At5g63290 does not encode coproporphyrinogen III oxidase
Wenjuan Ji, Huijuan Wang, Ning An, Xuan Zhou, Bingxiao Wen, Bernhard Grimm, Zhenhua Liu

TL;DR
This paper shows that the gene At5g63290 in Arabidopsis does not perform a key step in tetrapyrrole production, changing the known pathway.
Contribution
The study corrects the assumed function of At5g63290 in the tetrapyrrole biosynthesis pathway.
Findings
At5g63290 does not catalyze the decarboxylation of coproporphyrinogen III.
The tetrapyrrole biosynthesis pathway in Arabidopsis needs to be revised.
Abstract
At5g63290 is not responsible for the decarboxylation of coproporphyrinogen III, prompting a revision of the tetrapyrrole biosynthesis pathway in Arabidopsis thaliana.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
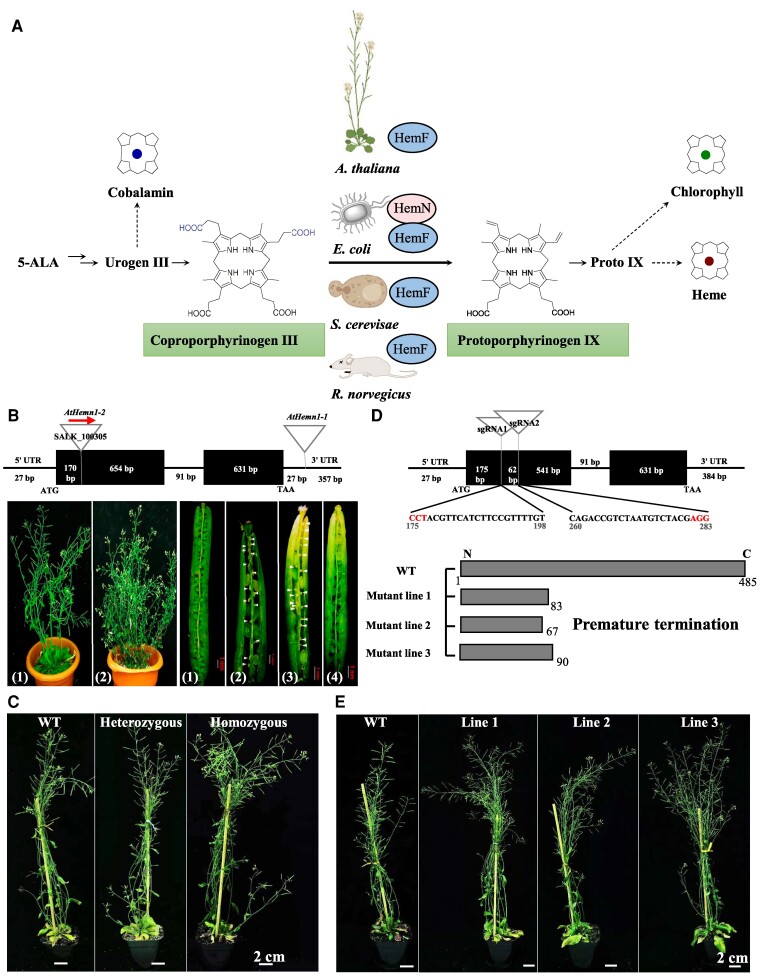 Figure 1
Figure 1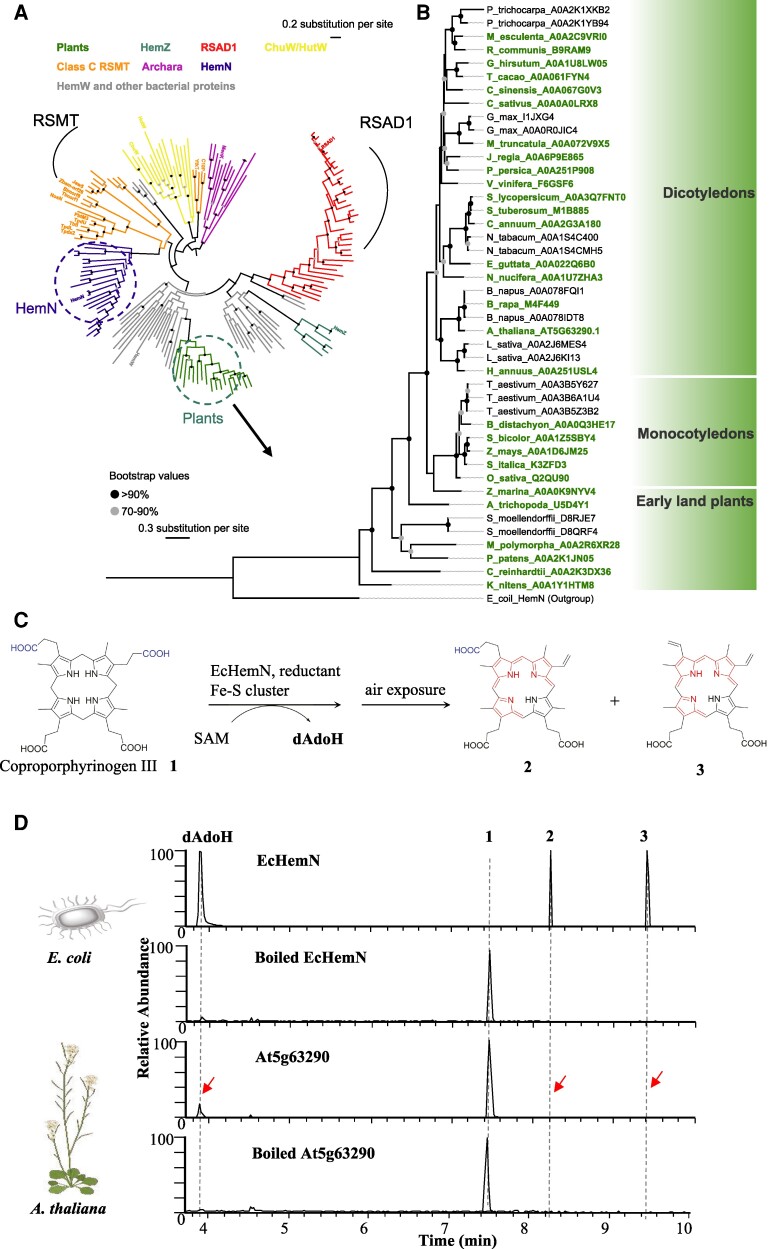 Figure 2
Figure 2- —National Natural Science Foundation of China for Youths
- —National Key R&D Program of China10.13039/501100012166
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhotosynthetic Processes and Mechanisms · Porphyrin Metabolism and Disorders · Porphyrin and Phthalocyanine Chemistry
Dear Editor,
Tetrapyrrole synthesis plays essential roles in the primary metabolism across all life kingdoms. The main end products of the biosynthetic pathway are chlorophyll and heme, but photosynthetic organisms also use several other tetrapyrroles, such as siroheme and phytochromobilin (Belot et al. 2024; Gao et al. 2024). It has been demonstrated that the synthesis of chlorophyll and heme includes the oxidative decarboxylation of coproporphyrinogen III to produce protoporphyrinogen IX (Fig. 1A). While an oxygen-dependent coproporphyrinogen III oxidase (also named HemF) has been characterized in several bacteria and across Eukaryotes, including thale cress (Arabidopsis thaliana) (Ishikawa et al. 2001), mice (Rattus norvegicus) (Mori et al. 2013), yeast (Saccharomyces cerevisiae) (Phillips et al. 2004), and Escherichia coli (Breckau et al. 2003), the oxygen-independent coproporphyrinogen III oxidase (also named HemN) has been thoroughly identified only in E. coli, as demonstrated by others and our own work (Fig. 1A) (Layer et al. 2002; Ji et al. 2020). The HemN requires S-adenosyl-l-methionine (SAM), a reductant, and additional cytoplasmatic components for catalysis and carries/contains a Fe–S cluster (Layer et al. 2002). Whether an oxygen-independent CPO exists in Eukaryotes, however, has not been clearly established. Recently, Pratibha et al. (2017) identified a potential HemN-like enzyme (At5g63290) from the model plant Arabidopsis. In their findings, the homozygous mutant line, designated Athemn1, was lethal, and the heterozygous progenies of the mutant showed defects in gametophyte development (Fig. 1B). However, when analyzing the same T-DNA insertion line used by Pratibha et al. and in addition to 3 independent mutant lines generated with CRISPR/Cas9, no obvious growth and developmental phenotypes were detectable for the HemN-deficient mutant lines (Fig. 1, C to E; Supplementary material and Methods). By analyzing the phylogeny of HemN-like proteins across all kingdoms (Cheng et al. 2022), our results indicate that At5g63290 is not grouped with HemN, but rather with the HemW clade (Fig. 2A; Supplementary material and Methods). Initial biochemical analysis of Lactococcus lactis revealed that HemW is a heme chaperone in heme transfer (Haskamp et al. 2018). We further purified E. coli HemN and At5g63290 and performed in vitro enzyme assays. While we clearly observed functional CPO activity of E. coli HemN, we failed to detect decarboxylation activity of uroporphyrinogen by the protein encoded by At5g63290, although it has SAM cleavage activity (Fig. 2D). Lastly, Pratibha et al. claimed that At5g63290 encodes a mitochondrial protein, but when we transiently expressed the protein in Nicotiana benthamiana, the dominant signal appeared in the chloroplast (Supplementary Fig. S1 and Material and Methods).
In view of the essential role of tetrapyrrole synthesis in plants, e.g. in respiration and photosynthesis, one would expect a severe growth phenotype of a mutant lacking a single gene for a specific step of tetrapyrrole biosynthesis (Tanaka and Tanaka 2007; Honkanen et al. 2016). Pratibha et al. (2017) presented the 2 homozygous T-DNA insertion mutants Athemn1-1 (with a T-DNA insertion in the 3′ UTR of At5g63290) and Athemn1-2 (with a T-DNA insertion in the first exon of At5g63290) as lethal mutants, while their heterozygous progenies contained shorter siliques, fewer ovules, and seed sterility under long-day conditions (Fig. 1B). We also obtained and conducted phenotyping of the same Athemn1-2 T-DNA mutant (SALK_100305) (Supplementary Fig. S2, Material and Methods, and Table S1). In our experiments, both heterozygous and homozygous Athemn1-2 mutants were fertile (Fig. 1C). To verify this analysis, we used CRISPR/Cas9-mediated mutagenesis and generated 3 independent knockout (KO) lines targeting the first exon of At5g63290 (Fig. 1D). Also, none of these KO mutants were lethal, and both the heterozygotes and homozygotes showed no noticeable differences in growth or development compared to the wild-type control (Fig. 1E). Recent publications confirm that all plant tetrapyrrole biosynthesis is exclusively localized in plastids. Earlier findings suggesting a second heme synthesis pathway with the final enzymes protoporphyrinogen oxidase and ferrochelatase in mitochondria have been refuted (Hedtke et al. 2023; Wittmann et al. 2024). Therefore, there seems to be no need for mitochondrial CPO in plants.
Phylogeny and functional analysis further supported our findings. Cheng et al. (2022) conducted a Bayesian-based phylogenic analysis of HemN-like proteins across various life kingdoms. Their findings suggest that putative plant HemN-like proteins do not cluster with the bacterial HemN and are instead arranged separately from other functional clades, such as class C radical SAM methyltransferase (RSMT), with members mostly function as methyltransferases (Mathew et al. 2022). This result was also confirmed by our phylogenetic analysis with maximum probability. The At5g63290 was not grouped with HemN but instead clustered with the HemW clade in bacterial and the radical SAM domain-containing 1 (RSAD1) clade in animals (Fig. 2A). The RASD1 was suggested to have a heme chaperone activity similar to HemW (Haskamp et al. 2018). At5g63290 exists as a broadly distributed single-copy gene across land plants (Fig. 2B). So far, functions of HemW are not well understood. It has been reported that the E. coli HemW shows no CPO activity either in vitro or in vivo (Haskamp et al. 2018). Emerging evidence suggests that HemW acts as a heme chaperone rather than as a biosynthetic protein (Haskamp et al. 2018). Motif analysis revealed that At5g63290 contains a HNXXYW domain, which is known to be involved in heme-binding activity of HemW (Supplementary Fig. S3) (Sharma et al. 2021).
The At5g63290 also contained a CxxxCxxC motif characteristic of the radical SAM superfamily. This motif utilizes 3 Cys residues to bind a [4Fe-4S] cluster, coordinating with SAM to reductively cleave its carbon–sulfur bond. This process generates a highly reactive 5′-deoxyadenosyl (dAdo) radical, which initiates a variety of reactions (Broderick et al. 2014; Nicolet 2020). When coexpressing the iron–sulfur cluster biogenesis pathway along with At5g63290 in E. coli, the protein was found to be soluble (Supplementary Fig. S4 and Material and Methods). This suggests that the Fe–S cluster is necessary for proper protein folding.
To investigate the CPO activity and elusive role of the putative radical SAM enzyme, purified recombinant protein was incubated with the substrate coproporphyrinogen III in the presence of dithionite and SAM under anaerobic conditions. The mixture was then analyzed using ultraperformance liquid chromatography high-resolution MS (UPLC-HRMS). Both EcHemN and At5g63290 displayed SAM cleavage activity, as indicated by the presence of 5-deoxyadenosine (dAdoH) in the extracts, which is characteristic of the superfamily of radical SAMs (Fig. 2, C and D). However, the m/z 609.27, corresponding to the oxidized harderoporphyrinogen (2), and the m/z 563.26, corresponding to the oxidized protoporphyrinogen IX (3), were only detected in EcHemN assay, not with At5g63290. Compounds 2 and 3 were the oxidized forms of HemN products, resulting from exposure to air before UPLC-HRMS analysis (Fig. 2, C and D).
Thus, based on our data, we conclude that At5g63290 does not encode a CPO. But the biochemical function of this single-copy gene remains to be characterized further. We do not exclude that exposure of the T-DNA and CRISPR KO lines to severe stress conditions may exhibit visible mutant phenotypes, which could provide insights into the biofunctions of this gene.
Supplementary Material
kiaf046_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Belot A , Puy H, Hamza I, Bonkovsky HL. Update on heme biosynthesis, tissue-specific regulation, heme transport, relation to iron metabolism and cellular energy. Liver Int. 2024:44(9):2235–2250. 10.1111/liv.1596538888238 PMC 11625177 · doi ↗ · pubmed ↗
- 2Breckau D , Mahlitz E, Sauerwald A, Layer G, Jahn D. Oxygen-dependent coproporphyrinogen III oxidase (Hem F) from Escherichia coli is stimulated by manganese*. J Biol Chem. 2003:278(47):46625–46631. 10.1074/jbc.M 30855320012975365 · doi ↗ · pubmed ↗
- 3Broderick JB , Duffus BR, Duschene KS, Shepard EM. Radical S-adenosylmethionine enzymes. Chem Rev. 2014:114(8):4229–4317. 10.1021/cr 400470924476342 PMC 4002137 · doi ↗ · pubmed ↗
- 4Cheng J , Liu WQ, Zhu X, Zhang Q. Functional diversity of Hem N-like proteins. ACS Bio Med Chem Au. 2022:2(2):109–119. 10.1021/acsbiomedchemau.1c 00058 PMC 1011471837101745 · doi ↗ · pubmed ↗
- 5Gao F , Guo J, Shen Y. Advances from chlorophyll biosynthesis to photosynthetic adaptation, evolution and signaling. Plant Stress. 2024:12:100470. 10.1016/j.stress.2024.100470 · doi ↗
- 6Haskamp V , Karrie S, Mingers T, Barthels S, Alberge F, Magalon A, Müller K, Bill E, Lubitz W, Kleeberg K, et al The radical SAM protein Hem W is a heme chaperone. J Biol Chem. 2018:293(7):2558–2572. 10.1074/jbc.RA 117.00022929282292 PMC 5818191 · doi ↗ · pubmed ↗
- 7Hedtke B , Strätker SM, Pulido ACC, Grimm B. Two isoforms of Arabidopsis protoporphyrinogen oxidase localize in different plastidal membranes. Plant Physiol. 2023:192(2):871–885. 10.1093/plphys/kiad 10736806676 PMC 10231370 · doi ↗ · pubmed ↗
- 8Honkanen S , Jones VAS, Morieri G, Champion C, Hetherington A, Kelly S, Proust H, Saint-Marcoux D, Helen P, Dolan L. The Mechanism forming the cell surface of tip-growing rooting cells is conserved among land plants. Curr Biol. 2016:26(23):3238–3244. 10.1016/j.cub.2016.09.06227866889 PMC 5154754 · doi ↗ · pubmed ↗