Characterization of temperature-sensitive alleles of Schizosaccharomyces pombe septation initiation network components
Lesley A. Turner, Alaina H. Willet, Kathleen L. Gould

TL;DR
This paper characterizes temperature-sensitive mutations in key components of a cell division network in a type of yeast.
Contribution
The study identifies and characterizes new temperature-sensitive alleles in four SIN components of Schizosaccharomyces pombe.
Findings
Temperature-sensitive mutations in cdc14, cdc16, sid1, and sid2 were characterized.
The mutations affect SIN signaling and growth at different temperatures.
These new mutants provide tools for studying cell division mechanisms.
Abstract
The Schizosaccharomyces pombe septation initiation network (SIN) promotes cytokinesis and septation. Comprised of a protein kinase cascade triggered by activation of a small GTPase and inhibited by a two-component GAP that localize to the spindle pole bodies in a cell cycle specific manner. Here, we characterized temperature-sensitive mutants isolated in the 1990s in four SIN components. We determined the mutations within each cdc14 , cdc16 , sid1 , and sid2 mutant allele and analyzed their growth at different temperatures compared with known mutant alleles. The new mutants described here expand the toolkit for studying SIN signaling.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
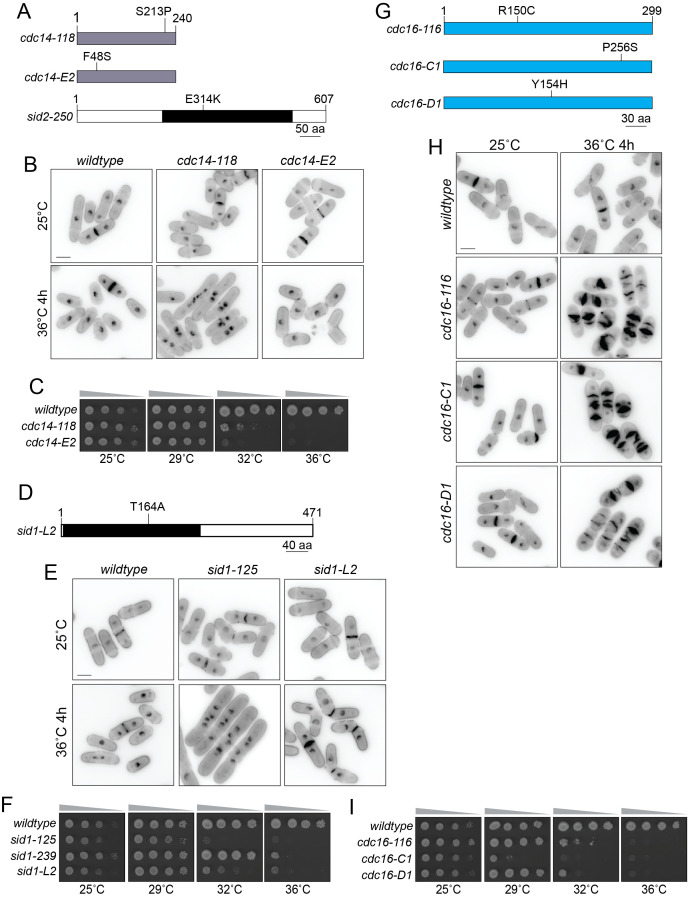 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAdipose Tissue and Metabolism · Invertebrate Immune Response Mechanisms · Lipid metabolism and biosynthesis
Description
Cell division in the yeast Schizosaccharomyces pombe requires a signaling cascade termed the septation initiation network (SIN) (reviewed in Cullati and Gould, 2019; Simanis, 2015; Xiao and Dong, 2021). The SIN is necessary for normal assembly, maintenance, and constriction of the actin- and myosin-based machinery required for cell division as well as activation of the cell wall enzymes necessary for septation (Cheffings et al., 2016; Glotzer, 2017; Mangione and Gould, 2019) . In *sin * mutants, cytokinesis fails and cells become elongated and multinucleate.
Activation of the SIN pathway is driven by the Spg1 GTPase (Schmidt et al., 1997; Sohrmann et al., 1998) that in turn activates the Cdc7 protein kinase (Fankhauser and Simanis, 1994; Schmidt et al., 1997) . The Sid1 kinase in complex with its cofactor Cdc14 acts next in the pathway followed by the Sid2 protein kinase in complex with its cofactor Mob1 (Fankhauser and Simanis, 1993; Guertin et al., 2000; Hou et al., 2000; Salimova et al., 2000; Sparks et al., 1999) . The Sid2- Mob1 complex is the only component of the SIN that localizes to the cell division site (Hou et al., 2000; Salimova et al., 2000; Sparks et al., 1999) . Byr4 and Cdc16 comprise a two-component GAP for Spg1 and are major SIN inhibitors (Furge et al., 1998; Minet et al., 1979) . SIN mutants have been isolated in genetic screens for general cell cycle regulators and also in those targeting cytokinesis factors (Balasubramanian et al., 1998; Nurse et al., 1976) . In several cases, the mutations within the mutant alleles have not been identified.
In addition to the cdc14-118 allele described and characterized previously (Marks et al., 1992; Nurse et al., 1976) , we isolated a second mutant allele mapping to the * cdc14 * gene, cdc14-E2 (Balasubramanian et al., 1998) . The * cdc14 * open reading frame (ORF) was amplified from each of the two strains and sequenced to determine what mutation was present. While the cdc14-118 allele encoded a S213P substitution, the cdc14-E2 allele encoded a F48S substitution ( Figure 1A ). To compare the cell phenotypes, we examined each mutant by staining for nuclei and septa after the cells were grown at 25°C and then shifted or not to 36˚C for 4 hours ( Figure 1B ). While cdc14-118 cells showed the classic *sin * phenotype of multinucleation and cell elongation at the non-permissive temperature, cdc14-E2 cells arrested uniformly at a very late stage of septation and frequently lysed ( Figure 1B ). A spot assay showed that the cdc14-E2 allele was comparable in its temperature sensitivity to cdc14-118 ( Figure 1C ).
Although the *sid2-250 * mutant has been extensively analyzed (Balasubramanian et al., 1998) , the identity of the causative mutation has not been reported. The * sid2 * open reading frame was therefore amplified from sid2-250 cells, sequenced, and a single mutation was identified leading to a E314K substitution within the catalytic domain ( Figure 1A ).
In our screen for cytokinesis mutants, we identified several mutants that mapped to the * sid1 * locus, one ( sid1-L2 ) that has not been previously characterized, and two mutants mapping to the * cdc16 * locus, cdc16-C1 and cdc16-D1 (Balasubramanian et al., 1998) . To determine if the *sid1-L2 * allele differed from sid1-125 (L114P) and *sid1-239 * (L12P) *, * the * sid1 * ORF was amplified from it and sequenced. A single point mutation causing a T164A substitution within the catalytic domain was found ( Figure 1D ). Nuclei and septa staining revealed predominantly a boomerang-shape phenotype that was often accompanied by cell lysis at septation ( Figure 1E ). A spot assay revealed that sid1-L2 had an intermediate restrictive temperature compared to sid1-125 and sid1-239 ( Figure 1F ).
The * cdc16 * ORFs were also amplified from cdc16-116, cdc16-C1, and cdc16-D1 and sequenced to determine if the C1 and D1 alleles differed from cdc16-116 . We found only single mutations in each ORF leading to three distinct amino acid substitutions ( Figure 1G ). To compare the cell phenotypes, we examined each mutant by staining for nuclei and septa after the cells were grown at 25°C and then shifted or not to 36˚C for 4 hours. The phenotypes of the three mutants were comparable. At 25˚C, the percent of septated cells was 17-20 with none showing more than one septa and at 36˚C, all cells arrested with multiple septa and one or two nuclei ( Figure 1H ). We next determined the range of temperature-sensitivity of each * cdc16 * allele by spotting at a variety of temperatures. All temperature-sensitive alleles grew less than wildtype at 36°C with the cdc16-C1 allele showing the greatest temperature-sensitivity ( Figure 1I ).
In sum, we have provided an initial characterization of new mutants of SIN components that expand the repertoire of reagents which can be used to study SIN signaling.
Methods
Yeast methods
S. pombe strains were grown in yeast extract (YE) and standard S. pombe mating, sporulation, and tetrad dissection techniques were used to construct new strains (Moreno et al., 1991) . All spot assays were performed twice with reproducible results.
Molecular biology methods
cdc14 * alleles were amplified using an oligonucleotide 20 bp upstream of the start site (TATTGCCCGCTTGGCATGAG) and another 20 bp downstream of the stop codon (TAAGTTAACAATGAGACTTTAAACATT) (Integrated DNA technologies). * cdc16 * alleles were amplified using an oligonucleotide 50 bp upstream of the start site (GAATTCATACTGGTCCTCATTTTAGT) and another 77 bp downstream of the stop codon (GAGTGAGAGGTGTGTGCTGA) (Integrated DNA technologies). The * sid2 * allele was amplified using an oligonucleotide 70 bp upstream of the start site (ACACGTAAGGTTATTATTGACAGGAG) and another 50 bp downstream of the stop codon (CATCAAAAGCGAAGCTCAGTATCTTC) (Integrated DNA technologies). The PCR products were each sequenced by Plasmidsaurus (Eugene, OR) using Oxford Nanopore Technology with custom analysis and annotation. The sid1-L2 open reading frame was amplified using an oligonucleotide 70 bp upstream of the start site (AGTACTTCGTGGTGCATCTAGCT) and another 70 bp downstream of the stop codon (GTAGAATATGCCATTATAAGTTCATT) (Integrated DNA Technologies). Sanger DNA sequencing was performed by GenHunter (Nashville, TN) using additional internal forward (GCGTGTCATCTTTGAAATTCCTCAATC) and reverse (CCAATACACTTTGATCCGAATCAT) primers (Integrated DNA technologies).
Microscopy and image analysis
Strains for fixed-cell imaging experiments were grown at 25°C in YE and then shifted to 36°C for 4 hours. Cells were fixed with 70% ethanol for DAPI and methyl blue (MB) staining as described previously (Roberts-Galbraith et al., 2009) . Images were acquired using a Zeiss Axio Observer inverted epifluorescence microscope with Zeiss 63× oil objective (1.46 NA) and captured using Zeiss ZEN 3.0 (Blue edition) software. A singular medial Z slice was obtained. All images were further processed using ImageJ (Schindelin et al., 2012) . All imaging experiments were repeated twice.
Reagents
The strains used in this study and their genotypes are listed below.
Strain Genotype Source
KGY184 * cdc16-116 h ^-^ (Minet et al., 1979) *
KGY187 * cdc14-118 h ^-^ (Nurse et al., 1976) *
KGY246 ade6-M210 leu1-32 ura4-D18 * h ^-^ * Lab stock
KGY1052 * cdc14-E2 ura1 leu1-32 mam2::LEU2 ade6-M216 h ^90^ (Balasubramanian et al., 1998) *
KGY1055 * cdc16-D1 ura1 leu1-32 mam2::LEU2 ade6-M216 h ^90^ (Balasubramanian et al., 1998) *
KGY1057 * cdc16-C1 ura1 leu1-32 mam2::LEU2 ade6-M216 h ^90^ (Balasubramanian et al., 1998) *
KGY1168 sid1-L2 * ade6-M21X ura4-D18 leu1-32 h ^-^ * This study
KGY2089 * sid1-239 leu1-32 h ^-^ * Lab stock
KGY4319 * sid1-125 ura4-D18 leu1-32 ade6-M21X h ^-^ * Lab stock
KGY4875-2 * cdc16-C1 ade6-M21X ura4-D18 leu1-32 h ^-^ * This study
KGY7146-2 * cdc16-D1 ade6-M21X leu1-32 ura4-D18 h ^-^ * This study
KGY9160-2 * cdc14-E2 leu1-32 mam2::LEU2 ura4-D18 h ^90^ * This study
KGY9560-2 * cdc14-E2 leu1-32 ura4-D18 h ^-^ * This study
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Balasubramanian MK Mc Collum D Chang L Wong KC Naqvi NI He X Sazer S Gould KL 199871 Isolation and characterization of new fission yeast cytokinesis mutants.Genetics 14930016-67311265127510.1093/genetics/149.3.12659649519 PMC 1460233 · doi ↗ · pubmed ↗
- 2Cheffings TH Burroughs NJ Balasubramanian MK 201688 Actomyosin Ring Formation and Tension Generation in Eukaryotic Cytokinesis.Curr Biol 26150960-9822 R 719R 73710.1016/j.cub.2016.06.07127505246 · doi ↗ · pubmed ↗
- 3Cullati SN Gould KL 201912 Spatiotemporal regulation of the Dma 1-mediated mitotic checkpoint coordinates mitosis with cytokinesis.Curr Genet 6530172-808366366810.1007/s 00294-018-0921-x 30600396 PMC 6511297 · doi ↗ · pubmed ↗
- 4Fankhauser C Simanis V 199351 The Schizosaccharomyces pombe cdc 14 gene is required for septum formation and can also inhibit nuclear division.Mol Biol Cell 451059-152453153910.1091/mbc.4.5.5318334307 PMC 300956 · doi ↗ · pubmed ↗
- 5Fankhauser C Simanis V 199471 The cdc 7 protein kinase is a dosage dependent regulator of septum formation in fission yeast.EMBO J 13130261-41893011301910.1002/j.1460-2075.1994.tb 06600.x 8039497 PMC 395190 · doi ↗ · pubmed ↗
- 6Furge KA Wong K Armstrong J Balasubramanian M Albright CF 1998827 Byr 4 and Cdc 16 form a two-component GT Pase-activating protein for the Spg 1 GT Pase that controls septation in fission yeast.Curr Biol 8170960-982294795410.1016/s 0960-9822(98)70394-x 9742395 · doi ↗ · pubmed ↗
- 7Glotzer M 2017103 Cytokinesis in Metazoa and Fungi.Cold Spring Harb Perspect Biol 91010.1101/cshperspect.a 02234328007751 PMC 5630000 · doi ↗ · pubmed ↗
- 8Guertin DA Chang L Irshad F Gould KL Mc Collum D 2000417 The role of the sid 1p kinase and cdc 14p in regulating the onset of cytokinesis in fission yeast.EMBO J 1980261-41891803181510.1093/emboj/19.8.180310775265 PMC 302011 · doi ↗ · pubmed ↗