Structure and function of a ligand-free GPCR-Gαβγ intermediate complex
Maxine Bi, Xudong Wang, Jinan Wang, Jun Xu, Wenkai Sun, Victor Ayo Adediwura, Yinglong Miao, Yifan Cheng, Libin Ye

TL;DR
Scientists discovered a new intermediate complex in GPCR signaling using cryo-EM and simulations, revealing how it initiates nucleotide exchange before full activation.
Contribution
The study reveals the structure and function of a ligand-free GPCR-Gαβγ intermediate complex and its role in nucleotide exchange.
Findings
The intermediate GPCR-Gαsβγ complex structure was determined at 2.8 Å resolution using cryo-EM.
The intermediate complex initiates nucleotide exchange without progressing to full activation.
MD simulations confirmed the partial opening of the Gα's α-helical domain during this process.
Abstract
Unraveling the signaling roles of intermediate complexes is pivotal for G protein-coupled receptor (GPCR) drug development. Despite hundreds of GPCR-Gαβγ structures, these snapshots primarily capture the fully activated complex. Consequently, the functions of intermediate GPCR-G protein complexes remain elusive. Guided by a conformational landscape visualized via 19F quantitative NMR and molecular dynamics (MD) simulation, we determined the structure of an intermediate GPCR-mini-Gαsβγ complex at 2.8 Å using cryo-EM, by blocking its transition to the fully activated complex. Furthermore, we presented direct evidence that the intermediate complex initiates a rate-limited nucleotide exchange without progressing to the fully activated complex, in which the α-helical domain (AHD) of the Gα is partially open engaged by a second nucleotide. Our MD simulation supported the pose of the AHD…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Extended Data Fig. 1
Extended Data Fig. 1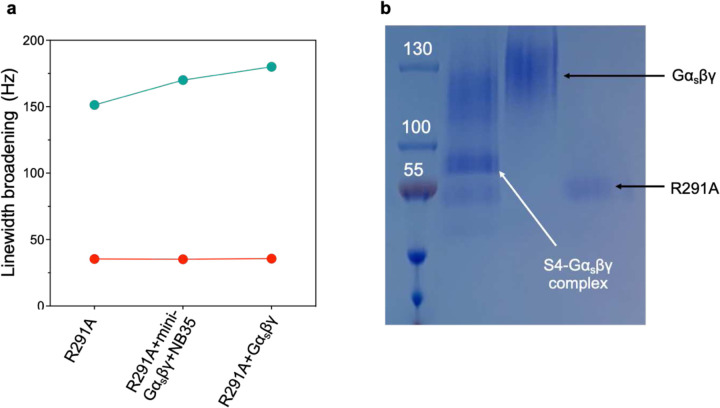 Extended Data Fig. 2
Extended Data Fig. 2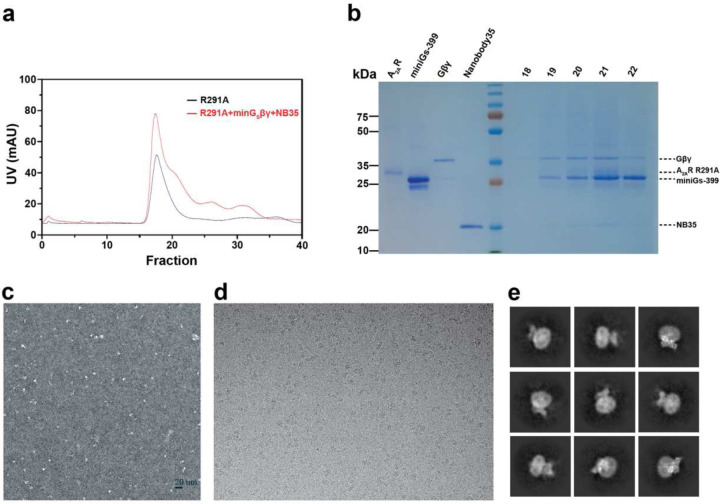 Extended Data Fig. 3
Extended Data Fig. 3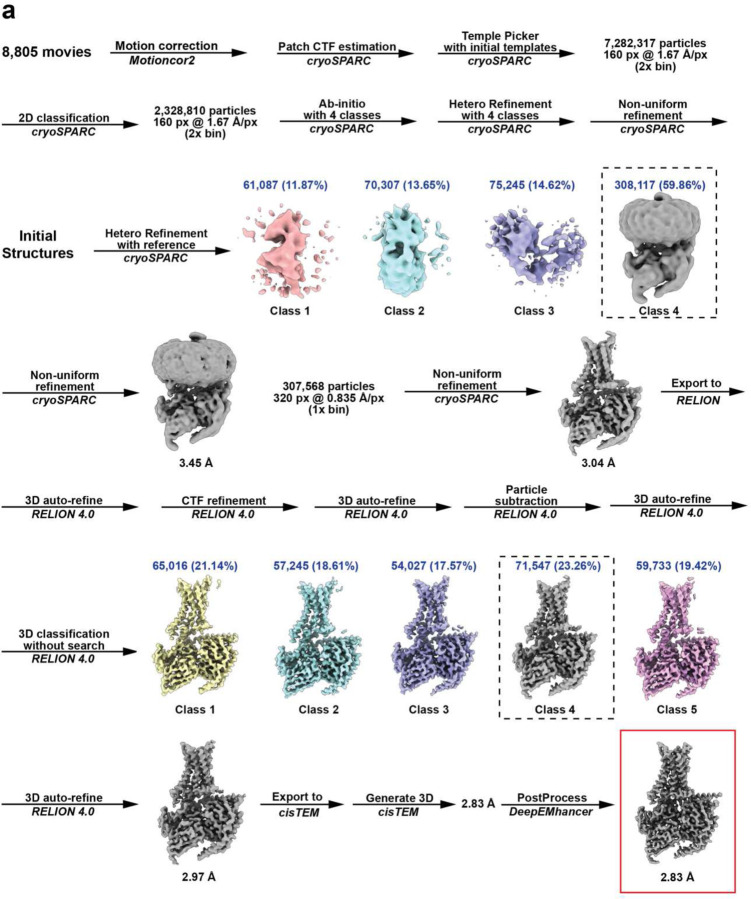 Extended Data Fig. 4
Extended Data Fig. 4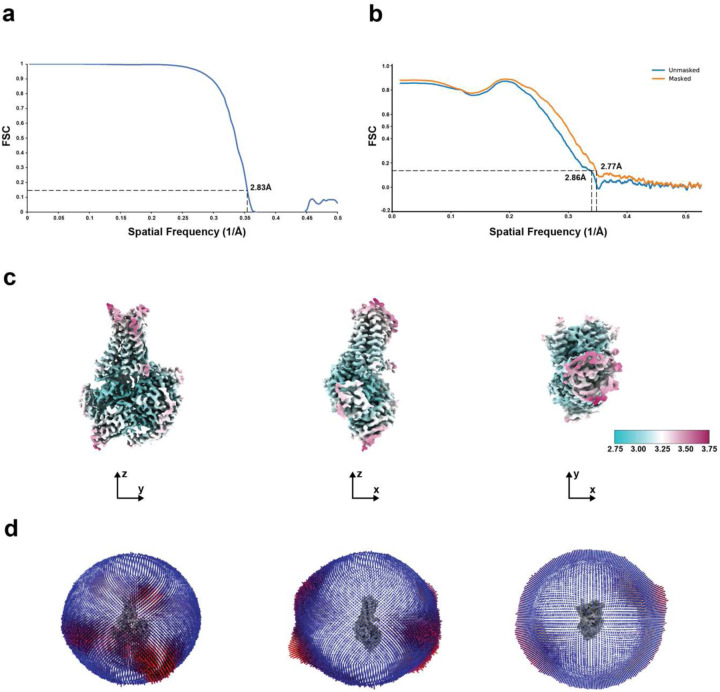 Extended Data Fig. 5
Extended Data Fig. 5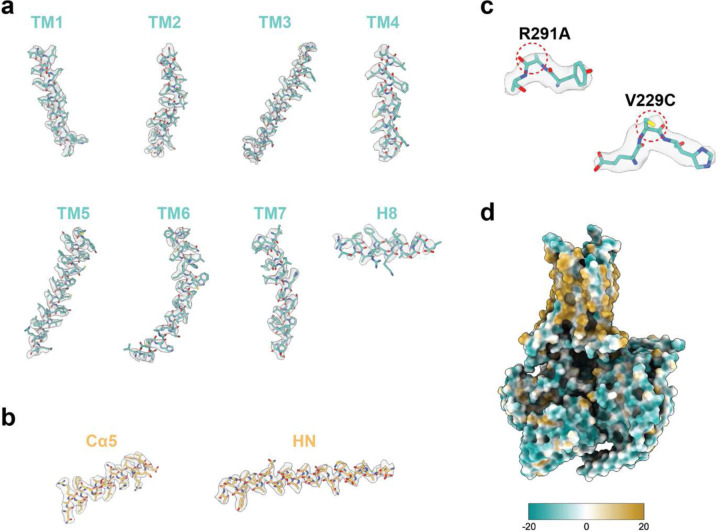 Extended Data Fig. 6
Extended Data Fig. 6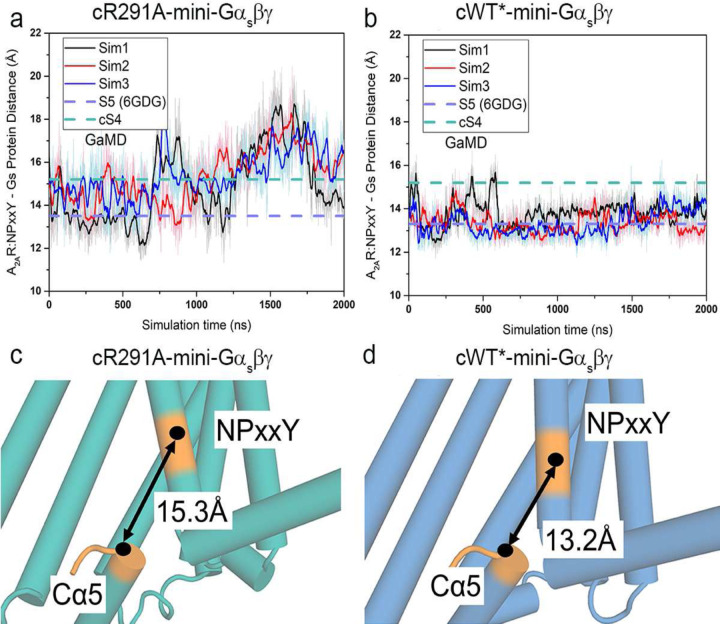 Extended Data Fig. 7
Extended Data Fig. 7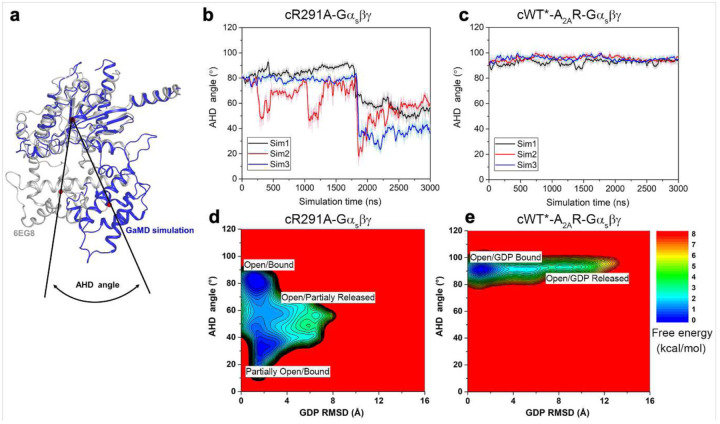 Extended Data Fig. 8
Extended Data Fig. 8Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsReceptor Mechanisms and Signaling · Protein Kinase Regulation and GTPase Signaling · Drug Transport and Resistance Mechanisms
G protein-coupled receptors (GPCRs) are the largest family of human membrane proteins, encompassing over 800 distinct members. Due to their vital roles in various (patho)physiological processes, significant efforts have been made to unravel their activation mechanisms, aiming to modulate therapeutic signaling with precision. Recent advancements of X-ray crystallography and single-particle cryo-electron microscopy (cryo-EM) have facilitated the structural determination of hundreds of GPCR-Gαβγ complex since 2017^1–3^. These structures primarily represent the receptors in their fully activated complexes. Yet, the GPCR signaling process is more intricate than what these structures alone can depict. Multiple studies have shown that GPCR activation entails transitions through several intermediate complexes, including pre-coupled and partially activated GPCR-G protein complexes^4–12^. Yet, isolating such transient complexes to study their signaling roles presents a significant challenge, since it is almost impossible to biochemically isolate GPCR in these intermediate states. Characterizing structural and functional roles of individual intermediate GPCR-G protein complexes will significantly advance our knowledge at the molecular mechanistic level regarding the signaling efficacy, bias, and allostery, which is pivotal for improving drug designs^5,13,14^.
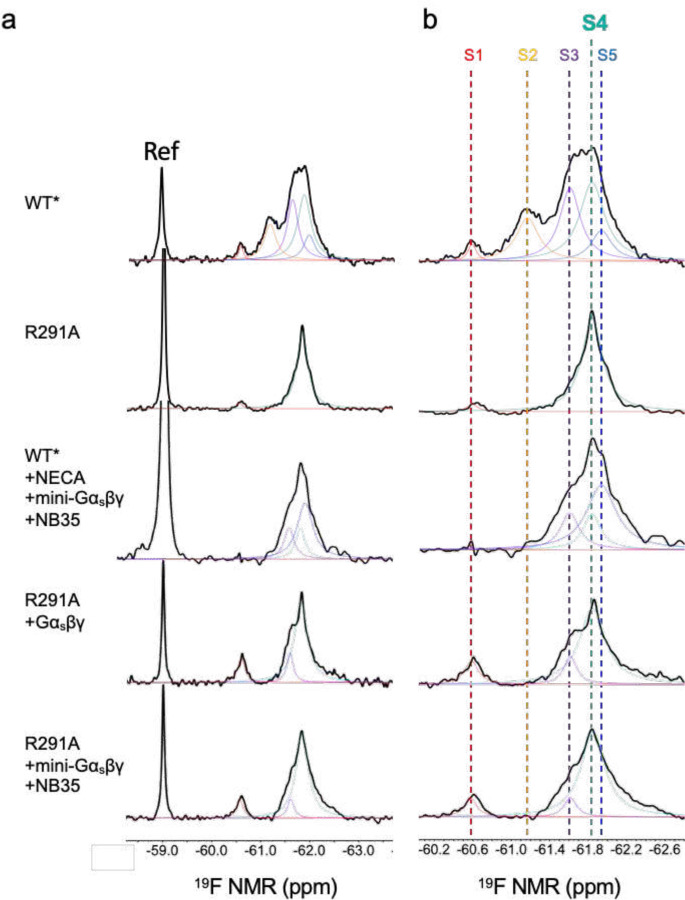
Using the adenosine-A_2A_ receptor (A_2A_R) as a model system and applying ^19^F quantitative NMR (^19^F-qNMR), we visualize the conformational landscape of this GPCR in response to ligand actions and transducers such as G proteins. Within this landscape, we identified two transient active-like intermediates (S3 and S4) that are positioned between the inactive (defined as S1 and S2) and the fully activated states (S5)^15^ (Fig. 1a). Among these conformations, S1 and S2 have been defined by crystal structures^16,17^, and the S5 state has been characterized by cryo-EM (PDB ID: 6GDG)^18^ (Figs. 1b and 1c, and Extended Data Fig. 1). The structures of S3 and S4, however, remain uncharacterized. With the R291A point mutation, which we previously identified to trap A_2A_R in the intermediate S4 state (Fig. 1)^15^, we investigate the structure and function of this intermediate state. We demonstrate that intermediate S4 can directly interact with and regulate G protein for a rate-limited nucleotide exchange without transitioning to the S5 state. We also determine structure of a ligand-free A_2A_R at S4 in complex with mini-Gα_s_βγ, and reveal how the G protein responds to the intermediate S4. Further, biochemical assessments and Gaussian accelerated MD (GaMD) simulation unravel a mechanism of how the intermediate GPCR-G protein complex elicits a slower GTP-GDP exchange prior or to the transition to the fully activated complex. Beyond A_2A_R, our approach can be applied to characterize transient intermediate states of other GPCRs.
S4 state binds to Gαsβγ and initiates nucleotide exchange
We first examined whether the WT*-R291A mutant, which is trapped in the S4 state, can bind to the Gαβγ. Here, the S4 trapped mutant was introduced to a previously defined construct (A_2A_R-316-V229C, denoted as WT*),^15^ and we referred to WT*-R291A simply as R291A. We incubated ^19^F-labeled R291A with Gαβγ, subjected it to ^19^F-NMR, and measured the linewidth of the NMR resonance. Indeed, we observed a linewidth broadening for the S4 resonance compared to the receptor alone, suggesting direct binding of Gαβγ to A_2A_R in the S4 state (Extended Data Fig. 2a). This was further confirmed by a complex band of apo R291A-Gα_s_βγ in native-PAGE (Extended Data Fig. 2b).
Next, we assessed how the S4 regulates G protein function by performing nucleotide hydrolysis, binding, and exchange assays. As shown in Fig. 2a, the R291A mutant-mediated GTP hydrolysis from the G protein occurred at a much slower rate compared to the full agonist-bound WT*-Gα_s_βγ (representing the S5-Gα_s_βγ). However, the addition of full, partial, or inverse agonists did not alter the GTP hydrolysis level substantially in the R291A mutant. This ligand-independent hydrolysis behavior is in stark contrast to the wild-type construct, where ligand binding dramatically changes the hydrolysis level^19,20^. To further investigate the reduced hydrolysis level, we measured the initial rates and Michaelis constants of GTP hydrolysis. The initial rate of GTP hydrolysis mediated by R291A was only one-twentieth of that mediated by WT* (Fig. 2b), while the Michaelis constant exhibited a similar pattern (Fig. 2c).
We proposed a simple model to explain the limited GTP hydrolyzing capacity of the S4-mediated G protein, where nucleotide binding to G protein is a two-step process. A nucleotide GTP initially binds to a site with low affinity and hydrolysis (site1) in the intermediate S4-G protein complex. Upon the transitioning from S4 to S5, the G protein changes its conformation to allow the GTP to access the site 2, the canonical nucleotide-binding site. We tested this model by examining the binding of BODIPY-FL-GDP (Figs. 2d and 2h). The BODIPY-FL-GDP bound to the WT*-mediated G protein exhibited a higher K_D_ than that of the R291A-mediated G protein, although both reached equilibrium at 1.5 hrs. For the R291A-mediated G protein, a secondary association event was observed after 1.5 hrs with a K_D_ value 10 times lower, which was not the case observed in the WT*. We attribute this difference to the restricted accessibility of the site 2, caused by spatial constraints resulting from the partially open AHD domain in the S4 mediated G-protein, while this site is freely accessible in the WT*. A similar pattern was noted in BODIPY-FL-GTP binding assays, where BODIPY-FL-GTP showed more efficient binding to the S5-mediated G protein with a higher K_D_ value (Fig. 2e and 2h). The spatial limited association process was also evident in BODIPY-FL-GTP-γ-S (Fig.2f) in both S5 and S4 mediated G protein because the BODIPY head was linked to γ-phosphate, which is meant to be inserted into the site 2. The bulky BODIPY head insertion restricted exchange and hydrolysis. Titration of GTP into the BODIPY-FL-GDP equilibrium system led us to conclude that the S4-mediated G protein has a reduced nucleotide exchange rate compared to the S5-mediated G protein (Fig. 2g). A previous study of β_2_AR indicated that GDP release occurs much faster than the formation of fully activated complex^21^, while MD simulations suggested that GDP is released when the Gα_s_ is not fully opened^22^. Collectively, our data suggests the intermediate S4 state mediated G-protein initiates nucleotide exchange before transitioning to the fully activated S5 state, albeit at a reduced rate.
Structure of the ligand-free intermediate S4-mini-Gαsβγ complex
Building upon the two-step nucleotide binding model, we hypothesized that the S4 state not only adopts a conformation distinct from S5 but also binds to the G protein in a manner conformationally different from the S5-bound state. To test this hypothesis and enhance our understanding of the molecular mechanisms involved, we aimed to determine the structure of the intermediate S4 state in complex with the G protein.
To capture the structure of intermediate S4 state in complex with G proteins, we assembled a stable complex by incubating the R291A mutation with mini-Gα_s_, known as mini-Gα_s_399, and the Gα_s_-stabilizing nanobody, NB35, in conjunction with Gβ1γ2. As evidenced by ^19^F-qNMR, the receptor preserves the conformation of the S4 state in complex with G proteins (Fig.1b and 1c). We determined a cryo-EM structure of this S4-mini-Gα_s_βγ complex at a 2.8 Å resolution, revealing the key interaction sites between A_2A_R and mini-Gα_s_βγ without ligand (Figs. 1d–f and Fig.3a). For a comprehensive understanding of our methods, we have detailed the sample preparation, cryo-EM data acquisition, and processing steps in Extended Data Figs. 3–6, and Supplementary Table 1.
Comparisons of the S4-mini-Gα_s_βγ complex with the S5-mini-Gα_s_βγ complex (PDB ID: 6GDG), as depicted in Supplementary videos 1 and 2, reveal key differences between the two states. The extracellular orthosteric binding pocket of the intermediate complex S4-mini-Gα_s_βγ is less compact than of the S5-mini-Gα_s_βγ (Fig. 3b), yet more constricted compared to the inactive state^16,17^. Transitioning from the inactive to the intermediate state S4 involves significant inward movements of all domains relative to TM1 in the extracellular domains. The shift from the intermediate to the fully activated complex results in observable movements of these domains. This is consistent with the ^19^F-qNMR study, which recorded a substantial chemical shift (~400 Hz) when the receptor transitioned from the inactive states S1–2 to intermediate S4, while the shift from S4 to S5 resulted in a minor chemical shift (~60 Hz) (Fig.3f). Closer inspection of the TM6 domain, using the ^19^F-tag labeling site as a reference, revealed a clockwise rotation consistent with the ^19^F-qNMR resonance, where the NMR signal for the S4 state is at a lower field than the S5 state. Indeed, the conformation of the intermediate S4 state is thus on the trajectory from the inactive S1–2 to the fully activated S5 states.
One of the key microswitches in the receptor is the P^5.50^I^3.40^F^6.44^ motif, which is a crucial hydrophobic switch between TM5, TM3 and TM6 and is implicated in receptor activation^1,23^. In the fully activated receptor, P^5.50^ and F^6.44^ are in cis positions, forming a strong CH-π interaction^24^, and the CH_3_ group of I^3.40^ also forms a robust CH_3_-π interaction with F^6.44^, stabilizing the fully activated conformation. In contrast, these interactions are weakened in the S4-mini-Gα_s_βγ complex because P^5.50^ and I^3.40^ are oriented away from F^6.44^ (Fig. 3d). Another key microswitch is the CW^6.48^xP motif, where P^6.50^ acts as a hinge in TM6 and facilitates the opening of the cytoplasmic cavity during activation^25^. Our cryo-EM structure clearly shows P^6.50^ in an “intermediate” position, bridging the inactive and active states (Fig. 3e). The third microswitch, the NPxxY^7.53^ on TM7, along with a conserved tyrosine Y^5.58^ in TM5, stabilizes the receptor’s active state^26–28^. As illustrated in Fig. 3g, a strong H-bond is observed between Y^5.58^ and Y^7.53^ in the S5-mini-Gαβγ complex. This interaction draws Y^5.58^ on TM5 closer to TM7, maintaining the compactness of the TM bundle in the S5-G protein complex, as shown in the animation from the S4-mini-Gα_s_βγ transitioning to the S5-mini-Gαβγ (Supplementary videos 1 and 2). Therefore, transitioning from the S4- to the S5-mini-Gα_s_βγ strengthens the H-bond interaction as Y^5.58^ moves closer to the TM7 domain, aiding in the insertion of the G protein into the cavity. Our analysis indeed supports the idea that the S4 conformation can regulate G protein signaling to some degree without transitioning to the fully activated S5 state.
Next, we examined the interfacial interaction between Cα5 helix of mini-Gα_s_ (Fig. 3c) and the transmembrane bundle comprising TM3, TM6 and TM7 domains - critical components in regulating G protein nucleotide exchange. As highlighted in Fig. 3c, the S4-mini-Gα_s_βγ adopts a slightly more open conformation than the S5-mini-Gα_s_βγ. The Cα5 helix is significantly rotated outward and retracted from the G protein-binding pocket, accompanied by the disengagement of the αN helix of the Gα_s_ protein from the H8 helix of the receptor. This retraction measures approximately 2 Å and the rotation is at a 2° spinal clockwise angle from the S5-mini-Gα_s_βγ. This motion is further detailed in Fig. 3h, which shows the primary interactions at the junction composed of TM3, TM6, TM7 domains, and the Cα5 segment from mini-Gα_s_. During the activation process, the clockwise rotation of TM6 and counterclockwise rotation of Cα5 culminate in a compact end-state complex. The transition from the S4- to the S5-mini-Gα_s_βγ entails a further insertion of the G protein and an inward rotation of the receptor domain, evidenced by the formation of the salt bridge R^7.56^-E392^Gα^ and the cation-π interaction between R^7.56^ and H^6.32^, coupled with a strengthened cation-π interaction between R^3.50^ and Y391^Gα^. However, the R^7.56^ to A^7.56^ mutation eliminates this interaction.
Molecular basis of A2AR function in the S4 state
To validate the conformational transition between the S4 and S5 states, we applied all-atom Gaussian accelerated MD (GaMD) simulations, starting from atomic structure of the S5-mini-Gα_s_βγ complex (6GDG) with a R291A mutation modeled in silico. Indeed, the R291A mutation facilitated transition of the 6GDG structure (S5 state, Fig.1b) to an energy minimum conformation that closely matches our new cryo-EM structure. We refer to this as cS4 state, where “c” stands for computational model, and here the Cα5 helix in Gα_s_ retracted from the receptor by ~2 Å. The center-of-mass distance between the receptor NPxxY motif and the last five residues of Cα5 helix increased from ~13.2 Å in the “S5” state to ~15.3 Å in the “cS4” state (Fig. 4a and Extended Data Fig. 7). As a control, GaMD simulations showed that the cWT-mini-Gα_s_βγ complex remained stable, mainly sampling one low-energy state, corresponding to the S5 (Fig.4b). These simulations independently validated the conformation of the intermediate S4 state determined from single particle cryo-EM.
Furthermore, we performed additional GaMD simulations to examine GDP release. In the cWT-Gα_s_βγ system, GDP was observed moving away from the initial binding site by up to ~12 Å repeatedly (Fig.4c). In contrast, GDP underwent significantly smaller movements up to only ~8 Å in the cR291A-Gα_s_βγ system (Fig. 4f). Free energy calculations also indicated a “GDP Released” state in the cWT-Gα_s_βγ system and only a “Partially Released” state in the cR291A-Gα_s_βγ system (Extended Data Figs. 8a–c), where the AHD domain of Gα_s_ transitioned from the “Open” to “Partially Open” conformation, with the orientation angle between the AHD and Ras-like domain decreasing to ~30° (Extended Data Figs. 8d–e)^1,29^. Together, these observations suggest that the S4 state mediated G protein has a reduced capacity for GDP release compared to the S5 state, aligning with the nucleotide exchange data shown above.
A limited nucleotide exchange model for intermediate GPCR-Gαsβγ complex
A mechanistic model is proposed to understand the rate-limited nucleotide exchange in the intermediate GPCR-G protein complex (Fig.5). The transition from the intermediate to the fully activated complex involves a conformational change at the interface, where the TM6 helix of the receptor rotates clockwise by 8° while the Cα5 helix from the Ras-like domain rotates anticlockwise by 2°. This leads to a more compact interaction between the receptor and G protein, coupled with a 2 Å uplift of the Cα5 helix. This uplift results in the disengagement of the H1 from the Cα5 helix, facilitating the separation of AHD domain from Ras-like domain and thereby exposing the bound GDP at site 2. Simultaneously, this process assists the relocation of GTP from site 1 to site 2, replacing GDP. However, in the S4-mediated G protein, the substitution of R^7.56^ with alanine (A^7.56^) results in the loss of both the intramolecular cation-π (R^7.56^-H^6.32^) interaction and the intermolecular salt bridge (R^7.56^-E392^Gα^). Instead, a partially opened AHD domain is seen (Fig.4e), which can be attributed to a weak electrostatic H-bond (Q59-T369, 4 Å)^30^ between Cα5 and H1 helices. While in the closed state of the AHD domain, an additional CH_3_-π interaction between F376 and M60 plays a significant role (Fig.4d and Figs.4g–i). Of note, in all complex structure resolved so far, the crystallographic complex structure of β2AR-Gαβγ represents the sole fully activated complex structure, in which the AHD domain with an open of 88° while all others are between 55–65°. This could be the effect from crystallographic packing in the crystallization process^31^. Collectively, these findings provide a molecular basis for the limited nucleotide exchange observed in the intermediate GPCR-G protein complex.
Discussion
Previous MD simulation has indicated that GTPγS and GDP exhibited different contact patterns with G protein^32^. A recent study also demonstrated a transition process of GTP nucleotide in a β_2_AR-regulated G protein from a “loose” interaction site to the final nucleotide-binding pocket by 2 Å^33^. Our study provides evidence for the dislocation of nucleotide, regulated by a “lift” of the Cα5 tail by 2 Å, which is essential for dislodging the AHD domain of the G protein from the Ras-like domain. This AHD open facilitates the opening of the canonical site 2, accommodating GTP for free-nucleotide exchange and hydrolysis^34^. However, the precise location of site 1 en route from the S4-Gαβγ to the S5-Gαβγ in the GPCR activation process and its specific functional role in G protein signaling remain unclear and require further characterization in the future.
GPCR activation is known to involve the transition through multi-states^4–12^. However, transient intermediate states represent higher energy substates from energy landscape perspective and are thus challenging to characterize, especially with regard to obtaining transient GPCR-G protein complexes and studying their functions. Our current study demonstrates a strategy whereby intermediate conformational states can be identified by ^19^F-qNMR, trapped by point mutation, and structurally and functionally characterized in their complexes with G proteins. Although we used A_2a_R as a model system, it is clear that such strategy of utilizing 19F-qNMR-guided cryo-EM, in which 19F-NMR serves as conformational indicator, can be broadly applied to study transient complexes of other GPCRs and proteins of interest during activation or inhibition processes. Deciphering the functions of intermediate states and their complexes enables a comprehensive understanding of GPCR signaling complexity and the exploration of alternative therapeutic approaches by targeting a specific disease-related conformation(complex). As the advance of our knowledge on structures and functions of individual conformational states and their complexes, it will become possible to design drug based on their conformational selectivity on the receptor, considering signaling bias originated from conformational bias^5^.
Methods
Plasmid construction and transformation
The full-length human A_2A_R gene, originating from construct pPIC9K_ADORA2A, was generously provided by Prof. Takuya Kobayashi (Kyoto University, Kyoto, Japan). The C-terminally truncated construct A_2A_R_316, constructed in our previous study, has an integrated FLAG tag on the N-terminus and a poly-his tag on the C-terminus^7^. Based on this construct, the mutations V229C and R291A were described elsewhere^15^. All constructs were sequenced by a facility at Eurofins genomics, with the AOX1 primer pair of PF_AOX1_ and PR_AOX1_. Freshly prepared competent cells of strain Pichia Pastoris SMD 1163 (Δhis4 Δpep4 Δprb1, Invitrogen) were electro-transformed with PmeI-HF (New England Biolabs) linearized plasmids containing different mutant genes using a Gene Pulser II (Bio-Rad). High-copy clone selection was performed using an in-house protocol described previously^8,35^. A high-yield construct was then screened by an immunoblotting assay with both anti-FLAG and anti-Poly-his for further large-scale expression screening.
Receptor expression, purification, and labeling
The screened WT* and mutants R291A, were pre-cultured on YPD [1% (w/v) yeast extract, 2% (w/v) peptone and 2% (w/v) glucose] plates containing 0.1 mg/mL G418. A single colony for each construct was inoculated into 4 mL YPD medium and cultured at 30 °C for 12 hours, then transferred into 200 mL BMGY medium [1% (w/v) yeast extract, 2% (w/v) peptone, 1.34% (w/v) YNB (yeast nitrogen base) without amino acids, 0.00004% (w/v) biotin, 1% (w/v) glycerol, 0.1 M PB (phosphate buffer) at pH 6.5] and cultured at 30 °C for another 30 h. The cells were then transferred into 1 L of BMMY medium [1% (w/v) yeast extract, 2% (w/v) peptone, 1.34% (w/v) YNB without amino acids, 0.00004% (w/v) biotin, 0.5% (w/v) methanol, 0.1 M phosphate buffer at pH 6.5, 0.04% (w/v) histidine and 3% (v/v) DMSO, 10 mM theophylline] at 20 °C. 0.5% (v/v) methanol was added every 12 h. 60 h after induction by methanol, cells were harvested for purification.
The cell pellets were collected by centrifugation at 4,000 ×g for 20 minutes and washed one time with washing buffer (50 mM HEPES, 10% glycerol, pH 7.4) before the addition of breaking buffer (50 mM HEPES, pH 7.4, 100 mM NaCl, 2.5 mM EDTA, 10% glycerol) in a ratio of 4:1 (buffer: cells). The resuspended cell pellets were subject to disruption 3 times using a Microfluidizer at a pressure of 20,000 psi. Intact cells and cell debris were separated by low-speed centrifugation (8,000 ×g) for 30 minutes. The supernatant was collected and centrifuged at 100,000 ×g for 2 h, and the precipitated cell membrane was then immediately dissolved in membrane lysis buffer (50 mM HEPES, pH 7.4, 100 mM NaCl, 0.5% LMNG-3 (Lauryl Maltose Neopentyl Glycol) and 0.1% CHS (cholesteryl hemisuccinate)) with rotation 2 h or overnight at 4 °C until the membrane was dissolved. Subsequently, Talon resin (Clontech) was added to the solubilized membranes and incubated for at least 2 h or overnight under gentle agitation.
The A_2A_R-bound Talon resin was washed twice with a buffer of 50 mM HEPES, pH 7.4, 100 mM NaCl, 0.02% LMNG-3 and 0.002% CHS and resuspended in the same buffer. The A_2A_R-bound Talon resin was then resuspended in buffer made of 50 mM HEPES, pH 7.4, 100 mM NaCl, 0.02% LMNG-3 and 0.002% CHS, and combined with 10–20 fold excess of the NMR label (2-bromo-N-(4-(trifluoromethyl)phenyl)acetamide, BTFMA, Apollo Scientific, Stockport, UK) under gentle agitation overnight at 4 °C. Another aliquot of NMR label was then added and incubated for an additional 6 h to ensure complete labeling. The A_2A_R-bound Talon resin was washed in a disposable column extensively with buffer containing 50 mM HEPES, pH 7.4, 100 mM NaCl, 0.02% LMNG-3 and 0.01% CHS, and apo A_2A_R was then eluted from the Talon resin with 50 mM HEPES, pH 7.4, 100 mM NaCl, 0.02% LMNG-3 and 0.01% CHS, 250 mM imidazole and concentrated to a volume of 5 mL. The XAC-agarose gel and A_2A_R were then incubated together for 2 h under gentle agitation. The functional A_2A_R was eluted with 50 mM HEPES, pH 7.4, 0.02% LMNG-3, 0.002% CHS, 100 mM NaCl, 20 mM theophylline. The eluted samples were concentrated to 1 mL by centrifugal filtration (MWCO, 3.5 KDa), and an extensive dialysis was performed to remove the XAC in the sample. The functional apo A_2A_R was then prepared for NMR. All receptors described in this manuscript were purified using poly-his resin followed with a ligand-column, in which the A_2A_R antagonist xanthine amine congener (XAC) was conjugated to Affi-Gel 10 activated affinity media.
Preparation of mini-Gαsβγ heterotrimer
The plasmid for mini-Gα_s_399 was generously provided by Drs. Christopher G. Tate from MRC Laboratory of Molecular Biology, Cambridge and Javier García Nafría from Institute for Biocomputation and Physics of Complex Systems (BIFI) and Laboratorio de Microscopías Avanzadas (LMA), University of Zaragoza. It was expressed in the E. coli strain BL21(DE3). Cells were collected by centrifugation at 4,000 ×g for 20 min and lysed by sonication. After another centrifugation, the supernatant was purified by Talon resin. The sample was loaded into a HiLoad16/60 column to obtain purified mini-Gα_s_ protein. The purified protein was concentrated to 3 mg/mL and flash frozen in liquid nitrogen and stored at −80 °C for further use. The expression and purification of the respective components and assembly to make the complex containing mini-Gαs_β_1_γ_2, and the preparation of nanobody Nb35, were all performed following the protocols described previously^1,36,37^.
19F NMR experiments
NMR samples typically consisted of 280–300 μL volumes with 20–50 μM A_2A_R in 50 mM HEPES buffer and 100 mM NaCl, doped with 10% D_2_O. The receptor was stabilized in 0.02% LMNG-3 and 0.002% CHS. All ^19^F NMR experiments were performed on a 600 MHz Varian Inova spectrometer using a ^19^F dedicated resonance probe. Typical experimental setup included a 16 μs 90° excitation pulse, an acquisition time of 200 ms, a spectral width of 15 kHz, and a repetition time of 1 s. Most spectra were acquired with 15,000–50,000 scans. Processing typically involved zero filling, and exponential apodization equivalent to 15 Hz line broadening.
GTPase hydrolysis assay
The GTPase hydrolysis assay was analyzed using a modified protocol of the GTPase-Glo^™^ assay (Promega)^38^. The reaction was started by mixing 300 nM Gα_s_βγ with the purified receptors in varied concentrations with a final volume of 10 μL in the buffer containing 50 mM HEPES, pH 7.4, 100 mM NaCl, 0.002% CHS, 0.02% LMNG-3. For the GTP hydrolysis capacity of the S4 state as a function of ligand measurement, 5x and 25x ligand compared to receptor concentration was added. After 30 min incubation at room temperature, 10 μL 2xGTP-GAP solution containing 10 μM GTP, 1 mM DTT and the cognate GAP was added to each well, followed with a 120 min incubation at room temperature. For the Michaelis–Menten constant measurement, the 2xGTP-GAP solution containing 5–50 μM GTP was used. Then, 20 μL reconstituted GTPase-Glo^™^ reagent containing 5 μM ADP was added to each sample and incubated for another 30 min at room temperature with shaking. Luminescence was measured following the addition of 40 μL detection reagent and incubation for 10 min at room temperature using a BioTEK-Flx800 plate reader at 528±20 nm. Determine the amount of GTP consumed in a biochemical reaction by referencing a standard curve that relates light units (RLU) indicative of product formation to GTP concentration. Using this data, calculate the rate of the enzymatic reaction (velocity, v) by applying the Michaelis-Menten equation: v = V_max_/(1+(K_m_/[S])). To facilitate the determination of Vmax and Km, construct a Lineweaver-Burk plot, which linearizes the relationship by graphing the reciprocal of the velocity (1/v) against the reciprocal of the substrate concentration (1/[S]). The calculations of initial rates were performed at 1.08 min in the linear reaction phase of catalysis. Analysis of data was performed by Excel and GraphPad Prism^®^ 9.0.
BODIPY-FL-GTP, BODIPY-FL-GDP binding, and nucleotide exchange
The nucleotide-binding assay utilized BODIPY-FL-GTP and BODIPY-FL-GDP, from Invitrogen^™^, each supplied as a mixture of two isomers with the fluorophore attached at either the 2′ or 3′ position on the ribose ring. BODIPY-FL-GTP could be hydrolyzed to BODIPY-FL-GDP. To form the GPCR-G protein complex, a solution containing 2 μM G protein and 100 μM receptor was incubated for 30 minutes at 22°C. The fully activated state of the WT* receptor was achieved by supplementing the mixture with 10 mM NECA. Assays were conducted at 22°C using 96-well half-area microtiter plates in a BioTek plate reader, with excitation at 475 nm and emission measured at 528 nm. The assay buffer comprised 20 mM HEPES (pH 7.4), 1 mM EDTA, and 10 mM MgCl_2_, supplemented with 0.01% LMNG-3 for protein stability. Initial kinetic data were acquired for 100 nM BODIPY-FL-GTP/BODIPY-FL-GDP in the absence of G protein for 70 seconds to establish a baseline fluorescence intensity. Subsequently, 200 nM heterotrimer G proteins, with or without the receptor, were added, and mixing was rapidly performed in the fluorescence cuvette. Data collection proceeded uninterrupted, and resulting kinetics spectra were plotted and fitted to a one-phase association function using GraphPad Prism 9.0. For the GTP-GDP exchange assay, the GPCR-G protein complex was formed as described, followed by incubation with 100 nM BODIPY-FL-GDP for 2 hours. Baseline fluorescence intensity was measured for 70 seconds using the plate reader, after which 1 μM GTP (in a concentration of 10×GDP), procured from Invitrogen^™^, was added to facilitate the exchange of BODIPY-FL-GDP to GTP. All experiments were repeated three times, and resulting kinetics spectra were analyzed using GraphPad Prism 9.0.
Gaussian accelerated molecular dynamics (GaMD)
GaMD is an enhanced sampling method that works by adding a harmonic boost potential to reduce the system energy barriers^39,40^. When the system potential is lower than a reference energy E, the modified potential of the system is calculated as:
where k is the harmonic force constant. The two adjustable parameters E and k are automatically determined on three enhanced sampling principles. First, for any two arbitrary potential values and found on the original energy surface, if , ΔV should be a monotonic function that does not change the relative order of the biased potential values; i.e., . Second, if , the potential difference observed on the smoothened energy surface should be smaller than that of the original; i.e., . By combining the first two criteria and plugging in the formula of and, we obtain
Where V_min_ and V_max_ are the system minimum and maximum potential energies. To ensure that Eq. 2 is valid, k has to satisfy: k≤1/V_max_-V_min_. Let us define: k=k_0_∙1/(V_max_-V_min_), then 0<k_0_≤1. Third, the standard deviation (SD) of ΔV needs to be small enough (i.e. narrow distribution) to ensure accurate reweighting using cumulant expansion to the second order: σ_ΔV_=k(E-V_avg_),σ_V_≤σ_0_, where V_avg_ and σ_V_ are the average and SD of ΔVwith σ_0_ as a user-specified upper limit (e.g., 10k_B_T) for accurate reweighting. When E is set to the lower bound E=V_max_ according to Eq. 2, k_0_ can be calculated as
Alternatively, when the threshold energy E is set to its upper bound E=V_min_+1/k, k_0_ is set to:
If is calculated between 0 and 1. Otherwise, k_0_ is calculated using Eq. 3.
System setup and simulation analysis
The cryo-EM structure of wild-type NECA-bound A_2A_R bound by mini-Gα_s_ (PDB ID: 6GDG^18^) was used for setting up simulation systems of the wild-type NECA-bound cWT*-A_2A_R-mini-Gα_s_βγ, apo cR291A-mini-Gα_s_βγ, apo cWT*-A_2A_R-Gα_s_βγ and apo cR291A-Gα_s_βγ. The missing residues in the extracellular loop 2 (ECl2) and intracellular loop 3 (ICL3) of the receptor were modelled with Swiss-Modeller^41^. For the apo cR291A-mini-Gα_s_βγ, the simulation structure was generated by mutanting the correspoding R291A and V229C in the wild-type system and deleting the agonist NECA. To build the wild-type apo A_2A_R bound by the full-length of Gs protein (apo cWT*-A_2A_R-Gα_s_βγ), the Swiss-Modeller^41^ was used to build the Gs protein with a geometry and orientation similar to the Gs protein in the fully active state of the β_2_AR-Gs complex (PDB ID: 3SN6^1^). The coordinates of the GDP and Mg^2+^ were obtained by aligning the Ras domain of the crystal structure of Gs-bound GDP (PDB ID: 6AU6^42^) to the modelled full-length Gs protein bound by the A_2A_R.The simulation structure of the apo cR291A-Gα_s_βγ was generated by substituting residues Arg291 and Val232 in the wild-type system (apo cWT*-A_2A_R-Gα_s_βγ) with Ala and Cys, respectively.
VMD was used to insert the NECA-bound cWT*-A_2A_R-mini-Gα_s_βγ, apo cR291A-mini-Gα_s_βγ, apo cWT*-A_2A_R-Gα_s_βγ, and apo cR291A-Gα_s_βγ complex into POPC (palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine) lipids to prepare simulation systems. In each simulation system, the protein and lipid bilayer were solvated with TIP3P water molecules in a box of 11.2 nm × 13.1 nm × 14.6 nm with the periodic boundary condition. The system charge was neutralized with 150 mM NaCl. The CHARMM36m parameter set^43–45^ was used for the proteins and lipids, and Guanosine diphosphate (GDP). Force field parameters of the NECA agonist were obtained from the ParamChem web server^46^. The four simulation systems were first energy minimized for 5,000 steps with constraints on the heavy atoms of the proteins and phosphor atom of the lipids. The hydrogen-heavy atom bonds were constrained using the SHAKE algorithm and the simulation time step was set to 2.0 fs. The particle mesh Ewald (PME) method^47^ was employed to compute the long-range electrostatic interactions and a cutoff value of 9.0 Å was applied to treat the non-bonded atomic interactions. The temperature was controlled using the Langevin thermostat with a collision frequency of 1.0 ps^−1^. The system was then equilibrate using the constant number, volume, and temperature (NVT) ensemble at 310K for 250 ps and under the constant number, pressure, and temperature (NPT) ensemble at 310 K and 1 bar for another 1 ns with constraints on the heavy atoms of the protein, followed by 10 ns short conventional MD (cMD) without any constraint.
The GaMD module implemented in the GPU version of AMBER22^48–50^ was then applied to perform the simulations of NECA-bound cWT*-A_2A_R-mini-Gα_s_βγ, apo cR291A-mini-Gα_s_βγ, apo cWT*-A_2A_R-Gα_s_βγ, and apo cR291A-Gα_s_βγ. GaMD simulations included a 8-ns short cMD run used to collect the potential statistics for calculating GaMD acceleration parameters, an 56-ns GaMD equilibration after adding the boost potential, and finally three independent 2,000-ns GaMD production simulations with randomized initial atomic velocities for the systems of the NECA-bound cWT*-A_2A_R-mini-Gα_s_βγ and apo cR291A-A_2A_R-mini-Gα_s_βγ complex, and 3,000-ns GaMD production simulations for the systems of apo cWT*-A_2A_R-Gα_s_βγ and apo cR291A-Gα_s_βγ with randomized initial atomic velocities. The average and SD of the system potential energies were calculated every 800,000 steps (1.6 ns). All GaMD simulations were performed at the “dual-boost” level by setting the reference energy to the lower bound. One boost potential was applied to the dihedral energetic term and the other to the total potential energetic term. The upper limit of the boost potential SD, σ_0_ was set to 6.0 kcal/mol for both the dihedral and the total potential energetic terms.
For each simulation system, all three GaMD production trajectories were combined together for analysis with CPPTRAJ^51^. The distance between the NPxxY motif of the receptor and the last five residues of Gα_s_ α5 helix (A_2A_AR:NPxxY-Gα_s_:α5 distance), the distance between the receptor TM3 and TM6 intracellular ends (measured by the distance between the Cα atoms of receptor residues Arg102^3.50^ and Glu228^6.30^), the root-mean-square derivation of GDP (GDP RMSD) relative to the simulation starting structure were selected as reaction coordinates. The angle between the Ras domain and α helical domain (AHD) was used as another reaction coordinate to indicate their relative orientation, which was defined by the two vectors of Gα_s_ AHD and Gα_s_ Ras domain. Vector 1 went through the Gα_s_ AHD and A161 centers and vector 2 went through Gα_s_ Ras domain and E299 centers. The PyReweighting^52^ toolkit was applied to reweight GaMD simulations to recover the original free energy profiles of the simulation systems. 2D free energy profiles were computed using the combined trajectories from all the three independent GaMD simulations for each system with the A_2A_R:NPxxY-Gα_s_:α5 distance, TM3-TM6 distance, GDP RMSD and the angle between the Ras domain and AHD as reaction coordinates. A bin size of 1.0 Å was used for the A_2A_R:NPxxY-Gα_s_:α5 distance, TM3-TM6 distance and GDP RMSD. A bin size of 6.0° was used for the angle between the Ras domain and AHD as reaction coordinate. The cutoff was set to 500 frames for 2D free energy calculations.
Preparation of the A2AR-mini-GSβ1γ2-Nb35 complex
A_2A_R, mini-G_S_-β_1_γ_2_ and Nb35 were mixed in a molar ratio of 1:2:4, to yield a final complex concentration of 1 mg/mL. To this mixture, 0.1 U of apyrase was added, followed by an overnight incubation at 4°C. The mixture was then concentrated with a 100 kDa MWCO Amicon filter and injected onto a Superdex200 Increase 10/300 GL gel filtration column equilibrated with buffer (50 mM HEPES, 100 mM NaCl, 0.002% LMNG-3, 0.0002% CHS (w/v)). Monodisperse fractions were concentrated with a 100 kDa MWCO Amicon filter immediately prior to cryo-EM grid preparation.
Negative staining of the complex was performed with 0.75% uranium formate, following an established protocol^53^.Grids were examined using an FEI T12 microscope operated at 120 kV, and images were recorded using a 4K × 4K charge-coupled device (CCD) camera (UltraScan 4000, Gatan).
Cryo-EM sample preparation and data acquisition
Freshly prepared A_2A_R-mini-G_S_-β_1_γ_2_-Nb35 complex at a final concentration of 1 mg/mL, was applied to glow-discharged gold grids coated with either holey carbon film (Quantifoil, 300 mesh 1.2/1.3, Au) or holey gold film (UltrAuFoil, 300, mesh 1.2/1.3). These grids were then plunge-frozen using a Vitrobot Mark IV with a blotting time of 4 s and blotting force of 0, at 4 °C and 100% humidity. Grids were subsequently examined and screened using an FEI Tecnai Arctica operated at 200 kV and equipped with an XFEG and a Gatan K3 camera. Cryo-EM data collection was performed on a Titan Krios at the UCSF Cryo-EM Center for Structural Biology, operated at an acceleration voltage of 300 kV, equipped with an XFEG, a BioQuantum energy filter (slit width set to 20 eV) and a K3 camera (Gatan).
All cryo-EM datasets were collected using SerialEM^54^. A multishot collection (3×3 arrays) was employed, incorporating beam-tilt compensation and maximum image shift is 3.5 micros. All images were acquired with a nominal magnification of 105 K, resulting in a super-resolution pixel size of 0.4175 Å (physical pixel size of 0.835 Å). Defocus range was set from −0.8 μm to −1.8 μm. A total of 8,805 images were collected, each was dose-fractionated into 80 movie frames with a total exposure time of 2.024 s, resulting in a total fluence of approximately 47.7 electrons per Å^2^. Data collection statistics are shown in Supplementary Table 1.
Image process
A total of 8,805 movie stacks were motion-corrected, dose-weighted, and binned by Fourier cropping to the physical pixel size of 0.835 Å on-the-fly using MotionCor2^55^. Motion-corrected, does-weighted sums were used for contrast transfer function (CTF) determination and resolution estimation in cryoSPARC^56^. 424,536 particles were picked from randomly selected 500 micrographs using cryoSPARC blob picker with a diameter of 80–150 Å. These particles were subjected to ab-initio reconstruction and multi-round heterogenous refinement. One class with distinct features of A_2A_R bound to mini-Ga_s_β_1_γ_2_-Nb35 was identified, from which 16 different projection images were created for template picking, yielding 7,282,317 particles from all micrographs.
After removing junk particles by extensive 2D classifications, 2,328,810 particles were selected for ab-initio reconstruction and multi-round heterogenous refinement. One distinct class of 307,568 particles was identified for further non-uniform refinement of the A_2A_R-mini-Ga_s_β_1_γ_2_-Nb35 complex, yielding a reconstruction with a global resolution of 3.04Å. This particle stack was then exported to RELION^57^ for multiple rounds of 3D refinement (initial low-pass filter: 10 Å; mask diameter: 360 Å; reference mask: no) to produce a new reconstruction, from which a mask without the detergent micelle was generated by using the segment map function in Chimera^58^ (initial threshold: 0.0001; extend_inimask: 4; width_soft_edge: 4). Particle subtraction function in RELION was applied by using this mask to generate a micelle removed particle stack.
Next, we applied 3D classification to the micelle subtracted particles with a reference model without micelle (reference mask: generated last step; initial low-pass filter: 10 Å; mask diameter: 260 Å; regularisation parameter T: 3; number of iterations: 50; number of classes: 5; perform local angular searches: no). A new reconstruction of the entire A_2A_R-mini-G_S_-β_1_γ_2_-Nb35 complex from 71,547 particles was identified for further refinement in RELION (initial low-pass filter: 3.5 Å; mask diameter: 260 Å; angular sampling interval: 1.8°; local search from auto-sampling: 0.9°). A final 3D reconstruction was calculated in cisTEM^59^ with default settings and without further refinement. The numeric resolution was determined from Fourier Shell Correction (FSC) using criterion of FSC = 0.143^60^. The final map was sharpened by DeepEMhancer^61^ and used for model building and figure generation.
Model building
For model building, the initial model was generated by fitting the existing coordinates of the activated state of A_2A_R-mini-G_S_-β_1_γ_2_-Nb35 complex (PDB: 6GDG) into our cryo-EM density maps using ChimeraX^62^. Discrepancies between the initial model and the density maps were then manually built and refined ISOLDE^63^ and coot^64^. Subsequent refinements were performed in Phenix^65^ with secondary structure constraints. The model was validated by wwPDB validation server^66^ and no major issues was reported. A summary of the parameters used in data collection and model building is provided in Supplementary Table 1.
Data representation
All statistical tests such as GTP hydrolysis assessments were conducted using GraphPad Prism 9.0. The central point of all data points gives the mean value with s.d. for all data unless otherwise specified. Visualization of the atomic models (figures and videos) were made using UCSF ChimeraX and PyMoL (The PyMOL Molecular Graphics System, Version 2.0 Schrödinger, LLC.)
Extended Data
Conformational profiles of the mutant R291A and its complexes probed by 19F-qNMR.(A) and (B), Deconvoluted conformational profiles probed by 19F-qNMR as a function of mini-Gαsβγ and Gαsβγ (S4-Gαβγ), in reference to the WT-mini-Gαsβγ +NB35 (S5-Gαsβγ), representing the conformational profile for the structure PDB ID: 6GDG.*
Both linewidth broadening of 19F-qNMR resonance for S4 intermediate and native-PAGE indicated the complex formation between the S4 and G protein.a, Linewidths of S4 resonance for R291A, R291A-Gαsβγ, and R291A-mini-Gαsβγ-NB35. b, Native-PAGE for R291A and Gαsβγ interaction.
Preparation of A2AR sample and cryo-EM screening.a, Size exclusion chromatography of purified A2AR-R291A with and without mini-Gsβγ+NB35. b, SDS-PAGE of the indicated fraction, corresponding to R291A+mini-Gsβγ+NB35. c, Negative staining of the R291A+mini-Gsβγ+NB35 complex. d, Representative cryo-EM micrograph of the complex. e, Representative 2D average image of the complex.
Cryo-EM data and image processing flow.Schematic flow-chat representing the image processing approach for A2AR-R291A. Thumbnail images of each 3D class or refinement are shown along with global GS-FSC resolution in black, particles counts in blue, and dashed black boxes to indicate selected 3D classes. Cryo-EM map (red box) and atomic model are used in main figures to present A2AR-R291A features.
Resolution estimation and atomic model quality.a, Fourier shell correlation curve with resolution determined by the ‘gold-standard’ FSC. b, Model-to-map FSC plots calculated by PHENIX between the map and the model. c, Cryo-EM map colored by local resolution estimates using RELION with resolution scale bar (Å). d, Histogram representation of the Euler angle distribution from the final particles used in the reconstruction.
Representative cryo-EM densities from selected structural features of A2AR.a, Densities of all transmembrane helices. b, Representative densities of mini-G. c, Cryo-EM densities of V229C and R291A. d, Representation of surface hydrophobicity.
GaMD simulations for cR291A-mini-Gαsβγ and cWT-mini-Gαsβγ.a-b, Time course of the center-of-mass (COM) distance between the receptor NPxxY motif in TM7 and the Cα5 helix (A2AR:NPxxY-Gαs:Cα5 distance) calculated from the GaMD simulations of cR291A-mini-Gαsβγ (a) and cWT*-mini-Gαsβγ (b) complex systems. c-d, Representative low-energy conformations of the cR291A-mini-Gαsβγ (c) and (d) cWT*-mini-Gαsβγ complex identified from the GaMD simulations. The receptor NPxxY motif and the last five residues of Gα5 helix were colored in orange.*
The behaviors of AHD domain in cS4 and cS5 mediated G protein.a, The AHD angle is illustrated by comparing the GaMD simulation conformation with the inactive conformation of Gαs (PDB: 6EG8); Vector 1 goes through centers of the Gαs AHD and Ras-like domain of the inactive conformation (6EG8). Vector 2 goes through the centers of the Gαs AHD and Ras-like domain of the GaMD simulation conformations after aligning the Ras-like domain to the inactive conformation of Gαs. b-c, The time course of the AHD angle calculated from the GaMD simulations for the apo cR291A-Gαsβγ (b) and apo cWT-A2AR-Gαsβγ (c) complexes. d-e, 2D free energy profiles of the apo cR291A-Gαsβγ (d) and apo cWT*-Gαsβγ (e) complexes calculated from GaMD simulations regarding the GDP RMSD and the AHD angle.*
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Rasmussen S. G. Crystal structure of the beta 2 adrenergic receptor-Gs protein complex. Nature 477, 549–555 (2011). 10.1038/nature 1036121772288 PMC 3184188 · doi ↗ · pubmed ↗
- 2Ye L., Wang X., Mc Farland A. & Madsen J. J. (19)F NMR: A promising tool for dynamic conformational studies of G protein-coupled receptors. Structure 30, 1372–1384 (2022). 10.1016/j.str.2022.08.00736130592 · doi ↗ · pubmed ↗
- 3Zhang Y. Cryo-EM structure of the activated GLP-1 receptor in complex with a G protein. Nature 546, 248–253 (2017). 10.1038/nature 2239428538729 PMC 5587415 · doi ↗ · pubmed ↗
- 4Hilger D. The role of structural dynamics in GPCR-mediated signaling. FEBS J 288, 2461–2489 (2021). 10.1111/febs.1584133871923 · doi ↗ · pubmed ↗
- 5Wang X., Mc Farland A., Madsen J. J., Aalo E. & Ye L. The potential of (19)F NMR application in GPCR biased drug discovery. Trends Pharmacol Sci 42, 19–30 (2021). 10.1016/j.tips.2020.11.00133250272 · doi ↗ · pubmed ↗
- 6Heo L. & Feig M. Multi-state modeling of G-protein coupled receptors at experimental accuracy. Proteins (2022). 10.1002/prot.26382 PMC 956104935510704 · doi ↗ · pubmed ↗
- 7Ye L., Van Eps N., Zimmer M., Ernst O. P. & Prosser R. S. Activation of the A 2A adenosine G-protein-coupled receptor by conformational selection. Nature 533, 265–268 (2016). 10.1038/nature 1766827144352 · doi ↗ · pubmed ↗
- 8Ye L., Orazietti A. P., Pandey A. & Prosser R. S. High-efficiency expression of yeast-derived G-protein coupled receptors and (19)F labeling for dynamical studies. Methods Mol Biol 1688, 407–421 (2018). 10.1007/978-1-4939-7386-6_1929151220 · doi ↗ · pubmed ↗