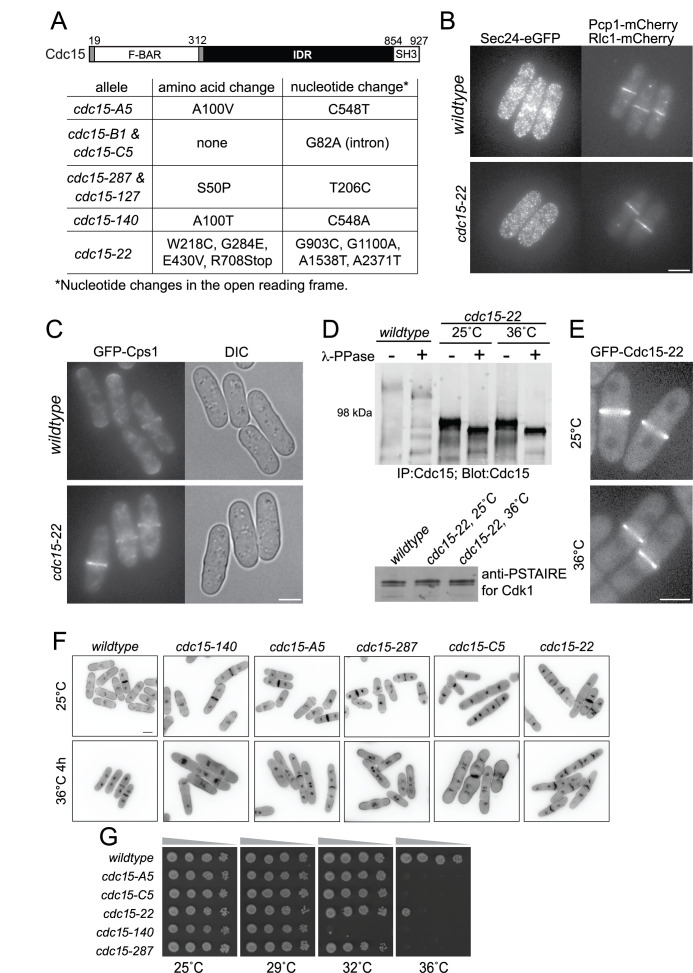 Figure 1
Figure 1|
|
|
|
|---|---|---|
|
KGY114-2 |
|
This study |
|
KGY188 |
|
Nurse et al., 1976 |
|
KGY282-2 |
|
This study |
|
KGY642-3 |
|
Nurse et al., 1976 |
|
KGY749 |
|
Chang et al., 1996 |
|
KGY1077 |
|
Balasubramanian et al., 1998 |
|
KGY1078 |
|
Balasubramanian et al., 1998 |
|
KGY1079 |
|
Balasubramanian et al., 1998 |
|
SO6279 |
|
This study |
|
SO5986 |
|
This study |
|
SO7141 |
|
This study |
|
SO7350 |
|
This study |
|
SO628
|
|
This study |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal and yeast genetics research · Cellular transport and secretion · Microtubule and mitosis dynamics
Description
In the fission yeast Schizosaccharomyces pombe, c ytokinesis requires the function of the plasma membrane (PM)-bound actin- and myosin-based cytokinetic ring (CR) that is assembled at the cell equator at the onset of mitosis. CR constriction at the end of anaphase ultimately results in the physical separation of two daughter cells.
One of the proteins mediating CR attachment to the PM is the essential dimeric F-BAR protein Cdc15 ( Figure 1A, top panel). Cdc15 is one of the first and most abundant components detected at the CR (Fankhauser et al., 1995; Nurse et al., 1976; Wu et al., 2003; Wu and Pollard, 2005). As cells progress into mitosis, the N-terminal dimeric F-BAR domain of Cdc15 oligomerizes, binds membranes, and interacts with the formin Cdc12 and the paxillin-related Pxl1 (Carnahan and Gould, 2003; McDonald et al., 2015; Snider et al., 2022; Snider et al., 2020; Willet et al., 2015). A C-terminal SH3 domain provides a second binding site for Pxl1 and also interacts with other CR components including the C2 domain protein Fic1 (Bhattacharjee et al., 2020; Cortes et al., 2015; Martin-Garcia et al., 2018; Ren et al., 2015; Roberts-Galbraith et al., 2009). Between the F-BAR and SH3 domains lies a predicted intrinsically disordered region (IDR) essential for Cdc15 function (Mangione et al., 2019) . Reducing the amount of Cdc15 , preventing Cdc15 membrane binding and/or F-BAR domain-mediated oligomerization, or deleting the Cdc15 SH3 domain destabilizes the CR during anaphase and leads to cytokinesis failure (Arasada and Pollard, 2011; McDonald et al., 2015; Roberts-Galbraith et al., 2009) . Abrogating Cdc15 function also prevents the enrichment of the early secretory machinery, transitional ER (tER) exit sites, at the nascent division site (Vjestica et al., 2008) .
c dc15 temperature-sensitive mutants have been isolated in genetic screens for general cell cycle regulators and also in those targeting cytokinesis factors (Balasubramanian et al., 1998; Chang et al., 1996; Nurse et al., 1976) . We isolated a novel * cdc15 * allele, cdc15-22 , in a genetic screen for mutants where the CR assembled and remained stable over an extended period of time but failed to recruit tER marked by the eGFP-tagged COPII coat protein Sec24 ( Figure 1B ). Yet, the cell wall synthase Cps1 / Bgs1 localized normally to the division site in cdc15-22 cells at the restrictive temperature of 36˚C ( Figure 1C ). The mutations within several * cdc15 * mutant alleles, including cdc15-22 , have not been identified or reported in PomBase (Rutherford et al., 2024) .
To determine the molecular lesions in all * cdc15 * temperature-sensitive mutants, we amplified and sequenced the * cdc15 * open reading frame (ORF) from each strain ( Figure 1A, bottom panel). In one of the first described alleles (Nurse et al., 1976) , cdc15-140 , we found a single mutation conferring a single amino acid substitution (A100T) within the F-BAR domain. The same amino acid was changed to a valine in cdc15-A5 . A distinct mutation within the F-BAR domain was also identified in cdc15-287 and this was an identical change to that in cdc15-127 . Since the F-BAR mediates membrane binding, it is likely that the F-BAR domain becomes destabilized at the non-permissive temperature. Two alleles, cdc15-B1 and cdc15-C5 , contained the same mutation at the 5'end of an intron. This mutation does not lead to an amino acid substitution. Instead, it likely inhibits splicing and causes a decrease in protein production exacerbated at the non-permissive temperature. Indeed, significantly decreasing Cdc15 levels leads to cytokinesis failure and cell death (Arasada and Pollard, 2011) . The last allele we sequenced was cdc15-22 . This allele contained three amino acid substitutions and a stop codon at the amino acid 708. Introduction of individual cdc15-22 mutations into the wild-type * cdc15 * locus indicated that the premature stop codon was responsible for the cytokinesis failure of cdc15-22 cells. We previously found that C-terminal truncations to amino acid 752 or amino acid 710 were viable (Mangione et al., 2019; Roberts-Galbraith et al., 2009) . However, these truncation alleles grew slowly and showed a variety of cell division and morphological defects similar to cdc15-22 cells.
To validate the sequencing results from cdc15-22 , we performed an immunoprecipitation and immunoblotting experiment. This confirmed that Cdc15-22 was a shorter protein ( Figure 1D ). However, the overall protein levels were not diminished, in fact they appeared increased, and the truncated protein was still phosphorylated. Cdc15's ability to oligomerize and bind membrane and protein partners is regulated by its phosphorylation state (Bhattacharjee et al., 2023; Bhattacharjee et al., 2020; Fankhauser et al., 1995; Roberts-Galbraith et al., 2010). N-terminal tagging of Cdc15-22 mutant protein with eGFP showed that it was efficiently recruited to the CR suggesting that its phosphoregulation was largely intact ( Figure 1E ).
To compare cellular phenotypes of the strains, we examined each mutant by staining for nuclei and septa at the permissive temperature of 25°C or after the shift to the restrictive temperature of 36˚C for 4 hours. The phenotypes of the cdc15-140 , cdc15-A5 , and cdc15-287 cells were comparable to each other, with an accumulation of multiple nuclei and occasionally a septa at the restrictive temperature ( Figure 1F ). In contrast, the cdc15-C5 and *cdc15-22 * cultures contained cells with multiple nuclei and septa even at 25˚C and cells that became wider and bulged at 36˚C ( Figure 1F ).
We next determined the range of temperature-sensitivity of each * cdc15 * allele. All * cdc15 * alleles grew less well than wildtype cells at 36°C, with cdc15-140 showing the greatest temperature-sensitivity ( Figure 1G ).
In sum, we have provided further information on range of * cdc15 * mutants as well as initial characterization of previously uncharacterized * cdc15 * mutants that expand the repertoire of reagents which can be used to study cytokinesis and subcellular polarization of the early secretory pathway compartments in S. pombe .
Methods
S. pombe * methods
S. pombe strains were grown in yeast extract (YE) and standard S. pombe mating, sporulation, and tetrad dissection techniques were used to construct new strains (Forsburg and Rhind, 2006) . Spot assays were performed twice with reproducible results.
The marker reconstitution mutagenesis strategy described in (Tang et al., 2011) was used to isolate cdc15-22 . Briefly, a non-functional fragment of the selective marker his5 (containing the promoter and the N-terminal ORF fragment), together with the entire ura4 gene were inserted at the 3' end of the native * cdc15 * genomic locus, in the his5- * ura4 ^-^ * mutant genetic background. This strain was obtained by transforming the HpaI-linearized plasmid cdc15-pH5-C∆ and we denoted the locus as * cdc15-his5C∆::ura4 ^+^ * . In parallel, we performed error-prone PCR of the * cdc15 * genomic sequence fused to the 3′-UTR and the C-terminal part of * his5 ^+^ * from the plasmid cdc15-pH5-C+, at high Mg ^2+^ concentration. The mutagenized amplicon was transformed into * cdc15-his5C∆::ura4 ^+^ * locus and the homologous recombination within cdc15- and his5 -encoding sequences resulted in the replacement of the genomic * cdc15 * sequence with a library of PCR fragments. Such recombination events were selected based on the restoration of histidine prototrophy. We screened all resulting colonies for temperature-sensitive * cdc15 * alleles showing impaired recruitment of the tER to the cellular division site.
To tag the mutant Cdc15-22 protein with eGFP we used the 3'region * ^ cdc15 ^ * -5'region * ^ cdc15 ^ * -eGFP- * cdc15 * ^ORF^ -pJK210 plasmid that upon SnaBI linearization has homology arms to the 5'UTR and the 3'UTR of * cdc15 * gene, introducing the * eGFP- cdc15 * construct and the ura4+ cassette at the * cdc15 * locus.
Molecular biology methods
cdc15 * alleles were amplified using an oligonucleotide 123 bp upstream of the start site (GATAGGCAACGGTTGCTAGG) and 142 bp downstream of the stop codon (ACGAAGCTTAGACCATGACG) (Integrated DNA technologies). The PCR products were each sequenced by Plasmidsaurus (Eugene, OR) using Oxford Nanopore Technology with custom analysis and annotation.
The cdc15-pH5-C∆ plasmid was obtained through overlap-extension PCR using primers prSO1372(ccgctcgagACACTGATTACCCTCTTTTCTAG), prSO1373(ccactataaccaccattgttAACATTGTTAATAAAATAAAATAAAATATATTCC), prSO1374(tttattttattaacaatgttAACAATGGTGGTTATAGTGGATCG), prSO1375(CGGAATTCctaTACCGTCTGAACAAAGTTCG) and the amplicon was introduced into the pH5-C∆ plasmid (Tang et al., 2011) using XhoI and EcoRI restriction sites. The plasmid cdc15-pH5-C+ was obtained using prSO1376(cgcggatccGGATCGAACATTAAATTTAACG) and prSO1377(gctacggatatcCTATACCGTCTGAACAAAGTTCG) to amplify * cdc15 * locus and cloned into pH5-C+ backbone (Ta ng et al., 2011) using BamHI and EcoRV restriction sites.
For the N-terminal eGFP tagging of Cdc15-22 protein variant, we amplified the mutant allele ORF sequence with prSO712 (CGCGGATCCATGGAGGTTAATGGAGTCTCTCAATCTG) and prSO2584 (tccccgcggCTATACCGTCTGAACAAAGTTCGACGGG) and used to clone it downstream of eGFP in pJK210 vector using BamHI and SacII restriction sites. We amplified the 3'region of * cdc15 * using prSO2585 (ccatcgatACACTGATTACCCTCTTTTCTAGATG) and prSO2586 (tcgcccgggttgttctgcggagtacGTACCTTGATTTTAATTTACTAAAATCGG) and 5' region of * cdc15 * using prSO2587 (ctccgcagaactacGTATCTGCTAGCAAGTTTGAATGAAAATAGG) and prSO2588 (tcgcccgggAACCTCCATTTTGTTATTGAAAGAAC) and cloned them in front of the eGFP using ClaI, SnaBI and XmaI restriction sites to obtain the final plasmid 3'region * ^ cdc15 ^ * -5'region * ^ cdc15 ^ * -eGFP- * cdc15 * ^ORF^ -pJK210 which upon linearization with SnaBI targeted the native cdc15 locus.
Microscopy and image analysis
Strains for fixed-cell imaging experiments were grown at 25°C in YE and then shifted to 36°C for 4 hours. Cells were fixed with 70% ethanol for DAPI and methyl blue (MB) staining as described previously (Roberts-Galbraith et al., 2009). Images were acquired using a Zeiss Axio Observer inverted epifluorescence microscope with Zeiss 63× oil objective (1.46 NA) and captured using Zeiss ZEN 3.0 (Blue edition) software. A singular medial Z slice was obtained. All images were further processed using ImageJ (Schindelin et al., 2012) . All imaging experiments were repeated twice.
Live imaging of cdc15-22 strains was performed on epifluorescence microscopy setup using mercury lamp as an illumination source with appropriate sets of filters on a Zeiss Axiovert 200M microscope (Carl Zeiss, Inc.) using a Plan Apochromat 100X, 1.4 N.A. objective lens, CoolSnap camera (Photometrics, Tucson, AZ) and Uniblitz shutter driver (Photonics, Rochester, NY) under the control of Metamorph software package (Universal Imaging, Sunnyvale, CA). When required, imaging was performed at 36˚C using an objective heater system (Bioptechs, Butler, PA, USA). We acquired whole cell image stacks that consisted of 9 z-sections with 0.5 µm spacing for red and green fluorophores. The z-stack maximum or average projection images were obtained by ImageJ 1.46b software package (http://rsb.info.nih.gov/ij/; National Institutes of Health, Bethesda, MD, USA).
Protein methods
Cell pellets (15 OD) were snap-frozen in dry ice–ethanol baths. Pellets were resuspended in 1 ml and lysates were prepared by bead disruption (Fastprep cell homogenizer; MP Biomedicals) in NP-40 buffer (6 mM Na 2 HPO 4 , 4 mM NaH 2 PO 4 , 1% NP-40, 150 mM NaCl, 2 mM EDTA, 50 mM NaF, 4 mg/mL leupeptin, 0.1 mM Na 3 VO 4 ) with the addition of 1 mM PMSF (Sigma-Alrdrich; P7626), 2 mM benzamidine (Sigma-Alrdrich; B6506), and 0.5 mM diisopropyl fluorophosphate (Sigma-Aldrich; D0879-1G). Immunoprecipitations and lambda phosphatase collapse were performed as described (Bhattacharjee et al., 2023) . Briefly, lysates were incubated with anti- Cdc15 serum (Roberts-Galbraith et al., 2009) for 2 h at 4°C and then protein A sepharose beads (GE Healthcare; 17-5280-04) for 30 minutes. Beads were washed three times with NP-40 buffer and two times with phosphatase buffer (150 mM NaCl, 50 mM Hepes pH 7.4) before being split in two and added to either lambda phosphatase reaction or control. Lambda phosphatase collapse was performed according to manufacturer's protocol (New England Biolabs; P0753). Reactions were stopped by the addition of gel sample buffer.
Protein samples were resolved by SDS-PAGE and transferred to polyvinylidene fluoride (PVDF) membrane (Immobilon P, EMD Millipore). Anti- Cdc2 (PSTAIRE) mouse monoclonal antibody at 1:10,000 dilution (Sigma-Aldrich; P7962) or anti- Cdc15 (1-405) rabbit polyclonal antibody at 1:10,000 dilution (Roberts-Galbraith et al., 2009) were used as primary antibodies in immunoblotting. Secondary antibodies were conjugated to IRDye800 or IRDye680 (LI-COR Biosciences). Blotted proteins were detected via an Odyssey CLx instrument (LI-COR Biosciences).
Reagents
The strains used in this study and their genotypes are listed below.
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arasada R Pollard TD 201191 Distinct roles for F-BAR proteins Cdc 15p and Bzz 1p in actin polymerization at sites of endocytosis in fission yeast.Curr Biol 21170960-98221450145910.1016/j.cub.2011.07.04621885283 PMC 3350781 · doi ↗ · pubmed ↗
- 2Balasubramanian MK, Mc Collum D, Chang L, Wong KC, Naqvi NI, He X, Sazer S, Gould KL. 1998. Isolation and characterization of new fission yeast cytokinesis mutants. Genetics. 149: 1265-75. 1626.10.1093/genetics/149.3.1265 PMC 14602339649519 · doi ↗ · pubmed ↗
- 3Bhattacharjee R Hall AR Mangione MC Igarashi MG Roberts-Galbraith RH Chen JS Vavylonis D Gould KL 202327 Multiple polarity kinases inhibit phase separation of F-BAR protein Cdc 15 and antagonize cytokinetic ring assembly in fission yeast.Elife 1210.7554/e Life.8306236749320 PMC 9904764 · doi ↗ · pubmed ↗
- 4Bhattacharjee R Mangione MC Wos M Chen JS Snider CE Roberts-Galbraith RH Mc Donald NA Presti LL Martin SG Gould KL 2020226 DYRK kinase Pom 1 drives F-BAR protein Cdc 15 from the membrane to promote medial division.Mol Biol Cell 3191059-152491792910.1091/mbc.E 20-01-002632101481 PMC 7185970 · doi ↗ · pubmed ↗
- 5Carnahan RH, Gould KL. 2003. The PCH family protein, Cdc 15p, recruits two F-actin nucleation pathways to coordinate cytokinetic actin ring formation in Schizosaccharomyces pombe. J Cell Biol. 162: 851-62. 2140.10.1083/jcb.200305012 PMC 217282812939254 · doi ↗ · pubmed ↗
- 6Chang F, Woollard A, Nurse P. 1996. Isolation and characterization of fission yeast mutants defective in the assembly and placement of the contractile actin ring. J Cell Sci. 109: 131-42. 1490.10.1242/jcs.109.1.1318834798 · doi ↗ · pubmed ↗
- 7Cortés JC Pujol N Sato M Pinar M Ramos M Moreno B Osumi M Ribas JC Pérez P 201571 Cooperation between Paxillin-like Protein Pxl 1 and Glucan Synthase Bgs 1 Is Essential for Actomyosin Ring Stability and Septum Formation in Fission Yeast.P Lo S Genet 1171553-7390 e 1005358 e 100535810.1371/journal.pgen.100535826132084 PMC 4489101 · doi ↗ · pubmed ↗
- 8Fankhauser C, Reymond A, Cerutti L, Utzig S, Hofmann K, Simanis V. 1995. The S. pombe cdc 15 gene is a key element in the reorganization of F- actin at mitosis [published erratum appears in Cell 1997 Jun 27;89(7):1185]. Cell. 82: 435-44. 1494.10.1016/0092-8674(95)90432-87634333 · doi ↗ · pubmed ↗