sqt-3(sc63) is an alternative CRISPR/Cas9 co-conversion marker in Caenorhabditis elegans
Isa Özdemir, Florian A. Steiner

TL;DR
This paper introduces a new genetic marker for tracking genome edits in C. elegans using CRISPR/Cas9.
Contribution
The study introduces sqt-3(sc63) as a novel co-conversion marker on chromosome V for CRISPR/Cas9 genome editing in C. elegans.
Findings
sqt-3(sc63) is a functional co-conversion marker for CRISPR/Cas9 genome editing in C. elegans.
Its location on chromosome V makes it useful for editing targets on chromosome II.
The conserved nature of sqt-3 suggests potential use in other nematode species.
Abstract
The identification of genome-edited individuals by CRISPR/Cas9 in Caenorhabditis elegans often relies on the introduction of a second mutation with a visible phenotype. Popular targets for such co-conversion are dpy-10 and sqt-1 , both located on chromosome II. In this study, we introduce sqt-3 ( sc63 ) V as an alternative CRISPR/Cas9 co-conversion marker to facilitate the generation and detection of genome edits in C. elegans . Its location on chromosome V makes sqt-3 a desirable alternative to dpy-10 and sqt-1 when editing targets on chromosome II. Additionally, the conserved nature of sqt-3 provides a potential applicability in other nematode species.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
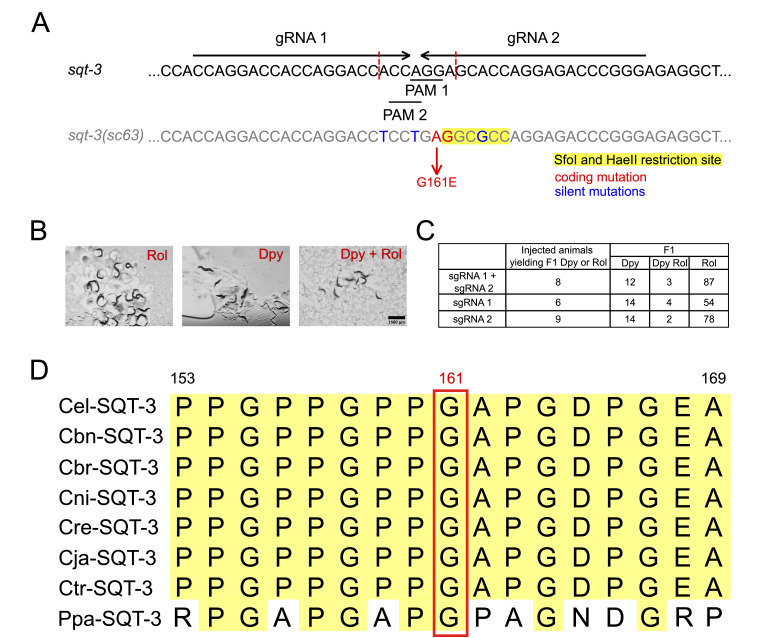 Figure 1
Figure 1|
|
|
|
|
|
pIO57 |
pRB1017 |
CCAGGACCACCAGGACCACC |
gRNA 1 |
|
pIO58 |
pRB1017 |
CCCGGGTCTCCTGGTGCTCC |
gRNA 2 |
|
|
|
|
|
oIO233 |
TCTTGCCAGGACCACCAGGACCACC |
To clone gRNA 1 (pIO57) |
|
oIO234 |
AAACGGTGGTCCTGGTGGTCCTGGC | |
|
oIO235 |
TCTTGCCCGGGTCTCCTGGTGCTCC |
To clone gRNA2 (pIO58) |
|
oIO236 |
AAACGGAGCACCAGGAGACCCGGGC | |
|
oIO237 |
CCACCATGCAAGCCATGCCCACAAGGACCACCAGGACCACCAGGACCTCCTGAAGCTCCAGGAGACCCGGGAGAGGCTGGAACCCCAGGACGCCCAGGGACCG |
ssDNA (Repair template) |
|
oIO263 |
GCCTACGGAGGACCAGAAG |
To genotype and sequence the
|
|
oIO264 |
CAAAACTCACTTTGGACAG |
|
|
|
|
|
pDD162 |
Cas9 expressing plasmid |
50 ng/µl |
|
pIO57 and pIO58 |
gRNA expressing plasmids |
25 ng/µl per gRNA when used together, 50ng/µl total |
|
Alternative: pIO57 or pIO58 |
gRNA expressing plasmids |
25 ng/µl per gRNA when used individually |
|
oIO237 |
Single stranded repair template |
500 nM |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCRISPR and Genetic Engineering · Genetics, Aging, and Longevity in Model Organisms · Innovation and Socioeconomic Development
Description
CRISPR/Cas9 genome editing has revolutionised genetic studies in biomedical research, including the model organism * C. elegans * [ (Friedland et al., 2013) ](https://www.zotero.org/google-docs/?vxMRan) . Cas9 is a double stranded DNA endonuclease that generates breaks upon the recognition of the target DNA locus through a protospacer adjacent motif (PAM) and a guide RNA (gRNA) that is complementary to the target sequence [ (Jinek et al., 2012; Liao et al., 2024) ](https://www.zotero.org/google-docs/?ywXVaA) . The Cas9-induced double-stranded breaks are repaired by either error prone nonhomologous end joining (NHEJ) or, less frequently, homologous recombination (HR) using a single- or double-stranded donor DNA (repair template). In genome-editing, these repair templates typically include the desired edits, insertions or deletions flanked by 30–50 nucleotide arms with homology to the Cas9 target site.
Although CRISPR/Cas9 is a highly versatile tool to generate targeted mutations with base-pair precision, the editing efficiency is variable, and obtaining the desired edits often requires extensive screening and genotyping effort. To reduce this effort, co-editing strategies were developed for * C. elegans * [ (Arribere et al., 2014) ](https://www.zotero.org/google-docs/?4aqN98) . The aim of co-editing is to target the gene of interest and a gene with a visible phenotype at the same time. Specific mutations in the * dpy-10 * or * sqt-1 * genes, both encoding members of the collagen gene family, have emerged as popular choices due to their visually distinguishable phenotypes. These marker mutations produce easily recognizable Roller (Rol) or Dumpy (Dpy) phenotypes in the F1 when one or both copies of DNA, respectively, have been edited. The presence of these phenotypes indicates successful genome editing in the individual at least at the co-edited locus, making it a candidate for also containing the edit at the desired target locus. However, both * dpy-10 * and * sqt-1 * are located in gene-rich regions on chromosome II, limiting their utility when targeting genetically linked loci on the same chromosome. In such cases, separating the desired genome edit from the genetically linked marker mutations can be labor-intensive and time-consuming. There are other co-conversion markers elsewhere in the genome, e.g. * unc-58 * on chromosome X. However, loss-of-function alleles of * unc-58 * are hard to distinguish from wildtype alleles (as is the case for another frequently used co-conversion marker, * rol-6 * ). This hinders the selection against CRISPR-edited loss-of-function mutations, thus limiting the utility of the locus in repeated rounds of CRISPR experiments [ (Arribere et al., 2014) ](https://www.zotero.org/google-docs/?PyeWpI) .
To address these limitations and improve the co-editing approach, we introduce * sqt-3 * as an alternative co-editing marker. * sqt-3 * encodes for a collagen and is located on chromosome V. The * sqt-3 ( sc63 ) * allele contains a missense mutation that replaces glycine (G) at position 161 with glutamic acid (E) ** ( Figure 1A ) ** . This G161E mutation results in a Squat (Sqt) phenotype, meaning heterozygotes display a Rol phenotype, while homozygotes exhibit a Dpy phenotype. The phenotypes are temperature-sensitive and more pronounced at 25 ^o^ C ** ( Figure 1B ) ** , while at 15 ^o^ C we observed a Long (Lon) phenotype [ (Kusch & Edgar, 1986) ](https://www.zotero.org/google-docs/?ngWLGY) . The phenotypes allow for phenotypic selection and counterselection of * sqt-3 ( sc63 ) * mutants in the F1 and F2 generations at 20 ^o^ C and 25 ^o^ C, similar to the * dpy-10 * and * sqt-1 * co-editing markers [ (Cox et al., 1980) ](https://www.zotero.org/google-docs/?0h8JnH) . Loss of function of * sqt-3 * results in a visible Dpy phenotype (as is the case for * sqt-1 * and * dpy-10 * ), and *mut/null * and null/null can be distinguished from mut /+ and +/+, respectively. This allows the selection against loss-of-function mutations, the presence of which might prevent subsequent rounds of co-conversion.
To introduce the * sqt-3 [G161E] * mutation in co-editing experiments, we designed a single-stranded DNA repair template and two gRNAs targeting the * sqt-3 * locus ** ( Figure 1A ) ** . These gRNAs exhibited editing efficiencies comparable to those reported for the widely used * dpy-10 ( cn64 ) * allele (Arribere et al., 2014) ** ( Figure 1C ) ** . The introduction of silent mutations within the repair template creates restriction sites for SfoI or HaeII , facilitating verification of the successful co-edit ** ( Figure 1A ) ** .
The amino acid sequence of SQT-3 around G161 is fully conserved in other * Caenorhabditis * species, including * C. brenneri * , * C. briggsae * , * C. nigoni * , * C. remanei * , * C. japonica * , and * C. tropicalis * , and the G161 residue is also present in P. pacificus ** ( Figure 1D ) ** . This conservation suggests that the * sqt-3 ( sc63 ) * mutation could serve as a versatile co-editing marker in other nematodes, expanding its utility beyond * C. elegans * .
In summary, * sqt-3 ( sc63 ) * V represents a valuable and convenient addition to the toolkit of CRISPR/Cas9 co-editing markers for * C. elegans * . Its different chromosomal location and temperature-dependent penetrance make it a practical alternative when * dpy-10 * or * sqt-1 * markers are unsuitable. The conserved amino acid sequence of SQT-3 also makes it potentially applicable in nematode species beyond * C. elegans * .
Methods
Worm Strains and Maintenance:
N2 (wildtype) strain was obtained from the * Caenorhabditis * Genetics Center, University of Minnesota. Strains were grown at 15°C, 20°C or 25 ^o^ C on NGM agar plates seeded with * E. coli WBStrain00041969 * OP50 .
CRISPR/Cas9 genome edits:
Cas9 and gRNAs were injected into the gonads of N2 adult hermaphrodites in the form of plasmids as described in [ (Arribere et al., 2014) ](https://www.zotero.org/google-docs/?u6skaH) . gRNAs were cloned as described in [ (Arribere et al., 2014) ](https://www.zotero.org/google-docs/?vkca3I) , and listed in Table 1 . The sequence of the repair oligonucleotide is given in Table 2 . All oligonucleotides were ordered from Microsynth. The Cas9 worm expression plasmid (pDD162, #47549) and empty vector for gRNA cloning (pRB1017, #59936) were obtained from Addgene [ (Arribere et al., 2014; Dickinson et al., 2013) ](https://www.zotero.org/google-docs/?gy19EG) . The composition of the injection mix is listed in Table 3 . A Leica DMIRE2 microscope and a Femtojet 4i (Eppendorf) were used for injections. F1 and F2 worms were screened for the desired phenotype under a standard dissecting microscope.
** Identification of the SQT-3 orthologues: **
The predicted SQT-3 protein sequences for * C. brenneri * , * C. briggsae * , * C. nigoni * , * C. remanei * , * C. japonica * , * C. tropicalis * , and P. pacificus were collected from Wormbase (WS294) based on nematode orthologs in the homology section.
The obtained sequences were aligned using Snapgene (version 8.0). Only the 8 aa upstream and 8 aa downstream of G161 in the alignment are shown in ** Figure 1D . **
Table 1. gRNA plasmids generated for this study
**: **
Table 2. Oligonucleotides used in this study.
**: **
** Table 3. Composition of the injection mix for the introduction of the * sqt-3 ( sc63 ) * *V * mutation. **
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arribere Joshua A Bell Ryan T Fu Becky X H Artiles Karen L Hartman Phil S Fire Andrew Z 2014826 Efficient Marker-Free Recovery of Custom Genetic Modifications with CRISPR/Cas 9 in Caenorhabditis elegans Genetics 19831943-263183784610.1534/genetics.114.16973025161212 PMC 4224173 · doi ↗ · pubmed ↗
- 2Cox George N Laufer John S Kusch Meredith Edgar Robert S 198061 GENETIC AND PHENOTYPIC CHARACTERIZATION OF ROLLER MUTANTS OF CAENORHABDITIS ELEGANS Genetics 9521943-263131733910.1093/genetics/95.2.31717249038 PMC 1214229 · doi ↗ · pubmed ↗
- 3Dickinson Daniel J Ward Jordan D Reiner David J Goldstein Bob 201391 Engineering the Caenorhabditis elegans genome using Cas 9-triggered homologous recombination Nature Methods 10101548-70911028103410.1038/nmeth.264123995389 PMC 3905680 · doi ↗ · pubmed ↗
- 4Friedland Ari E Tzur Yonatan B Esvelt Kevin M Colaiácovo Monica P Church George M Calarco John A 2013630 Heritable genome editing in C. elegans via a CRISPR-Cas 9 system Nature Methods 1081548-709174174310.1038/nmeth.253223817069 PMC 3822328 · doi ↗ · pubmed ↗
- 5Jinek Martin Chylinski Krzysztof Fonfara Ines Hauer Michael Doudna Jennifer A. Charpentier Emmanuelle 2012817 A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity Science 33760960036-807581682110.1126/science.122582922745249 PMC 6286148 · doi ↗ · pubmed ↗
- 6Kusch Meredith Edgar R S 198671 GENETIC STUDIES OF UNUSUAL LOCI THAT AFFECT BODY SHAPE OF THE NEMATODE CAENORHABDITIS ELEGANS AND MAY CODE FOR CUTICLE STRUCTURAL PROTEINS Genetics 11331943-263162163910.1093/genetics/113.3.6213732788 PMC 1202859 · doi ↗ · pubmed ↗
- 7Liao Hongyu Wu Jiahao Van Dusen Nathan J. Li Yifei Zheng Yanjiang 2024121 CRISPR-Cas 9-mediated homology-directed repair for precise gene editing Molecular Therapy - Nucleic Acids 3542162-253110234410234410.1016/j.omtn.2024.10234439494147 PMC 11531618 · doi ↗ · pubmed ↗