Generation and characterization of temperature-sensitive alleles encoding GPI anchored proteins Psu1 and Dfg502 in Schizosaccharomyces pombe
Bita Tavafoghi, Liping Ren, Kathleen L. Gould, Alaina H. Willet

TL;DR
This paper studies yeast proteins involved in cell wall formation and finds their roles in cell separation and viability.
Contribution
The study introduces new temperature-sensitive alleles for analyzing GPI-anchored proteins in S. pombe.
Findings
Psu1 is essential for cell separation, proper cell shape, and viability.
Dfg501 and Dfg502 have redundant and essential roles in cell separation and viability.
Abstract
Glycosyl-phosphatidylinositol (GPI) anchored proteins are implicated in remodeling of the yeast cell wall during growth and division. Schizosaccharomyces pombe proteins, Psu1 , Dfg501 , and Dfg502 are predicted GPI anchored proteins with likely cell wall modifying activity. Here, we isolated and characterized null and temperature-sensitive alleles that will allow further analysis of the function of these proteins and S. pombe cell wall formation. Our data confirm that Psu1 is necessary for cell separation, maintaining proper cell shape, and viability. Additionally, we found that Dfg501 and Dfg502 share a redundant and essential function necessary for cell separation and viability.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
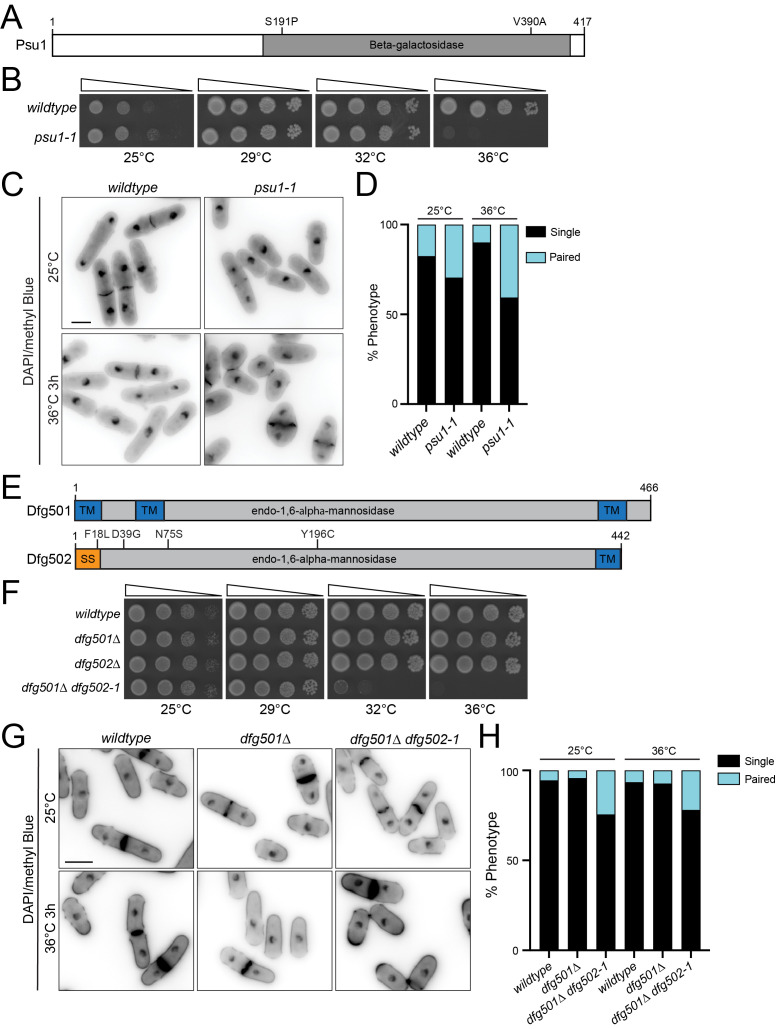 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Metabolic Engineering and Bioproduction · Fungal and yeast genetics research · Microbial Natural Products and Biosynthesis
Description
Cell-walled organisms build and remodel their cell wall during growth and division. The Schizosaccharomyces pombe cell wall is mainly composed of linear and branched glucan chains and galactomannan (Bush et al., 1974; Humbel et al., 2001; Osumi et al., 1998) . Additionally, the cell wall contains a cohort of proteins linked directly to it (de Groot et al., 2007) . Many of these are modified with a glycosyl-phosphatidylinositol (GPI) anchor and established to play important roles in the structure and function of the yeast cell wall (De Groot et al., 2003; Pittet & Conzelmann, 2007) . To become GPI-anchored, proteins are modified on a ω site near their C-terminus by lipid addition in the endoplasmic reticulum (Frieman & Cormack, 2004) . The lipid anchor can either be cleaved at the plasma membrane or remodeled (Fankhauser et al., 1993; Martin-Yken et al., 2001) . The specific way that GPI-anchored proteins are attached to the cell surface might be critical for maintaining cell wall homeostasis and allow cells to adapt to different environmental stresses. The function of GPI modified proteins is not fully understood. Here, we isolated and characterized a temperature-sensitive allele of * psu1 * . We also generated null alleles of the adjacent and presumably duplicated * dfg501 * and * dfg502 * genes and a temperature-sensitive * dfg502 * allele in the dfg501∆ strain.
Psu1 contains a beta glucosidase domain belonging to the glycosidase hydrolase 132 (GH132) enzyme family (Drula et al., 2022; Gastebois et al., 2013) . Other fungal members of this enzyme class hydrolyze linear β-(1,3)-glucans (Gastebois et al., 2013) and are implicated in promoting cell separation and septum integrity (Mouassite et al., 2000; Ritch et al., 2010) . S. pombe * psu1 * is essential and when deleted the cells display cell separation defects, become rounded, and eventually lyse and die (Omi et al., 1999) .
To further study Psu1 function, we isolated a new * psu1 * temperature-sensitive allele using an error-prone PCR method (Tang et al., 2019) . Sequencing revealed substitutions of serine 191 to proline and valine 390 to alanine within the catalytic domain ( Figure 1A ). Serial dilutions of wildtype and psu1-1 cells spotted at a variety of temperatures showed that both strains grew similarly at 25°C, 29°C and 32°C, but psu1-1 did not grow at 36°C while wildtype grew robustly ( Figure 1B ). We next grew cells at 25°C, shifted some of them to 36°C for 3 hours, and then fixed the cells from each condition. The cells were stained with DAPI and methyl blue (MB) to mark DNA and the cell wall, respectively, and imaged. We found that at 25°C psu1-1 cells had an increase in paired cells compared to wildtype and this difference was exacerbated at 36°C, indicating a cell separation defect ( Figure 1C ). Additionally, consistent with the psu1∆ phenotype, psu1-1 cells were rounded at high temperature ( Figure 1C ) (Omi et al., 1999) . Thus, the temperature-sensitive and null mutant phenotypes are concordant, and our data validates the importance of Psu1 for maintaining proper cell shape and promoting cell separation following cytokinesis (Omi et al., 1999) .
dfg501 * and * dfg502 * encode putative endo-1,6-α-mannosidases belonging to the glycoside hydrolase 76 (GH76) enzyme family (Drula et al., 2022) and are orthologous to *Saccharomyces cerevisiae * Dfg5 (Kitagaki et al., 2002) . Endo-1,6-α-mannosidases hydrolyze unbranched 1,6-α-mannose and these enzymes are hypothesized to have a role in transferring GPI anchored proteins from the plasma membrane to the cell wall, however their precise function has not been confirmed biochemically (Kitagaki et al., 2002) .
To study * dfg501 * and * dfg502 * function we aimed to make single and double gene deletions of these adjacent genes. While we were able to construct dfg501∆ and *dfg502∆ * single deletion strains, we were unable to recover a double deletion strain. We therefore generated a * dfg502 * temperature-sensitive allele in a dfg501∆ genetic background. The temperature-sensitive allele contained F18L, D39G, N75S and Y196C substitutions within Dfg502 ( Figure 1D ). Analysis of cell growth revealed that each single deletion grew similarly to wildtype at all temperatures tested but dfg501∆ dfg502-1 had reduced growth at 32°C compared to wildtype and did not grow at 36°C ( Figure 1E ). Fixing and staining of the cells with DAPI and MB revealed that dfg501∆ dfg502-1 cells had an increase in the frequency of paired cells at both 25°C and 36°C compared to wildtype cells, indicative of a cell separation defect ( Figure 1F ). These results indicate that Dfg501 and Dfg502 are important for promoting cell separation, which may be a common theme among GPI anchored enzymes. Interestingly, gas1-1 , psu1-1 and dfg501∆ dfg502-1 all have cell separation defects (Howard et al., 2024) . Dfg5 and Gas1 defective cells also have similar phenotypes noted in S. cerevisiae , suggesting they may play similar roles in remodeling the cell walls of these two yeast species (Kitagaki et al., 2002) .
In conclusion, GPI anchored proteins are physically poised to enact cell wall modification to allow for cell growth and division and it will be interesting to learn how they collaborate in space and time to allow precise cell wall remodeling in response to cell growth and environmental changes.
Methods
Yeast methods
S. pombe strains were grown in yeast extract (YE) and standard S. pombe mating, sporulation, and tetrad dissection techniques were used to construct new strains (Moreno et al., 1991) .
Molecular biology methods
The * psu1 * allele was sequenced by generating a PCR product with an oligonucleotide 85 bp upstream of the start site (CTTCGTTCGTTCCTTGAATTTTAGACACA) and a reverse oligonucleotide within kanMX6 (Integrated DNA technologies). The PCR product was sequenced with a forward oligonucleotide 580 bp into the open reading frame (GGTGATGGTGCTGGTTCCTCTTGCGTTG) and a reverse oligonucleotide 679 bp into the open reading frame (GCAAACCACCACGGGTTTCACCGTCAG).
The dfg502-1 allele was sequences by generating a PCR product with an oligonucleotide 300 bp upstream of the start site (GCTCGCATTGAAATTTATTTGGTTAC) and a reverse oligonucleotide within kanMX6 (Integrated DNA technologies). The PCR product was sequenced with a forward oligonucleotide 144 bp into the open reading frame (CCATAAATAGTGCCTTGACTACCGTCACTGACGG) and a reverse oligonucleotide 860 bp into the open reading frame (CCTTCCATATGGGTTTGCCACACAGAACTGCC). The * dfg501 * and * dfg502 * gene deletions were made as previously described (Chen et al., 2015) .
Isolation of temperature sensitive alleles with error-prone PCR
Temperature-sensitive mutants of * psu1 * and * dfg502 * were constructed and isolated based on the previously described protocol (Tang et al., 2019) but using EX taq polymerase (Takara, 4025) and accompanying dNTPs (Takara, RR01BM). The * dfg502 * temperature sensitive allele was made in the * dfg501 ::kanMX6 * genetic background and hphMX6 selection linked to * dfg502 * .
Microscopy and image analysis
Strains for fixed-cell imaging experiments were grown at 25°C in YE and then shifted to 36°C for 3 hours. Cells were fixed with 70% ethanol for DAPI and methyl blue (MB) staining as described previously (Roberts-Galbraith et al., 2009) . Images were acquired using a Zeiss Axio Observer inverted epifluorescence microscope with Zeiss 63× oil (1.46 NA) and captured using Zeiss ZEN 3.0 (Blue edition) software. A singular medial Z slice was obtained. All images were further processed using ImageJ (Schindelin et al., 2012) . Graphs were constructed with Prism 8.0 (GraphPad Software).
Reagents
The strains used in this study and their genotypes are listed below.
Strain Genotype Source
KGY246 * ade6-M210 leu1-32 ura4-D18 h ^-^ * Lab stock
KGY5216-2 *psu1-1(S191, V390A):kanMX6 ade6-M210 leu1-32 * This study
ura4-D18 h ^-^ *
KGY3698-2 dfg501Δ::kanMX6 ade6-M210 leu1-32 ura4-D18 * h ^-^ * This study
KGY3699-2 dfg502Δ::kanMX6 * ade6-M210 leu1-32 ura4-D18 h ^-^ * This study
KGY5820-2 dfg501Δ::kanMX6 ade6-M210 leu1-32 ura4-D18 * h ^-^ * This study
dfg502-1(F18L, D39G, N75S, Y196C):hphMX6
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bush D. A. Horisberger M. Horman I. Wursch P. 200011 The Wall Structure of Schizosaccharomyces pombe Microbiology 8111350-087219920610.1099/00221287-81-1-1994822119 · doi ↗ · pubmed ↗
- 2Chen Jun-Song Beckley Janel R Mc Donald Nathan A Ren Liping Mangione Maria Santa Jang Sylvia J Elmore Zachary C Rachfall Nicole Feoktistova Anna Jones Christine M Willet Alaina H Guillen Rodrigo Bitton Danny A Bähler Jürg Jensen Michael A Rhind Nick Gould Kathleen L 201531 Identification of New Players in Cell Division, DNA Damage Response, and Morphogenesis Through Construction of Schizosaccharomyces pombe Deletion Strains G 3 Genes|Genomes|Genetics 532160-183636137010.1534/g 3.114.015701 PMC 434909025552606 · doi ↗ · pubmed ↗
- 3de Groot Piet W. J. Yin Qing Yuan Weig Michael Sosinska Grazyna J. Klis Frans M. de Koster Chris G. 2007118 Mass spectrometric identification of covalently bound cell wall proteins from the fission yeast Schizosaccharomyces pombe Yeast 2440749-503X 26727810.1002/yea.144317230583 · doi ↗ · pubmed ↗
- 4de Groot Piet W. J. Hellingwerf Klaas J. Klis Frans M. 200369 Genome‐wide identification of fungal GPI proteins Yeast 2090749-503X 78179610.1002/yea.100712845604 · doi ↗ · pubmed ↗
- 5Drula Elodie Garron Marie-Line Dogan Suzan Lombard Vincent Henrissat Bernard Terrapon Nicolas 20211129 The carbohydrate-active enzyme database: functions and literature Nucleic Acids Research 50D 10305-1048 D 571D 57710.1093/nar/gkab 1045 PMC 872819434850161 · doi ↗ · pubmed ↗
- 6Fankhauser, C, Homans, S W, Thomas-Oates, J E, Mc Conville, M J, Desponds, C, Conzelmann, A, Ferguson, M A 1993. Structures of glycosylphosphatidylinositol membrane anchors from Saccharomyces cerevisiae. The Journal of biological chemistry. 268: 26365.8253761 · pubmed ↗
- 7Frieman M. B. 2004101 Multiple sequence signals determine the distribution of glycosylphosphatidylinositol proteins between the plasma membrane and cell wall in Saccharomyces cerevisiae Microbiology 150101350-08723105311410.1099/mic.0.27420-015470092 · doi ↗ · pubmed ↗
- 8Gastebois Amandine Aimanianda Vishukumar Bachellier-Bassi Sophie Nesseir Audrey Firon Arnaud Beauvais Anne Schmitt Christine England Patrick Beau Rémi Prévost Marie-Christine d'Enfert Christophe LatgéJean-Paul Mouyna Isabelle 201351 SUN Proteins Belong to a Novel Family of β-(1,3)-Glucan-modifying Enzymes Involved in Fungal Morphogenesis Journal of Biological Chemistry 288190021-9258133871339610.1074/jbc.m 112.44017223508952 PMC 3650377 · doi ↗ · pubmed ↗