The regulation of triglyceride storage by Acsx4 and Acsx5 in Drosophila fat tissue
Erick Astacio, Justin R. DiAngelo

TL;DR
This study explores how two genes, Acsx4 and Acsx5, regulate triglyceride levels in fruit fly fat tissue.
Contribution
The study identifies Acsx4 and Acsx5 as regulators of lipid metabolism in Drosophila adipose tissue.
Findings
Reducing Acsx4 and Acsx5 in fly fat tissue leads to increased triglyceride accumulation.
Acsx4 and Acsx5 may regulate lipid breakdown, compensating for triglyceride buildup when 9G8 is reduced.
9G8 downregulation in adipose tissue upregulates Acsx4 and Acsx5.
Abstract
The production of energy is one of the most fundamental requirements for organismal survival. Decreasing expression of Drosophila 9G8 , an mRNA splicing protein, specifically in adipose tissue results in triglyceride accumulation. Decreasing 9G8 in adipose also results in upregulation of the acyl-CoA synthetases Acsx4 and Acsx5 ; however, the functions of these genes in regulating lipid metabolism is not fully understood. Here, we decreased Acsx4 and Acsx5 in fly adipose tissue and this resulted in high triglycerides. This suggests that these genes regulate lipid breakdown, and their upregulation is perhaps compensating for the triglyceride accumulation observed when 9G8 levels are decreased.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
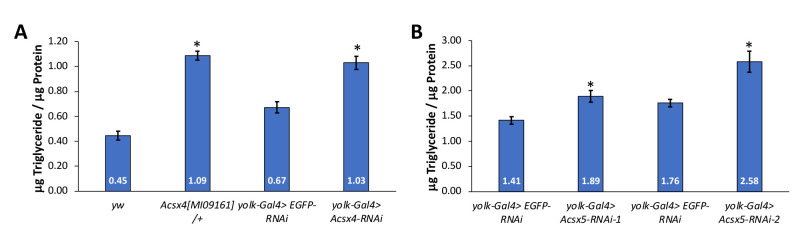 Figure 1
Figure 1|
|
|
|
|
|
|
BL#1495 |
|
|
|
BL#51390 |
|
|
|
BL#41555 |
|
|
|
BL#55267 |
|
|
|
BL#57155 |
|
|
|
BL#62912 |
|
|
|
BL#58814 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDiet, Metabolism, and Disease · Lipid metabolism and biosynthesis · Peroxisome Proliferator-Activated Receptors
Description
Alteration of the pathways that produce energy such as the breakdown of carbohydrates and fats can result in chronic disorders such as obesity and type 2 diabetes (Hotamisligil, 2006) . These disorders have been classified as a major global crisis due to the comorbidities associated with them, so a better understanding of the metabolic pathways that are altered in obesity and type 2 diabetes will hopefully help with the prevention of these diseases (Verma and Hussain, 2017) . Excess storage of triglycerides is observed in both obesity and type 2 diabetes; however, all the genes responsible for the regulation of nutrients stored as triglycerides is not known.
To identify and better understand the genes important for triglyceride storage, the fruit fly, Drosophila melanogaster , has been selected as a useful model organism to meet this objective. Flies are an excellent model system to study fat metabolism since they share similar genes to humans and have analogous organs to humans such as the fat body which functions like human liver and adipose tissue (Musselman and Kuhnlein, 2018) . To identify novel genes responsible for lipid storage, genome-wide RNA interference (RNAi) screening was performed in *Drosophila * cells, and many different types of genes were identified (Guo et al., 2008; Beller et al., 2008) . Splicing factors, proteins involved in the processing of mRNAs to generate mature transcripts, were among the genes identified in these screens (in each of the two RNAi screens described above, approximately 7% of the genes that resulted in fewer/smaller lipid droplets when knocked down were involved in mRNA splicing) and our lab has characterized several splicing factors that act in fat tissue to regulate triglyceride storage. Approximately 10% of the splicing factors from the Guo and Beller screens were SR proteins, a family of proteins that function to identify intron/exon borders (Long and Caceres, 2009) , and two of the eight SR proteins in the *Drosophila * genome were identified in these screens (Guo et al. 2008; Beller et al., 2008) . We have specifically focused on SR proteins in regulating metabolic homeostasis in Drosophila . For example, when the expression of one SR protein, 9G8 , was decreased specifically in fat tissue, the flies displayed a high triglyceride storage phenotype (Gingras et al., 2014) . To identify the genes regulated by 9G8 to result in this accumulation of triglycerides, RNA sequencing was performed on fat tissue with 9G8 levels decreased (Weidman et al., 2022) . Interestingly, two novel genes predicted to be acyl-CoA synthetases (ACS) (named * CG4830 * and * CG11453 * ) were identified as being upregulated when 9G8 expression was decreased (Weidman et al., 2022) . However, whether these ACS genes are responsible for promoting triglyceride storage when 9G8 is decreased is not known.
Both * CG11453 * and * CG4830 * are members of a group of ACS genes that are found specifically in worms and flies (Watkins et al, 2007) . Since the substrates of these ACS enzymes are not characterized, this group of ACS genes has been referred to as *Acsx * (Watkins et al, 2007) and * CG11453 * has already been named Acsx4 . Since * CG4830 * has not been characterized, but is a member of this *Acsx * gene group, we have decided to name this gene Acsx5 . ACS enzymes are responsible for adding CoA groups to fatty acids and these fatty acids can either be targeted for breakdown via beta-oxidation or they can be esterified with glycerol to make triglycerides (Watkins, 1997) . However, whether Acsx4 and Acsx5 function in *Drosophila * to promote fatty acid oxidation or triglyceride esterification is not known.
To characterize the role of Acsx4 in regulating lipid metabolism in Drosophila , triglycerides were measured in flies heterozygous for a transposon insertion in * Acsx4 (Acsx4 ^[MI09161]^ /+) * . In these Acsx4 mutant flies, an increase in triglyceride was observed (Fig 1A). To determine if the increased triglycerides in *Acsx4 * mutants results from *Acsx4 * function in the fat body, RNAi towards *Acsx4 * was induced specifically in the adult female fat body *. * The knockdown of *Acsx4 * via RNAi in adipose tissue showed similar increases in triglyceride observed in the Acsx4 mutants (Fig 1A). To assess the function of Acsx5 in regulating lipid storage, RNAi towards *Acsx5 * was induced specifically in the fat body using two independent RNAi lines. When *Acsx5 * was decreased using both RNAi lines, triglyceride accumulated, similar to the phenotypes observed with Acsx4 (Fig 1B). Together, these data suggest that both *Acsx4 * and
- Acsx5 * limit lipid storage, perhaps by promoting the addition of CoA groups onto fatty acids to target them for beta-oxidation in the mitochondria. In addition, the upregulation of these genes in adipose-specific *9G8-RNAi * flies is probably compensating for the increased triglycerides observed in these flies as the fold increase in triglycerides in fat body-specific *9G8-RNAi * flies is much higher than the fold increases in triglycerides observed in fat body-specific knockdown of *Ascx4 * or *Acsx5 * (Gingras et al., 2014; Fig 1).
Two additional acyl-CoA synthetases, pudgy and bubblegum , have been identified in Drosophila to regulate triglyceride metabolism. Overexpressing pudgy in the fat body in vivo results in blunted organismal fat levels, while decreasing *pudgy * increases triglyceride storage (Xu et al., 2012) . Another acyl-CoA synthetase in flies, bubblegum , was found to exhibit elevated levels of very long chain fatty acids when its expression was decreased (Min and Benzer, 1999) . These phenotypes in previously characterized *Drosophila * acyl-CoA synthetases are consistent with the phenotype observed when Acsx4 and Acsx5 were knocked in the fat body of female flies, suggesting that *Drosophila * have multiple functional acyl-CoA synthetases that regulate lipid breakdown. Additional experimentation is needed to determine the regulation of these enzymes in different tissues and throughout different stages in *Drosophila * development to regulate metabolic homeostasis.
Acyl-CoA synthetases are also well conserved in mammals. Five subfamilies of mammalian acyl CoA synthetases have been identified in which the length of the carbon chain of the fatty acid species defines the substrate specificity for these enzymes (Soupene and Kuypers, 2008) . The function of one of these families, long-chain acyl-CoA synthetase-1 (ACSL1), has been studied in both mouse and rat adipose tissue. When ACSL1 was knocked out in the adipose tissue of mice, triglycerides accumulated, and a 50-90% lower beta oxidation rate was observed when compared to the controls (Ellis et al., 2010) . This suggests a role for ACSL1 in regulating lipid breakdown which is like what we see with Acsx4 and Acsx5 in Drosophila . Moreover, ACSL1 mRNA and protein levels have also been shown to be regulated by fasting and refeeding in rats (Mashek et al., 2006) , suggesting that perhaps diet regulates Acsx4 and Acsx5 expression in *Drosophila * fat tissue. Future experiments are necessary to address any regulation of Acsx4 and Acsx5 by different dietary conditions in Drosophila .
Methods
** Fly genetics **
Experimental crosses of virgin GAL4 females with UAS males were performed at 25°C in a 12h:12h light: dark cycle. * Acsx4 ^[MI09161]^ /TM3, Sb[1] Ser[1] * flies were crossed to *yw * background controls to generate * Acsx4 ^[MI09161]^ /+ * flies used in this study. Flies were grown on sugar-yeast-cornmeal food (9g *Drosophila * agar (Genesee Scientific), 100mL Karo Lite Corn Syrup, 65g cornmeal, 40g sucrose, and 25g whole yeast in 1.25L water). Flybase was used to find background information and fly stocks available for Acsx4 and Acsx5 (Öztürk-Çolak et al., 2024).
** Triglyceride and Protein Measurements **
Throughout this study one-week old female flies were used; females were mated and aged with male flies. Two whole flies were homogenized in lysis buffer (140 mM NaCl, 50 mM Tris-HCl, pH 7.5, 0.1% Triton-X, and 1X protease inhibitor (ThermoFisher, Waltham, MA, USA)). Triglyceride and protein were measured using the Infinity Triglyceride Reagent kit (ThermoFisher, Waltham, MA, USA)), and Pierce BCA Protein Assay kit (ThermoFisher, Waltham, MA, USA), respectively, according to manufacturer's instructions and as previously described (Bennick et al., 2019) .
** Statistics **
The mean triglyceride/protein results were analyzed comparing values between * Acsx4 ^[MI09161]^ /+ * mutants and *yw * controls, and the yolk-Gal4>Acsx4-RNAi , and yolk-Gal4>Acsx5-RNAi flies and respective yolk-Gal4>EGFP-RNAi control flies via an unpaired, two-sample t-test as calculated in Microsoft Excel. A p<0.05 was used to determine statistical significance.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Beller M Sztalryd C Southall N Bell M Jäckle H Auld DS Oliver B 20081125 COPI complex is a regulator of lipid homeostasis.P Lo S Biol 6111544-9173 e 292e 29210.1371/journal.pbio.006029219067489 PMC 2586367 · doi ↗ · pubmed ↗
- 2Bennick RA Nagengast AA Di Angelo JR 201972 The SR proteins SF 2 and RBP 1 regulate triglyceride storage in the fat body of Drosophila.Biochem Biophys Res Commun 51630006-291X 92893310.1016/j.bbrc.2019.06.15131277943 · doi ↗ · pubmed ↗
- 3Ellis JM Li LO Wu PC Koves TR Ilkayeva O Stevens RD Watkins SM Muoio DM Coleman RA 201077 Adipose acyl-Co A synthetase-1 directs fatty acids toward beta-oxidation and is required for cold thermogenesis.Cell Metab 1211550-4131536410.1016/j.cmet.2010.05.01220620995 PMC 2910420 · doi ↗ · pubmed ↗
- 4Gingras RM Warren ME Nagengast AA Diangelo JR 20131211 The control of lipid metabolism by m RNA splicing in Drosophila.Biochem Biophys Res Commun 44320006-291X 67267610.1016/j.bbrc.2013.12.02724333419 PMC 4412844 · doi ↗ · pubmed ↗
- 5Guo Y Walther TC Rao M Stuurman N Goshima G Terayama K Wong JS Vale RD Walter P Farese RV 2008413 Functional genomic screen reveals genes involved in lipid-droplet formation and utilization.Nature 45371950028-083665766110.1038/nature 0692818408709 PMC 2734507 · doi ↗ · pubmed ↗
- 6Hotamisligil GS 20061214 Inflammation and metabolic disorders.Nature 44471210028-083686086710.1038/nature 0548517167474 · doi ↗ · pubmed ↗
- 7Long JC Caceres JF 200911 The SR protein family of splicing factors: master regulators of gene expression.Biochem J 41710264-6021152710.1042/BJ 2008150119061484 · doi ↗ · pubmed ↗
- 8Mashek DG Li LO Coleman RA 2006613 Rat long-chain acyl-Co A synthetase m RNA, protein, and activity vary in tissue distribution and in response to diet.J Lipid Res 4790022-22752004201010.1194/jlr.M 600150-JLR 20016772660 · doi ↗ · pubmed ↗