Characterization of temperature-sensitive Schizosaccharomyces pombe mutants in the septation initiation network Spg1 GTPase
Anna Bowman Fletcher, Lesley A. Turner, Liping Ren, Alaina H. Willet, Kathleen L. Gould

TL;DR
This study examines temperature-sensitive mutations in the Spg1 GTPase of fission yeast, which is crucial for cell division.
Contribution
The paper provides a detailed characterization of temperature-sensitive spg1 alleles and their interactions with other SIN alleles.
Findings
Temperature-sensitive spg1 alleles were identified and their mutations characterized.
Growth defects of these alleles were assessed under different conditions.
Interactions between the spg1 alleles and other SIN alleles were evaluated.
Abstract
The Schizosaccharomyces pombe GTPase, Spg1 , activates the septation initiation network (SIN) protein kinase cascade to trigger septation. In the absence of functional Spg1 , cells fail cytokinesis and become multinucleate. In this study, we characterize a set of temperature-sensitive spg1 alleles isolated in the 1990s. We identify the mutations within each new and previously characterized allele, characterize the extent of relative growth defects, and assess their interaction with other SIN alleles.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
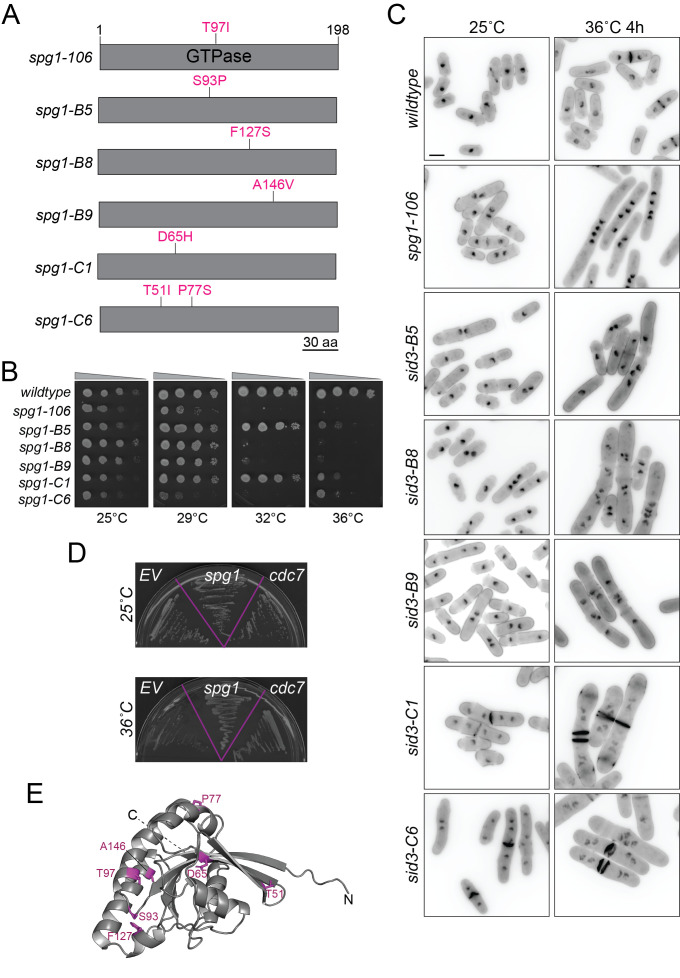 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal and yeast genetics research · Bacterial Genetics and Biotechnology · RNA Research and Splicing
Description
In the yeast Schizosaccharomyces pombe , cytokinesis is enabled by an actin- and myosin-based cytokinetic ring (CR) coupled to formation of a division septum (Cheffings et al., 2016; Glotzer, 2017; Mangione and Gould, 2019) . A signaling pathway named the septation initiation network (SIN) is essential for normal CR assembly and maintenance, constriction, and septation (reviewed in Cullati and Gould, 2019; Simanis, 2015; Xiao and Dong, 2021). At the top of the SIN cascade is the Spg1 GTPase (Schmidt et al., 1997; Sohrmann et al., 1998) . When Spg1 is inactivated, SIN signaling and cytokinesis fail, and this produces inviable cells that are elongated and multinucleated. Reciprocally, enhanced Spg1 activity induces the formation of multiple CRs and septa in the absence of nuclear division and is also lethal.
Here, we examine six previously uncharacterized temperature-sensitive strains obtained in a screen for mutants that fail cytokinesis and septation (Balasubramanian et al., 1998) . By genetic analysis, these strains were found to be in the same complementation group. Further, none of these mutants segregated in genetic crosses away from spg1-106 and it was concluded that these six strains were additional mutant alleles of * spg1 * (Balasubramanian et al., 1998) .
To further investigate the nature of these alleles, the * spg1 * open reading frame was amplified from each of the six strains and sequenced to determine what mutations were present. In each case, one or more mutations were detected ( Figure 1A ). Two alleles ( spg1-B5 and spg1-B10 ) had the same mutation causing a serine to proline change at residue 93 (S93P). Two other alleles ( spg1-B9 and spg1-B11 ) contained an alanine to valine substitution at position 146 (A146V). A fifth allele ( spg1-C1 ) had a change at residue 65 from aspartic acid to histidine (D65H). A sixth allele ( spg1-C6 ) contained a threonine to isoleucine change at residue 51 and a proline to serine substitution at residue 77. Finally, we also sequenced the spg1-106 and spg1-B8 open reading frames because the mutations in these alleles had not previously been reported and found a threonine to isoleucine change at position 97 in spg1-106 and a phenylalanine to serine change at position 127 in spg1-B8 ( Figure 1A ). Thus, there are now six known and distinct temperature-sensitive mutant alleles of * spg1 * .
We characterized the four new alleles further (eliminating the duplicative spg1-B10 and spg1-B11 alleles from further analysis) by first determining the range of temperature-sensitivity of each by spotting at a variety of temperatures. All temperature-sensitive alleles grew less than wildtype at 36°C. At 32°C, most alleles grew less than wildtype except for spg1-B5 and spg1-C1, and spg1-106 and spg1-C6 also showed reduced growth at 29°C ( Figure 1B ). Interestingly, spg1-C6 reproducibly grew better at 36˚C than 32˚C ( Figure 1B ). To visualize the cell phenotypes, we examined each mutant by staining for nuclei and septa after the cells were grown at 25°C and then shifted to 36˚C for 4 hours. We found that wildtype, spg1-106 , spg1-B8, *spg1-B5 * and spg1-B9 looked similar to wildtype at 25°C, but at 36°C the cells were multinucleated with no septum present ( Figure 1C ), as has been previously reported for spg1-106 (Balasubramanian et al., 1998) . Interestingly, spg1-C1 and spg1-C6 , even at 25°C, had many multinucleated cells with one medial septum. Additionally, at 36°C most cells were multinucleated with 1 or 2 medial septa ( Figure 1E ). We conclude that all * spg1 * alleles have defects in cell division.
We next tested if each new mutant could be rescued by wildtype * spg1 ^+^ * . Indeed, a * spg1 ^+^ * genomic clone contained within pUR19 (Barbet et al., 1992) rescued the temperature-sensitive growth of all four mutants, illustrated by the rescue of spg1-C1 in Figure 1D . The spg1-106 allele can also be rescued by over-expression of Cdc7 , a protein kinase that it activates for SIN signaling (Balasubramanian et al., 1998) . We found that all four mutants were also rescued by over-expression of Cdc7 , illustrated by the rescue of spg1-C1 in Figure 1D .
We also modeled the encoded mutations onto the AlphaFold2 predicted Spg1 protein structure (Jumper et al., 2021; Varadi et al., 2022) ( Figure 1F ). It will be interesting to investigate if the alleles with distinct phenotypes from spg1-106 ( *spg1-C1 * and spg1-C6 ) are simply milder alleles or if perhaps they have different molecular defects. Deciphering if GTP binding, GTP hydrolysis, partner binding or a different function are disrupted in the proteins encoded by these alleles will be exciting to determine in future studies.
Methods
Yeast methods
S. pombe strains were grown in yeast extract (YE) and standard S. pombe mating, sporulation, and tetrad dissection techniques were used to construct new strains (Moreno et al., 1991) .
Molecular biology methods
Plasmids were constructed using standard molecular biology techniques. * spg1 * alleles were sequenced by generating a PCR product with an oligonucleotide 72 bp upstream of the start site (CGAACGAGGCTTCTTATCCT) and 47 bp downstream of the stop codon (ACCCACACTCCTTTATCATG) (Integrated DNA technologies). The PCR product was sequenced with an additional forward oligonucleotide at 33 bp upstream of the * spg1 * start site (ATCTACTGCCGGGGTTCAAG).
Microscopy and image analysis
Strains for fixed-cell imaging experiments were grown at 25°C in YE and then shifted to 36°C for 3 hours. Cells were fixed with 70% ethanol for DAPI and methyl blue (MB) staining as described previously (Roberts-Galbraith et al., 2009) . Images were acquired using a Zeiss Axio Observer inverted epifluorescence microscope with Zeiss 63× oil (1.46 NA) objective and captured using Zeiss ZEN 3.0 (Blue edition) software. A singular medial Z slice was obtained. All images were further processed using ImageJ (Schindelin et al., 2012) .
Reagents
The strains used in this study and their genotypes are listed below.
Strain Genotype Source
KGY1038 * spg1-B5 ura1 leu1-32 mam2::LEU2 ade6-M216 h90 (Balasubramanian et al., 1998) *
KGY1054 * spg1-B9 ura1 leu1-32 mam2::LEU2 ade6-M216 h90 (Balasubramanian et al., 1998) *
KGY1044 * spg1-C1 ura1 leu1-32 mam2::LEU2 ade6-M216 h90 (Balasubramanian et al., 1998) *
KGY1048 * spg1-C6 ura1 leu1-32 mam2::LEU2 ade6-M216 h90 (Balasubramanian et al., 1998) *
KGY246 ade6-M210 leu1-32 ura4-D18 * h ^-^ * Lab stock
KGY8016 * spg1-B8 ade6-M210 h ^-^ * Lab stock
KGY1103 * spg1-106 ade6-M21X ura4-D18 leu1-32 h ^-^ * Lab stock
KGY4491-2 * spg1-B5 ade6-M21X leu1-32 ura4D-18 h ^-^ * This study
KGY4683-2 * spg1-B9 ade6-M21X leu1-32 ura4D-18 h ^-^ * This study
KGY4879-2 * spg1-C1 ade6-M21X leu1-32 ura4D-18 h ^+^ * This study
KGY5068-2 * spg1-C6 ade6-M21X leu1-32 ura4D-18 h ^-^ * This study
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Balasubramanian, M. K., Mc Collum, D., Chang, L., Wong, K. C., Naqvi, N. I., He, X., Sazer, S., Gould, K. L. 1998. Isolation and characterization of new fission yeast cytokinesis mutants. Genetics. 149: 1265-75. 1626.10.1093/genetics/149.3.1265 PMC 14602339649519 · doi ↗ · pubmed ↗
- 2Barbet, N., Muriel, W. J., Carr, A. M. 1992. Versatile shuttle vectors and genomic libraries for use with Schizosaccharomyces pombe. Gene. 114: 59-66. 789.10.1016/0378-1119(92)90707-v 1587485 · doi ↗ · pubmed ↗
- 3Cheffings TH Burroughs NJ Balasubramanian MK 201688 Actomyosin Ring Formation and Tension Generation in Eukaryotic Cytokinesis.Curr Biol 26150960-9822 R 719R 73710.1016/j.cub.2016.06.07127505246 · doi ↗ · pubmed ↗
- 4Cullati SN Gould KL 201912 Spatiotemporal regulation of the Dma 1-mediated mitotic checkpoint coordinates mitosis with cytokinesis.Curr Genet 6530172-808366366810.1007/s 00294-018-0921-x 30600396 PMC 6511297 · doi ↗ · pubmed ↗
- 5Glotzer M 2017103 Cytokinesis in Metazoa and Fungi.Cold Spring Harb Perspect Biol 91010.1101/cshperspect.a 02234328007751 PMC 5630000 · doi ↗ · pubmed ↗
- 6Jumper J Evans R Pritzel A Green T Figurnov M Ronneberger O Tunyasuvunakool K Bates R Žídek A Potapenko A Bridgland A Meyer C Kohl SAA Ballard AJ Cowie A Romera-Paredes B Nikolov S Jain R Adler J Back T Petersen S Reiman D Clancy E Zielinski M Steinegger M Pacholska M Berghammer T Bodenstein S Silver D Vinyals O Senior AW Kavukcuoglu K Kohli P Hassabis D 2021715 Highly accurate protein structure prediction with Alpha Fold.Nature 59678730028-083658358910.1038/s 41586-021-03819-234265844 PMC 8371605 · doi ↗ · pubmed ↗
- 7Mangione MC Gould KL 2019617 Molecular form and function of the cytokinetic ring.J Cell Sci 132120021-953310.1242/jcs.22692831209062 PMC 6602304 · doi ↗ · pubmed ↗
- 8Moreno, S., Klar, A., Nurse, P. 1991. Molecular genetic analysis of fission yeast Schizosaccharomyces pombe. Methods Enzymol.. 194: 795-823. 532.10.1016/0076-6879(91)94059-l 2005825 · doi ↗ · pubmed ↗