The C. elegans SET1 histone methyltransferase SET-2 is not required for transgenerational memory of silencing
Cécile Bedet, Piergiuseppe Quarato, Francesca Palladino, Germano Cecere, Valérie J Robert

TL;DR
This study investigates whether the SET-2 protein is needed for RNAi-induced gene silencing in C. elegans, finding it is not required for transgenerational silencing memory.
Contribution
The study clarifies that RNAi pathways are not responsible for transcriptional misregulation in the absence of SET-2 in C. elegans.
Findings
SET-2 is not required for RNAi-induced transgenerational silencing in C. elegans.
RNAi pathway defects are not responsible for transcriptional misregulation in SET-2 absence.
Endogenous small RNA pools remain unaffected without SET-2.
Abstract
The SET-2 /SET1 histone H3K4 methyltransferase and RNAi pathway components are required to maintain fertility across generations in C. elegans . SET-2 preserves the germline transcriptional program transgenerationally, and RNAi pathways rely on small RNAs to establish and maintain transgenerational gene silencing. We investigated whether the functionality of RNAi-induced transgenerational silencing and the composition of pools of endogenous small RNA are affected by the absence of SET-2 . Our results suggest that defects in RNAi pathways are not responsible for the transcriptional misregulation observed in the absence of SET-2 .
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
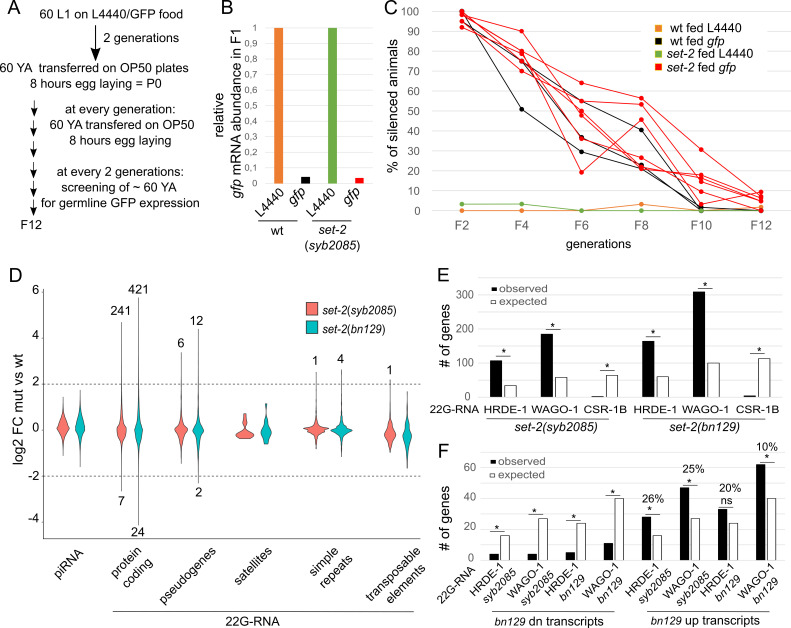 Figure 1
Figure 1|
strain |
genotype |
from |
|
|
wildtype |
sunybiotech |
|
|
|
CGC |
|
|
|
|
|
|
|
This study |
|
|
|
This study |
|
|
|
|
|
plasmid |
information |
from |
|
GFP feeding clone |
Full-length GFP cDNA cloned between 2 T7 promoters to produce GFP dsRNA. |
Gift from Marie-Anne Félix. |
|
L4440 feeding clone |
Empty vector |
|
- —Agence Nationale de la Recherche (France)https://ror.org/00rbzpz17
- —French National Centre for Scientific Research (France)https://ror.org/02feahw73
- —French National Centre for Scientific Research (France)https://ror.org/02feahw73
- —Institut Pasteur (France)https://ror.org/0495fxg12
- —Horizon 2020
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms · Epigenetics and DNA Methylation · Sirtuins and Resveratrol in Medicine
Description
In C. elegans , transgenerational loss of fertility, also referred to as the mortal germline (Mrt) phenotype, is observed in both wild isolates and mutant strains grown in laboratory conditions (Ahmed and Hodgkin 2000; Smelick and Ahmed 2005; Frézal et al. 2018; 2023). Mutants with mortal germlines fall into two classes: those that become sterile at all temperatures, and those that display a reversible temperature-sensitive Mrt (tsMrt) phenotype at the non-permissive temperature (25°C). Mutants in the first class identified genes involved in telomere replication, genome stability and histone methylation (Ahmed and Hodgkin 2000; Meier et al. 2006, 2009; Andersen and Horvitz 2007; Katz et al. 2009) . Mutants in the second class define RNAi pathway components and chromatin associated proteins including SET-2 , the C. elegans homolog of the SET1 H3K4 histone methyltransferase that plays context dependent roles in transcription (Simonet et al. 2007; Li and Kelly 2011; Xiao et al. 2011; Buckley et al. 2012; Sakaguchi et al. 2014; Spracklin et al. 2017; Howe et al. 2017; Weiser et al. 2017; Saltzman et al. 2018; Manage et al. 2020; Wan et al. 2021; Caron et al. 2021; Seroussi et al. 2023) .
RNAi pathways are key players in the regulation of gene expression both at the transcriptional and post-transcriptional levels (Billi et al. 2014; Almeida et al. 2019; Seroussi et al. 2022) . They rely on primary RNA signals that trigger RdRP-dependent production of secondary 22G-RNAs. 22G-RNAs are amplified through a self-sustaining loop, and can be transmitted to the progeny for several generations without the primary trigger, thereby playing an essential role in the transgenerational inheritance of gene expression regulation (Gu et al. 2012) . It has been proposed that alteration in the composition of 22G-RNA pools can result in the deregulation of transcriptional programs over generations, ultimately leading to loss of sterility (Buckley et al. 2012) . Supporting this model, it was shown that the absence of piRNAs (one type of primary RNA signal triggering RNAi pathways) affects the pools of 22G-RNA targeting histone mRNAs over generations. As a result, histone mRNAs are silenced and animals become sterile (Barucci et al. 2020) .
We and others have previously reported that the tsMrt phenotype observed in the absence of SET-2 correlates with loss of germline identity and transgenerational deregulation of the germline transcriptional program (Xiao et al. 2011; Robert et al. 2014, 2020; Caron et al. 2021) . In this study, we asked if transgenerational sterility and progressive deregulation of transcription programs observed in the absence of SET-2 result from a functional defect in RNAi pathways. In wild-type animals, single-copy transgenes expressing a Green Fluorescent Protein (GFP) under the control of germline promoters can be silenced when animals are fed dsRNA molecules targeting GFP , and this silencing is transmitted over 9 to 12 generations after elimination of the dsRNA trigger (Buckley et al. 2012) . To test whether this silencing process is still functional in the absence of SET-2 , we fed * set-2 ( syb2085 ) * animals carrying a catalytically dead allele of * set-2 * and the * oxIs279 * [ pie-1p :: gfp ::H2B + * unc-119 * (+)] transgene expressed in the germline with a bacteria clone expressing gfp dsRNA ( Figure 1A ) (Frøkjær-Jensen et al. 2008; Caron et al. 2021). We then monitored GFP silencing and re-expression across generations after the elimination of the dsRNA trigger. Both wild-type and * set-2 * ( * syb2085 ) * animals show a substantial decrease in mRNA gfp, which correlates with efficient silencing of the * oxIs279 * transgene observed by fluorescent imaging ( Figure 1B and 1C). Following removal of dsRNA, 100% of the wild-type animals re-expressed GFP at generation F10 in 3 independent lines, while 9-30% of the * set-2 * ( * syb2085 * ) animals at the same generation still showed GFP silencing in 5 out of 6 independent lines. By the F12 generation, 4-9% of the * set-2 * ( * syb2085 * ) animals were still silenced in 5 out of 6 independent lines. Statistical analysis (chi-square test, see Methods section) performed on pooled data at F10 and F12 shows that the small difference observed between wild-type and mutant animals is significant, suggesting that RNAi silencing memory induced at the * oxIs279 * transgene may be slightly longer in the absence of SET-2 .
Next, we asked whether * set-2 * mutant display misregulation of small RNA pools that could correlate with altered germline transcriptional programs and result in transgenerational loss of fertility. We sequenced the small RNA pools present in * set-2 * ( * syb2085 ) * animals *, * also including * set-2 * ( * bn129 ) * animals carrying a null allele of * set-2 * in our analysis. While similar pools of piRNA were found in * set-2 * mutants and wild-type animals, we identified 249 and 437 DNA elements (including protein coding genes, pseudogenes and repetitive elements) with significantly more 22G-RNAs (mean log2 foldchange ( * set-2 * vs WT) > 2) in * syb2085 * and * bn129 * mutants respectively than in wild-type. A few DNA elements with significantly less 22G-RNAs in * syb2085 * and * bn129 * mutants than in wild-type ( Figure 1D ) were also found. We further analyzed 241 and 421 genes with more 22G-RNAs in * syb2085 * and * bn129 * mutants, respectively, and that target protein coding genes. We found that, in wild-type animals, this set of genes is enriched for targets of HRDE-1- and WAGO-1-associated 22G-RNAs, and depleted for targets of CSR-1-associated 22G-RNAs ( Figure 1E ). In wild-type germlines, 22G-RNA associated with HRDE-1 and WAGO-1 are involved in germline gene silencing (Gu et al. 2009; Buckley et al. 2012) . By contrast, in * set-2 * mutant germlines we observed that instead of correlating with downregulation of target genes, overproduction of this subset of HRDE-1- or WAGO-1 associated 22G-RNA correlates with upregulation of target genes with 12% to 26% of the genes enriched for HRDE-1- or WAGO-1-associated-22G-RNA in * set-2 * mutants being upregulated in * set-2 * ( * bn129 * ) mutant germlines ( Figure 1F ). We speculate that the overproduction of 22G-RNAs in this context may result from increased transcription resulting in greater availability of mRNA template. At this stage, it is not possible to conclude whether overproduced 22G-RNA have any regulatory function on gene expression. However, their presence might disturb the balance between small RNA pathways and account for the slightly longer transgenerational memory that we observed in the absence of SET-2 .
In conclusion, SET-2 does not significantly contribute to the mechanisms underlying small RNA pathways, and small RNA pathway deregulation is unlikely to be responsible for the transgenerational loss of fertility in * set-2 * mutants. This conclusion fits with our previous observation that SET-2 works in parallel with the NRDE pathway (involved in the heritability of RNAi silencing of gene expression) to support germline immortality (Robert et al. 2014) .
Methods
dsRNA induced GFP silencing and monitoring of GFP re-expression
For RNAi induction of silencing, L4440 and dsRNA GFP clones were grown overnight at 37°C in liquid LB media complemented with Ampicillin. dsRNA expression was induced by adding IPTG to the liquid culture to a final concentration of 1mM and bacteria were grown at 37°C for 2 additional hours before seeding 1 ml on standard NGM plates complemented with 1 mM IPTG. PFR733 and PFR725 strains were synchronized by sodium hypochlorite treatment and 60 L1 animals of each strain were transferred on RNAi (L4440 and dsRNA GFP) plates. Animals developed for 48 hours at 20°C and were transferred as young adults on fresh RNAi plates for a second generation of silencing induction. Their progeny (considered as P0 in our experiment) were transferred in pools of 60 animals on OP50 plates and allowed to lay eggs for 8 hours before elimination. This “transfer/egg laying” procedure was repeated every 3 days and GFP re-expression was scored every 2 generations on 50-60 young adults immobilized on agarose pad using an AZ100M Nikon macrozoom microscope. Animals were scored as “re-expressing” GFP as soon a signal could be detected. A chi-square test was perfomed at the F10 and F12 generations on pooled data (F10, WT: 1 GFP- animal and 191 GFP+ animals, * set-2 * ( * syb2085 * ) : 61 GFP- animals and 317 GFP+ animals. Chi2 = 30.3 p-value = 3.66x10 ^-8 ^ ; F12, WT = 0 GFP- and 112 GFP+, * set-2 * ( * syb2085 * ) : 8 GFP- and 119 GFP+ ; chi2 = 7.29 and p-value = 0.007).
qRT-PCR
Mix stage populations were harvested at F1 generation, washed in M9 buffer and resuspended in 300 µl of NucleoZOL (Macherey-Nagel #740404-200). RNA was extracted according to manufacturer's protocol using Nucleospin RNA Set for NucleoZOL kit (Macherey-Nagel #740406.50). RNA was eluted in 40 µL of UltraPure water and the integrity and concentration of RNA was measured with TapeStation 4200 and RNA Screen Tape (Agilent). RNAs were retrotranscribed using the Transcriptor Universal cDNA Master kit (Roche #05893151001). qPCRs were performed with Takyon SYBR 2X MasterMix (Eurogentec #UF-NSMT-B0705) on a CFX real-time detection system (CFX 96 Biorad) and gfp RNA levels were normalized to the mean of * act-1 * and * cdc-42 * genes. primers: act-1 _for: gctggacgtgatcttactgattacc / act-1 _rev: gtagcagagcttctccttgatgtc / cdc-42 _for: ctgctggacaggaagattacg / cdc-42 _rev: ctcggacattctcgaatgaag / gfp_for: ggcggtaccggtagaaaaa / gfp_rev: ttgtgcccattaacatcacc.
small RNA sequencing and analysis
For small RNA sequencing, worm sorting was performed using a COPAS Biosorter (Union Biometrica) to obtain a population of worms enriched for the young adult stage (Cornes et al. 2022) . The mutant worms were collected four generations after homozygosis to obtain enough worms for sorting. Total RNA with RIN > 9 was used to generate small RNA libraries. The library preparation and data analysis were performed as previously described (Barucci et al. 2020) .
Reagents
**: **
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahmed Shawn Hodgkin Jonathan 200011 MRT-2 checkpoint protein is required for germline immortality and telomere replication in C. elegans Nature 40367660028-083615916410.1038/3500312010646593 · doi ↗ · pubmed ↗
- 2Almeida Miguel Vasconcelos Andrade-Navarro Miguel A. Ketting René F. 2019115 Function and Evolution of Nematode RN Ai Pathways Non-Coding RNA 512311-553X 8810.3390/ncrna 501000830650636 PMC 6468775 · doi ↗ · pubmed ↗
- 3Andersen Erik C. Horvitz H. Robert 2007815 Two C. elegans histone methyltransferases repress lin-3 EGF transcription to inhibit vulval development Development 134161477-91292991299910.1242/dev.00937317634190 · doi ↗ · pubmed ↗
- 4Barucci Giorgia Cornes Eric Singh Meetali Li Blaise Ugolini Martino Samolygo Aleksei Didier Celine Dingli Florent Loew Damarys Quarato Piergiuseppe Cecere Germano 202021 Small-RNA-mediated transgenerational silencing of histone genes impairs fertility in pi RNA mutants Nature Cell Biology 2221465-739223524510.1038/s 41556-020-0462-732015436 PMC 7272227 · doi ↗ · pubmed ↗
- 5Billi Allison C. 201457 Endogenous RN Ai pathways in C. elegans Worm Book 1551-850714910.1895/wormbook.1.170.1PMC 478113324816713 · doi ↗ · pubmed ↗
- 6Buckley Bethany A. Burkhart Kirk B. Gu Sam Guoping Spracklin George Kershner Aaron Fritz Heidi Kimble Judith Fire Andrew Kennedy Scott 2012718 A nuclear Argonaute promotes multigenerational epigenetic inheritance and germline immortality Nature 48974160028-083644745110.1038/nature 1135222810588 PMC 3509936 · doi ↗ · pubmed ↗
- 7Caron Matthieu Gely Loïc Garvis Steven Adrait Annie CoutéYohann Palladino Francesca Fabrizio Paola 20211210 Loss of SET 1/COMPASS methyltransferase activity reduces lifespan and fertility in Caenorhabditis elegans Life Science Alliance 532575-1077 e 202101140 e 20210114010.26508/lsa.20210114034893559 PMC 8675910 · doi ↗ · pubmed ↗
- 8Cornes Eric Bourdon Loan Singh Meetali Mueller Florian Quarato Piergiuseppe Wernersson Erik Bienko Magda Li Blaise Cecere Germano 202211 pi RN As initiate transcriptional silencing of spermatogenic genes during C. elegans germline development Developmental Cell 5721534-5807180196.e 710.1016/j.devcel.2021.11.02534921763 PMC 8796119 · doi ↗ · pubmed ↗