Exogenous interactome analysis of bovine viral diarrhea virus-host using network based-approach and identification of hub genes and important pathways involved in virus pathogenesis
Seyedeh Elham Rezatofighi

TL;DR
This study maps interactions between bovine viral diarrhea virus and host proteins to identify key genes and pathways involved in the virus's disease process.
Contribution
The paper presents a network-based analysis of BVDV-host interactions and identifies novel hub genes and pathways for potential therapeutic targets.
Findings
BVDV-host interactions mainly target translation, protein synthesis, and cellular metabolism pathways.
Npro is the BVDV protein with the most interactions with host proteins.
CD46, EEF-2, and TXN are identified as hub genes in the interaction network.
Abstract
Bovine viral diarrhea (BVD) is one of the most important diseases in livestock, caused by BVD virus (BVDV). During the pathogenesis of the virus, many interactions occur between host and viral proteins. Studying these interactions can help better understand the pathogenesis of the virus, identify putative functional proteins, and find new treatment and prevention strategies. To this aim, a BVDV-host protein-protein interaction (PPI) network map was constructed using Cytoscape and analyzed with cytoHubba, Kyoto Encyclopedia of Genes and Genomics (KEGG), Gene Ontology (GO), and Protein Analysis Through Evolutionary Relationships (PANTHER). Npro with 125 connections had the greatest number of interactions with host proteins. CD46, EEF-2, and TXN genes were detected as hub genes using different ranking algorithms in cytoHubba. BVDV interactions with its host mainly focus on targeting…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
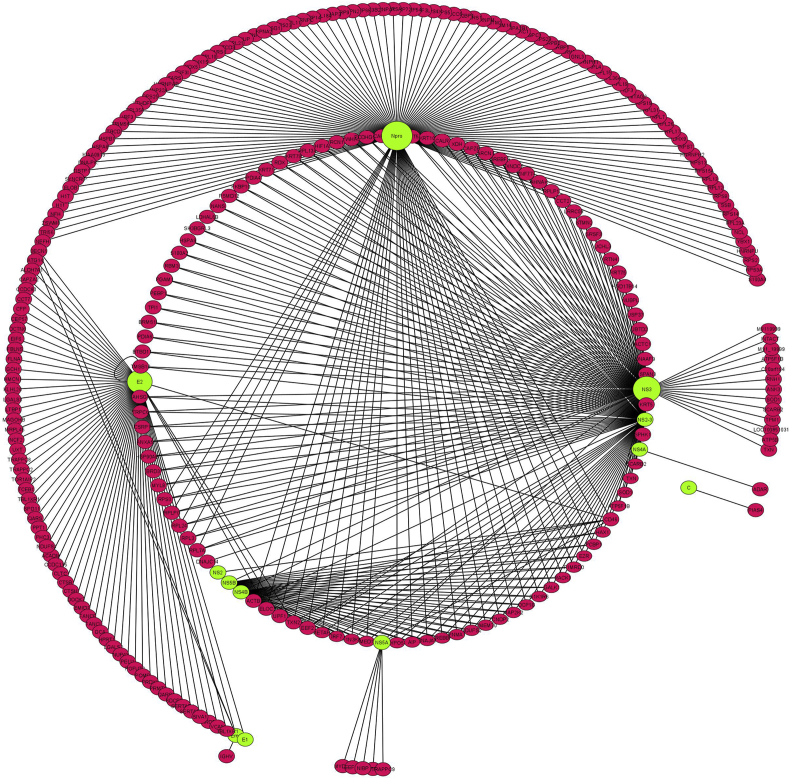 Figure 1
Figure 1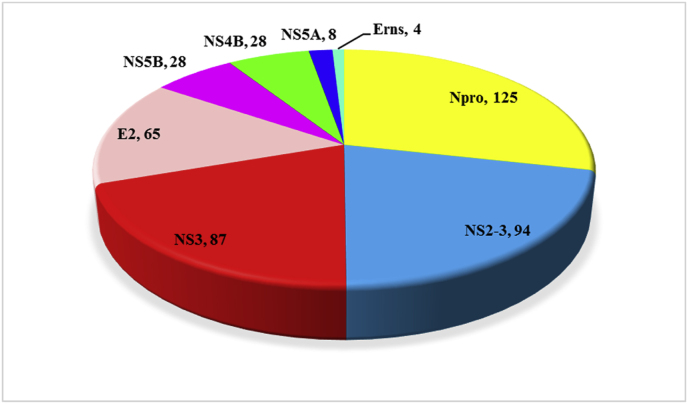 Figure 2
Figure 2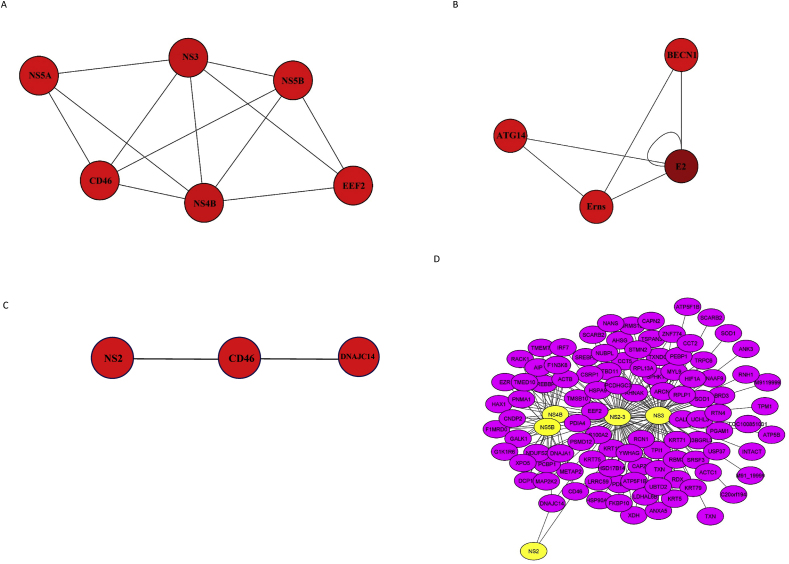 Figure 3
Figure 3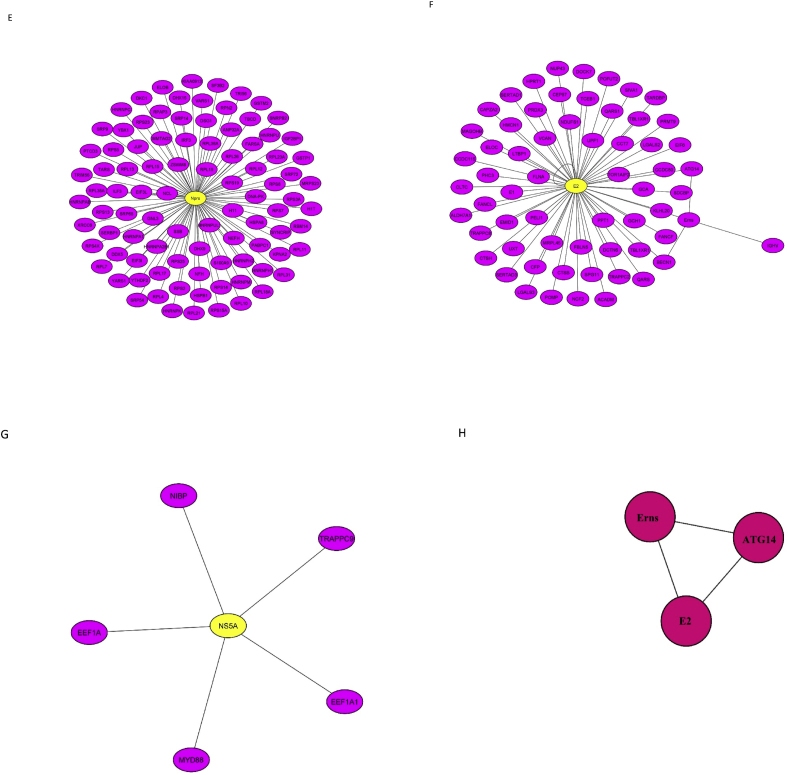 Figure 4
Figure 4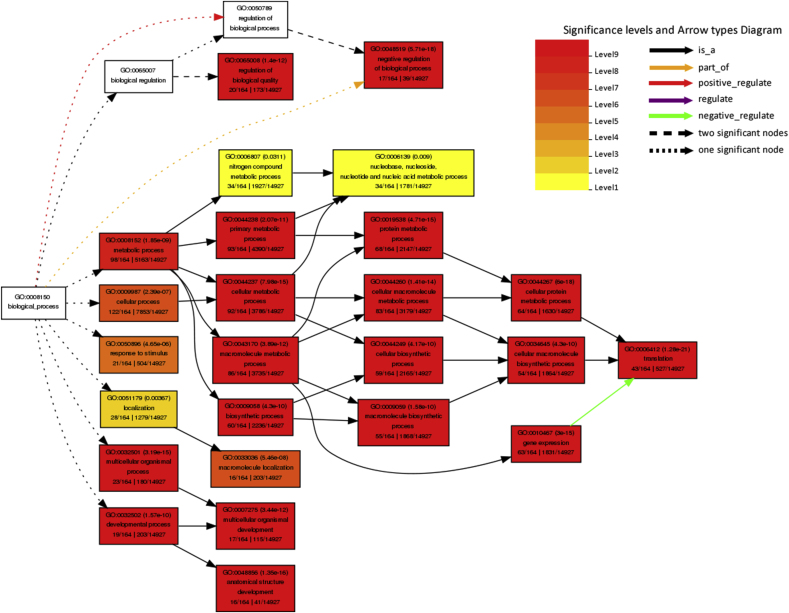 Figure 5
Figure 5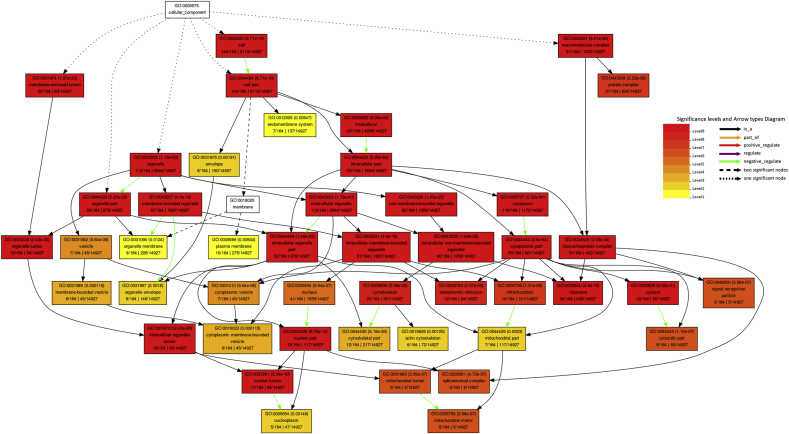 Figure 6
Figure 6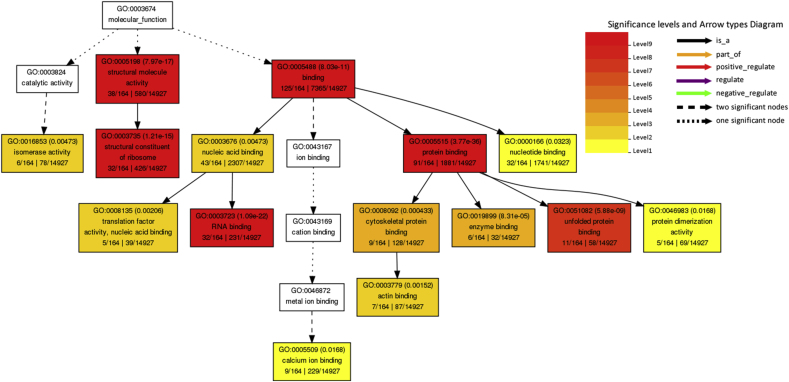 Figure 7
Figure 7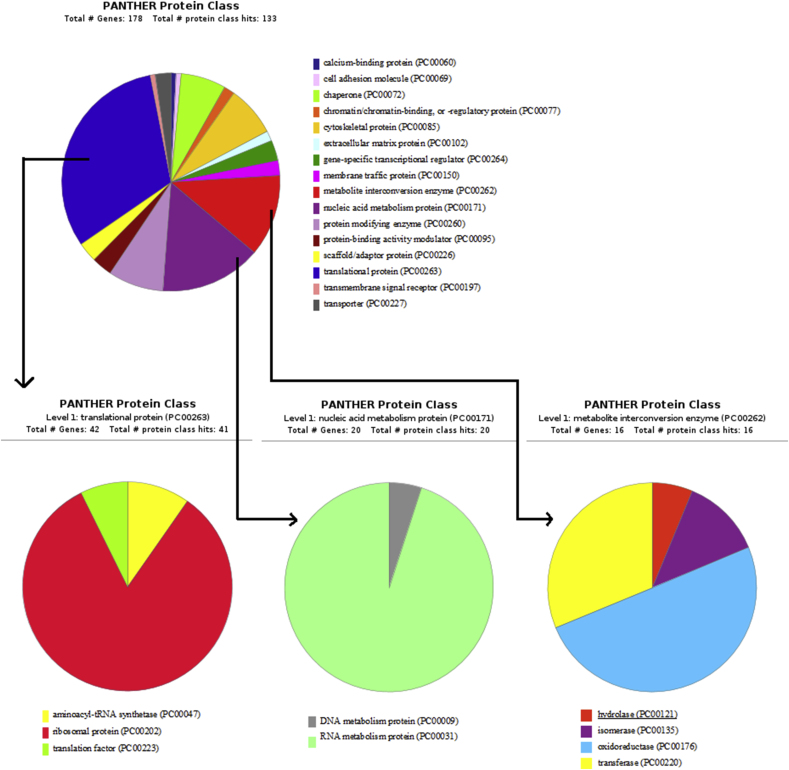 Figure 8
Figure 8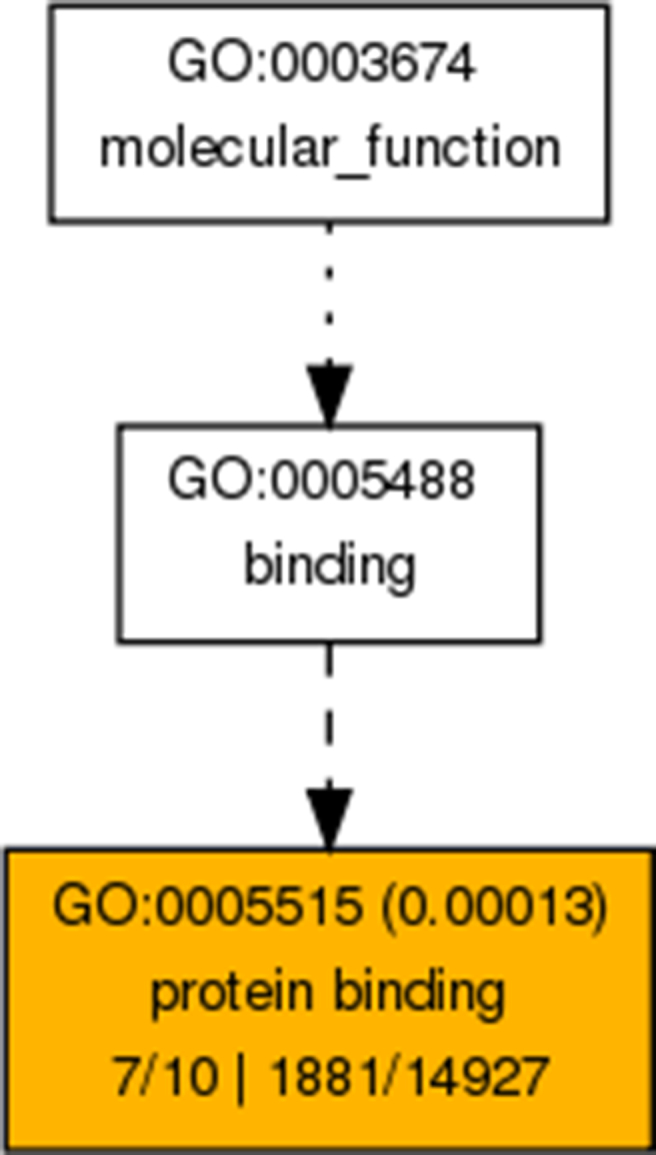 Figure 9
Figure 9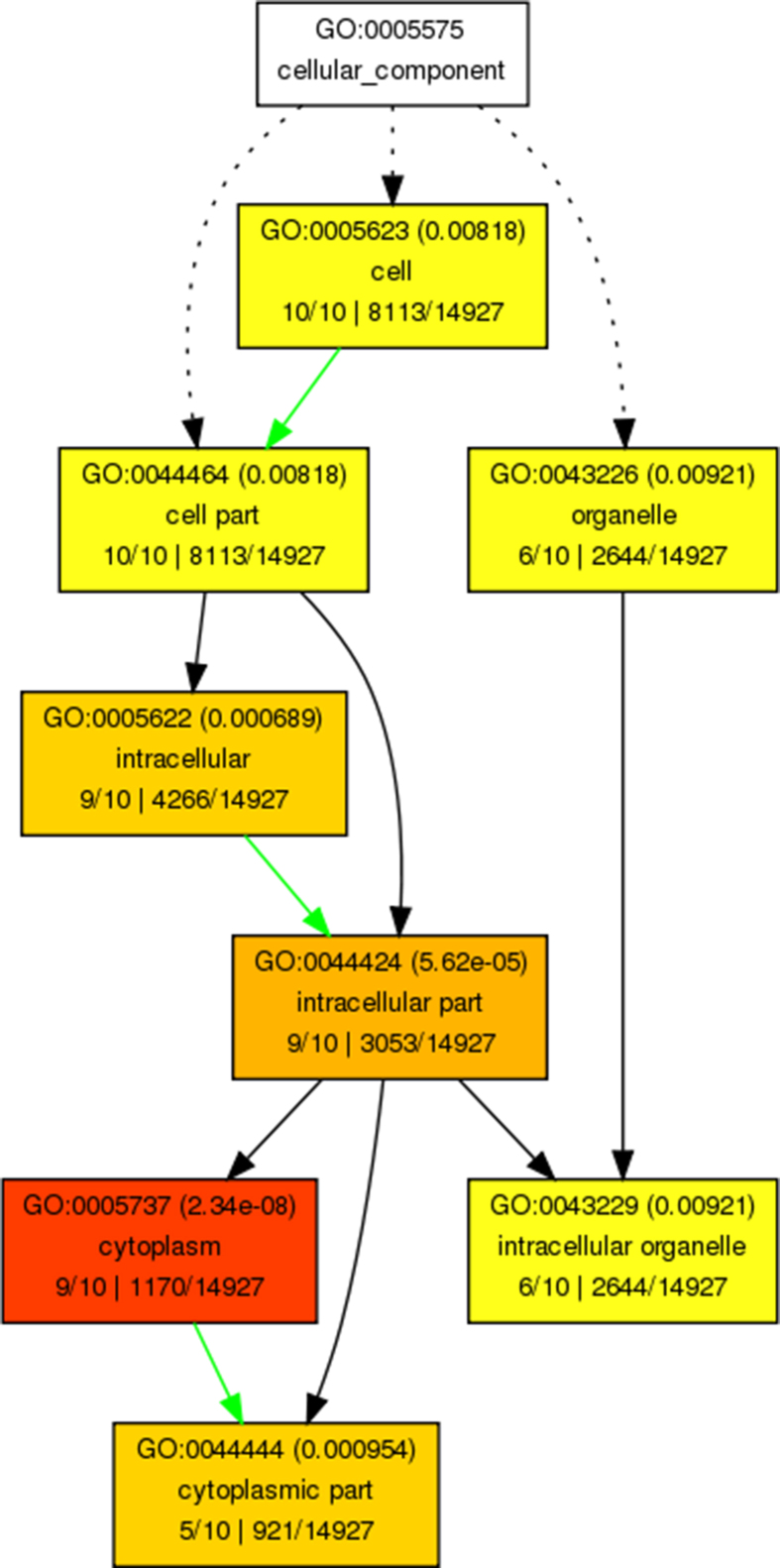 Figure 10
Figure 10Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Disease Management and Epidemiology · interferon and immune responses · Viral Infections and Vectors
Introduction
1
Bovine viral diarrhea (BVD) is a major disease in livestock, causing considerable economic losses to this industry. BVD is associated with gastrointestinal, respiratory, and reproductive disorders in cattle. The etiological agent of the disease is the BVD virus (BVDV), which belongs to the Flaviviridae family, Pestivirus genus. According to latest classification of Pestivirus, this genus has 11 species including pestivirus A (BVDV-1), pestivirus B (BVDV-2), pestivirus C (classical swine fever virus), pestivirus D (border disease virus), and pestiviruses E to K [1]. BVDV is a positive-sense single-stranded RNA virus that replicates in the cytoplasm. The RNA genome is approximately 12.3 kb in length and contains an open reading frame flanked by untranslated regions (UTRs). The ORF encodes a polyprotein precursor that is cleaved into 11 or 12 proteins in noncytopathic (ncp) or cytopathic (cp) virus biotypes, respectively. Mature viral proteins from N-terminus to C-terminus include N_pro_, capsid, E_rns_, E1, E2, p7, NS2-3, or NS2 and NS3, NS4A, NS4B, NS5A, and NS5B. BVDV-1 and BVDV-2 cause the same disease, but they are differ in the 5′-UTR sequences of their viral genomes [2].
Based on the cytopathic effects of BVDV on cell cultures, the virus is classified into two biotypes: cp and ncp [3]. The different features of BVDV biotypes result from differences in the processing of BVDV NS2-3. In cpBVDV infections, the NS2-3 protein is continuously cleaved by NS2 autoprotease activity to form NS3. However, in ncpBVDV infections, NS2-3 cleavage occurs during the early stages, and free NS3 is detected only up to 9 h post-infection. Therefore, the effects of NS2-3 and NS3 on cellular signaling appear to be different [4].
Viruses are intracellular parasites that need cells to replicate and survive. During various stages of viral infection, numerous interactions occur between host and viral proteins [5,6]. Investigating these interactions can improve our understanding of viral pathogenesis, help identify putative functional proteins, and facilitate the discovery of new treatment and prevention strategies. Host-viral protein-protein interactions (PPIs) have been studied through experimental approaches such as yeast two-hybrid, co-immunoprecipitation, tandem affinity purification-mass spectrometry, affinity chromatography, protein microarrays, protein-fragment complementation, and/or computational programs. Researchers have applied network biology approaches to construct disease networks that explore host-pathogen relationships and disease pathways [[6], [7], [8], [9], [10]]. Modeling virus-host interactome maps can help identify functionally important proteins [6], sub-networks, and potential hubs involved in the pathogenesis of specific diseases [7]. Researchers can then study these components using various bioinformatics methods. Viral proteins target bottlenecks, hubs, and rich clubs within cellular protein networks, thereby affecting cellular processes such as transcription, translation, cell signaling, cell trafficking, and cell cycle regulation [5,[11], [12], [13]].
The interaction of some BVDV proteins with host cell proteins has been previously investigated using experimental methods; however, there is no a single comprehensive network that encompasses all the research conducted on the BVDV interactome. This study aimed to combine previous works on BVDV-host interactions and construct a PPI network that encompasses all aspects of this topic. The goal of this study was to identify the proteins that significantly contribute to infection pathways, viral pathogenesis, and the virus's effects on host cells.
Material and methods
2
Data Collection
2.1
To perform this study, in the first step, all data related to the BVDV protein-protein interactions were collected. For this purpose, the advanced search option of PubMed was used. Multiple keywords related to interactions between BVDV and host proteins or virus proteins were applied. The search yielded 2690 research studies in which the virus was studied in different aspects including pathogenesis, epidemiology, and others. After a careful review of the articles, 24 papers contained the PPI data (Table 1) [2,4,[14], [15], [16], [17], [18], [19], [20], [21], [22], [23], [24], [25], [26], [27], [28], [29], [30], [31], [32], [33], [34], [35]]. In these articles, various PPI detection methods such as yeast two-hybrid, immunoprecipitation, co-immunoprecipitation and mass spectrometry were carried out. Besides the data, other PPI database including STRING Viruses [36], VirusMINT [37], IntAct [38], VirHostNet [39], and VirusMentha [40] were investigated. The information related to every single interaction of BVDV-1 or BVDV-2 with host were gathered. Because the BVDV-1 and BVDV-2 genes are identical and differ only in the 5′-UTR region, one network was drawn for both. In the context of virus–host interactions, protein-protein interfaces can be classified as exogenous or endogenous. Host-host or virus-virus PPIs are classified as endogenous interfaces, while interactions between different proteomes including virus–host PPIs are grouped as exogenous interfaces [41,42]. In the present study, host-host endogenous interfaces were excluded. All data gathered were convert to a unique format in order to construct a network in Cytoscape, because all data must be in the same format. Different identifiers including protein name, Uniprot, EMBL/GenBank, and others were converted to the same format, which here was the name gene. This work was done by an online Uniprot ID Mapping (http://www.uniprot.org/mapping/) [43]. Then, the data gathered from all resources were merged and duplicates were removed.Table 1. List of studies which reported the protein-protein interaction data of BVDV-host.Table 1. NoPaperNo. of interactionsReferences1Weiskircher et al., 2009222Yamane et al., 2009243Lazar et al., 20031144Hilton et al., 20031155Zahoor et al., 20101166Gong et al., 20201177Johnson et al., 20011188Leveringhaus et al., 20201199Mu et al., 202112010Branza-Nichita et al., 200412111Fu et al., 201442212Darweesh et al., 201812313Schaut et al., 201512414Xu et al., 199742515Branza-Nichita et al., 200112616Maurer et al., 200412717Lackner et al., 200512818Chen et al., 200712919Dubrau et al., 201723020Mohamed et al., 201413121Wang et al., 201173222Jefferson et al., 2014893323Ammari et al., 2017723424Gladue et al., 20143135
PPI network Construction and analysis
2.2
The latest version of Cytoscape (v3.9.0) [44] was used to construct the BVDV-host PPI network. Cytoscape is a user-friendly platform which help to construct, visualize and analyze of the biomolecular interaction networks. This platform has various tools and plugins for understanding and exploration of the various aspects of the network. Cytoscape Analyzer tool was used to define the topological characters of the network including diameter, radius, average number of neighbors, density, clustering coefficient, and so on.
Hub genes and modules analysis
2.3
CytoHubba (v0.1) [45], a plugin in Cytoscape was used to identify the hub genes. Top ten genes were selected with five algorithms in cytoHubba including MCC, Density of Maximum Neighborhood Component (DMNC), Edge Percolated Component (EPC), Radiality centrality, and Stress centrality. Different clustering algorithms including MCODE (v2.0.0) [46], SCODE (v1.0.4) [47], CytoCluster (v2.1.0) [48], and ClusterViz (v1.0.3) [49] were applied to construct modules and sub-networks. These are plugins in Cytoscape software for clustering nodes based on different algorithms. These plugins can find densely connected nodes [50], important protein complexes [2] and valuable functional motifs. Default parameters were applied for all plugins to construct sub-networks.
Functional enrichment analysis
2.4
Gene enrichment analysis was performed to evaluate the biological pathways of proteins that might be involved in the development and pathogenesis of BVDV. KEGG pathway enrichment analysis was performed using free web based search engine DAVID program (v6.8) with P < 0.05 threshold (https://david.ncifcrf.gov/) [51]. GO enrichment analysis was done using AgriGO (v2.0) to evaluate the CC, MF, and BP of involving host proteins. Interacting host proteins were analyzed with Singular Enrichment Analysis (SEA) tool. Gene names were converted to UniProtKB format because AgriGO allowed ID types including Bos taurus NCBI-GI, ENSEMBL ID, Bovine genome locus (Bovine Genome Database), GenBank ID, DDBJ ID, EMBL ID, UniProt ID, RefSeq Peptide ID, PDB ID, and Bovine Affymetrix Genome Array. The AgriGO can be found at (http://bioinfo.cau.edu.cn/agriGO/) [52].
BVDV-interacting host proteins were entered into the PANTHER classification program (v16.0). The PANTHER system classifies proteins and their genes according to family and subfamily, molecular function, biological process, and pathway. The PANTHER is online freely program and can be found at (http://www.pantherdb.org/) [53].
Results
3
The BVDV-host protein interaction network
3.1
After extracting data from literature and databases, the PPI network was constructed using Cytoscape. In total, 224 interactions were confirmed by experiment-derived data from previous studies. These interactions involved viral proteins including N_pro_ (99 interactions), NS3 (77 interactions), E2 (34 interactions), E_rns_ (5 interactions), NS5A (4 interactions), NS4A (one interaction), C (one interaction), E1 (one interaction), NS4B (one interaction), and E1 (one interaction) with host or viral proteins. The total number of interactions derived from previous studies and data bases was 561; however, after removing duplicates, 279 nodes and 436 interactions were identified (Fig. 1). Among the 279 nodes, 12 were viral proteins, while remaining were host proteins. N_pro_ with 125 connections, had the greatest number of interactions with host proteins, followed by NS2-3, NS3, and E2, with 94, 87, and 65 associations, respectively. Fig. 2 shows the number of interactions between each BVDV-protein and host proteins. The PPI network was analyzed using the network analysis tool, and important statistical features are presented in Table 2.Fig. 1BVDV-Host protein interaction network constructed and visualized using Cytoscape. Red nodes represent host genes, green nodes represent virus genes, and size of nodes represent the number of genes which interacted with the node. Larger nodes have more interactions with the genes. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)Fig. 1. Fig. 2A pie chart representing the number of BVDV genes interaction with host genes. In each section, the name of the virus gene and the number of interactions with the host genes are represented.Fig. 2. Table 2Topological characters of BVDV-host network.Table 2. Number of nodes279Number of edges436Avg. number of neighbors3.118Network diameter8Characteristics path length3.506Clustering coefficient0.012Network density0.006
Screening of hub genes and modules analysis
3.2
Different ranked algorithms in CytoHubba were used to identify host hub genes. The ranks of top ten hub genes selected by each feature are shown in Table 3. CD46 and EEF2 genes were identified by all five algorithms, also TXN2 was detected by four of five ranked methods in cytoHubba.Table 3. Top ten hub genes detected using different topological algorithms in cytoHubba.Table 3. AlgorithmMCCDMNCMNCRadialityStress1CD46aCD46aCD46aCD46aCD46a2EEF2aEEF2aEEF2aTXN2bTXN2b3TXN2bACTBTXN2bEEF2aEEF2a4ACTBHAX1TNEM70RPLPORPLPO5HAX1PCBP1CNDP2RPL7ARPL7A6PCBP1EZRPNMA1RPL24RPL247EZRACTG1ACTG1RPL3RPL38ACTG1RACK1TMED 10RPS3RPS39RACK1GALK1AIPACTBELOC10GALK1G1K1R6G1K1R6HAX1UPF1MCC: Maximal Clique Centrality; DMNC: Density of Maximum Neighborhood Component; EPC: Edge Percolated Component.aThe hub genes identified by all algorithms in cytoHubba.bThe hub genes identified by four ranked algorithms in cytoHubba.
Different sub-networks were constructed using various clustering algorithms to find important protein complexes and functionally modules in BVDV-host protein interaction network. Fig. 3 shows multiple clusters made for the whole network by Molecular Complex Detection (MCODE), CytoCluster, ClusterViz, and Supervised Complex Detection (SCODE). Two sub-cluster were constructed by MCODE. Sub-cluster 1 had 6 nodes in which 4 nodes belonged to viral proteins and 2 belonged to host (Fig. 3A). Sub-cluster 2 consisted of 4 nodes, two of which belonged to host genes (Fig. 3B). In the cluster made by CytoCluster, there were 3 nodes among which NS2 belonged to viral proteins while CD46 and DNAJC14 were bovine proteins (Fig. 3C). According to cluster analysis performed by ClusterVis, four smaller sub-networks were constructed. First sub-network (Fig. 3D) contained 106 nodes among which NS2, NS3, NS2-3, NS4B, and NS5B were viral proteins and the others belonged to host proteins. Second sub-network consisted of 92 nodes, in which N_pro_ was viral protein (Fig. 3E). Third sub-network had 63 nodes among which E2 was viral protein (Fig. 3F). Forth sub-network was consisted of viral protein NS5A and five host proteins (Fig. 3G). Cluster constructed by SCODE had 3 nodes of E_rns_ and E2 (viral proteins), and ATG14 (host protein).Fig. 3. Key moddules for the whole network by MCODE, CytoCluster, ClusterViz, and SCODE. (A and B) Sub-clusters constructed using the MCODE plugin. (C) Sub-cluster identified using CytoCluster. (D, E, F, and G) Sub-clusters constructed using the ClusterVis plugin. Virus genes are showon in red, while host genes are presented in purple. (H) Modules constructed using SCODE. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)Fig. 3
KEGG and GO enrichment analysis
3.3
Kyoto Encyclopedia of Genes and Genomics (KEGG) pathway analysis was performed using the Database for Annotation, Visualization, and Integrated Discovery (DAVID) database for all host proteins interacted with BVDV and specifically top ten hub genes ranked by CytoHubba based on Maximal Clique Centrality (MCC) algorithm. Table 4 shows the KEGG pathway analysis for all BVDV-interacting host proteins. The interacting proteins were highly enriched in ribosome, spliceosome, Influenza A, protein processing in endoplasmic reticulum, proteoglycans in cancer, measles, RNA transport, and protein export while KEGG pathway analysis of top 10 hub genes revealed that these genes were involved in leukocyte transendothelial migration. The Gene Ontology (GO) function analysis was carried out using AgriGO. The results showed that the proteins were significantly involved in biological processes such as metabolic (98 genes), cellular (122 genes), localization (28 genes), response to stimulus (21 genes), biosynthetic (60 genes), gene expression (63 genes), translation (43 genes), multicellular organismal (23 genes), developmental (19 genes), etc processes (Fig. 4A). For GO cellular component analysis, the proteins were mainly associated with cytoplasm (116 genes), organelle (112 genes), nucleus part (41 genes), ribosome (34 genes), cytoskeleton (20 genes), mitochondrion (14 genes), and endoplasmic reticulum (12 genes) (Fig. 4B). The proteins significantly enriched in molecular functions including binding (125 genes), protein binding (91 genes), structural molecule activity (38 genes), structural constitute of ribosome (32 genes), RNA binding (32 genes), translation factor activity (5 genes), etc (Fig. 4C). The GO cellular component (CC), and molecular function (MF) of the top hub genes ranked by MCC algorithm are shown in Supplementary Fig. 1. The GO biological process (BP) of hub genes was not significant (P > 0.05).Table 4. Kyoto encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of host genes interacted with BVDV genes.Table 4. TermCount (%)P-ValueBenjaminiRibosome35 (14.5)8.7E-301.4E-27Spliceosome13 (5.4)5.7E-64.7E-4Influenza A10 (4.1)5.7E-32.4E-1Protein processing in endoplasmic reticulum9 (3.7)1.5E-25.1E-1Proteoglycans in cancer8 (3.3)9.3E-21.0E0Measles7 (2.9)5.1E-21.0E0RNA transport7 (2.9)8.3E-21.0E0Protein export5 (2.1)1.0E-35.5E-2Fig. 4The Gene Ontology (GO) enrichment analysis using AgriGO**.** (A) GO terms biological process (BP). (B) GO terms cellular compenets (CC). (C) GO terms molecular functions (MF). GO term codes are shown in braces.Fig. 4
PANTHER classification of proteins
3.4
The host proteins that interact with BVDV were classified using the Protein Analysis Through Evolutionary Relationships (PANTHER) Classification System. According to the protein classes of host targeted genes, translational proteins, nucleic acid metabolism proteins, metabolite interconversion enzymes, and protein modifying enzymes were the most likely candidates as targets of viral proteins (Fig. 5). Some of these proteins involve in various biological categories as cellular proteins often play several roles in several pathways.Fig. 5. Panther pie-chart of host interacting proteins classification. The three main GO terms were subclassified to child categories (pie charts at the bottom of the image). GO term codes are shown in braces.Fig. 5
Discussion
4
We constructed a comprehensive BVDV-host protein interaction map to describe the interactome between BVDV and host proteins, aiming to better understand the pathogenicity of this virus and identify potential drug targets for the treating of BVDV infection. Although BVDV, like other RNA viruses, produces few proteins, these proteins often interact with multiple host proteins to create favorable conditions for the virus's survival and replication [14]. Identifying of host proteins targeted by viral proteins and investigating under both of their functions in normal and infectious conditions can help us to uncover the key processes required for infection [14]. In this context, network biology can be a useful tool for studying host-pathogen relationships at the molecular level. Various researchers have used network biology to discover key viral proteins and their targets in infection processes [6,7,9]. In the present study, all previous small-scale, large-scale, and computational investigations on BVDV-host interactions were integrated to detect highly BVDV-associated host proteins and their roles in viral pathogenesis.
In cellular network analysis, proteins with a wide range of connecting partners are called “hubs” [54]. After constructing the PPI network, 10 hub genes were identified using five different ranking methods. The CD46, EEF-2, and TXN genes were detected by at least four out of the five ranking methods in cytoHubba. CD46, the most prominent identified hub gene, is a ubiquitously expressed “multitasker,” found on all nucleated cells, protecting them from autologous damage by the complement system [55,56]. CD46 regulates both adaptive immunity and the complement system [55]. It has been identified as interacting with other pathogens, such as like the measles virus [55], human herpesvirus 6 [56], group B adenoviruses [57], and pathogenic Neisseria [58]. TXN-2, also known as Trx-2, is involved in various cellular processes, including the regulation of ROS production, apoptosis, cell growth, and the modulation of transcription factors [59]. Researchers have reported that TXN-2 plays a role in the life cycle of the classical swine fever virus (CSFV) and BVDV [59,60]. TXN-2 inhibits CSFV replication, and in the case of BVDV, it is downregulated by the virus during early infection. EEF-2 encodes eukaryotic elongation factor 2, which is essential for protein synthesis [61].
KEGG pathway analysis revealed that BVDV proteins primarily target biological pathways involved in translation and protein synthesis, including ribosome, spliceosome, protein processing in the endoplasmic reticulum, and RNA transport. BVDV interacts with ribosomal complexes to ensure that viral RNAs are translated and can produce substantial amounts of viral proteins even under the strict conditions imposed by the host shutoff mechanism, where cellular metabolism is severely limited [14,62]. As found in the present study and reported by other researchers, BVDV manipulates biological processes such as cellular metabolism, gene expression, translation, cell development and differentiation [[62], [63], [64]]. The cellular proteins and pathways targeted by BVDV proteins, including RNA binding, protein binding, spliceosome, translation, and biosynthesis, are also involved in the pathogenesis of other viruses, indicating common strategies among these viruses [54,63,65]. According to KEGG analysis, the enriched gene set is found to be involved in other infectious pathways such as Influenza A virus and measles infections.
GO enrichment analysis revealed that most interacting proteins function in various biological processes, including cellular protein metabolism, cellular macromolecular biosynthesis, gene expression, and translation. These proteins are primarily found in the nucleus, ribosome, cytoskeleton, mitochondrion, and endoplasmic reticulum. Most of the host-interacting proteins have binding molecular functions, such as protein binding, RNA binding, translation factor activity-nucleic acid binding, and cytoskeletal protein binding. Some of these proteins also serve as structural constituents of the ribosome. The PANTHER, GO, and KEGG evidence highlights the widespread effects of BVDV on cellular processes, particularly in the translation and synthesis of proteins, as reported in other studies [62,63].
Conclusion
5
Although BVDV encodes few proteins (like other RNA viruses), it can effectively affect multiple host pathways. The BVDV-host PPI network map, constructed based on small-scale, large-scale, and computational studies, revealed that viral proteins target various protein classes, including translational proteins, nucleic acid metabolism proteins, metabolite interconversion enzymes, and protein-modifying enzymes. CD46, EEF-2, and TXN were identified as hub genes using different ranking algorithms. The present study shows that BVDV interactions with its host primarily focus on targeting translation, protein synthesis, and cellular metabolic pathways. Further studies are needed to explore the effects of these proteins on cellular pathways and the molecular mechanisms of BVDV infection.
Funding
No funding.
Compliance with ethical standards
Not applicable.
Availability of data and materials
The datasets used and/or analyzed during the current study are available through the paper and additional files.
CRediT authorship contribution statement
Seyedeh Elham Rezatofighi: Writing – original draft, Visualization, Validation, Supervision, Project administration, Methodology, Investigation, Formal analysis, Data curation.
Declaration of competing interest
The authors declare no conflict of interests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Walker P.J.Siddell S.G.Lefkowitz E.J.Mushegian A.R.Adriaenssens E.M.Dempsey D.M.Dutilh B.E.Harrach B.Harrison R.L.Hendrickson R.C.Changes to virus taxonomy and the statutes ratified by the international committee on taxonomy of viruses Arch. Virol.16520202737274810.1007/s 00705-023-05797-432816125 · doi ↗ · pubmed ↗
- 2Weiskircher E.Aligo J.Ning G.Konan K.V.Bovine viral diarrhea virus NS 4B protein is an integral membrane protein associated with Golgi markers and rearranged host membranes Virol. J.6200918510.1186/1743-422X-6-18519887001 PMC 2777160 · doi ↗ · pubmed ↗
- 3Ridpath J.Bolin S.Dubovi E.Segregation of bovine viral diarrhea virus into genotypes Virology 2051994667410.1006/viro.1994.16207975238 · doi ↗ · pubmed ↗
- 4Yamane D.Zahoor M.A.Mohamed Y.M.Azab W.Kato K.Tohya Y.Akashi H.Inhibition of sphingosine kinase by bovine viral diarrhea virus NS 3 is crucial for efficient viral replication and cytopathogenesis J. Biol. Chem.2842009136481365910.1074/jbc.M 80749820019293152 PMC 2679466 · doi ↗ · pubmed ↗
- 5Halehalli R.R.Nagarajaram H.A.Molecular principles of human virus protein-protein interactions Bioinformatics 3120121025103310.1093/bioinformatics/btu 76325417202 · doi ↗ · pubmed ↗
- 6Farooq Q.U.A.Shaukat Z.Zhou T.Aiman S.Gong W.Li C.Inferring Virus-Host relationship between HPV and its host Homo sapiens using protein interaction network Sci. Rep.102020871910.1038/s 41598-020-65837-w 32457456 PMC 7251128 · doi ↗ · pubmed ↗
- 7Qua Farooq Khan F.F.Construction and analysis of a comprehensive protein interaction network of HCV with its host Homo sapiens BMC Infect. Dis.19201936710.1186/s 12879-019-4000-931039741 PMC 6492420 · doi ↗ · pubmed ↗
- 8Wei W.Kong W.Identification of key genes and signaling pathways during Sendai virus infection in vitro Braz. J. Microbiol.502019132210.1007/s 42770-018-0021-630637656 PMC 6863206 · doi ↗ · pubmed ↗