Comparison of tet(X4)-containing contigs assembled from metagenomic sequencing data with plasmid sequences of isolates from a cohort of healthy subjects
Yichen Ding, Shuan Er, Abel Tan, Jean-Sebastien Gounot, Woei-Yuh Saw, Linda Wei Lin Tan, Yik Ying Teo, Niranjan Nagarajan, Henning Seedorf

TL;DR
This study compares metagenomic sequencing with cultivation methods to detect the antibiotic resistance gene tet(X4) in healthy individuals' gut microbiota.
Contribution
The study demonstrates the potential and limitations of metagenomic sequencing for detecting antibiotic resistance genes in clinical samples.
Findings
Metagenomic sequencing detected tet(X4) in 2 out of 109 subjects, while cultivation found it in 10.1%.
tet(X4)-containing sequences from metagenomic data showed high similarity to plasmid sequences from isolates.
Cultivation-based methods remain more sensitive for low-abundance antibiotic resistance gene detection.
Abstract
Recently discovered tet(X) gene variants have provided new insights into microbial antibiotic resistance mechanisms and their potential consequences for public health. This study focused on detection, analysis, and characterization of Tet(X4)-positive Enterobacterales from the gut microbiota of a healthy cohort of individuals in Singapore using cultivation-dependent and cultivation-independent approaches. Twelve Tet(X4)-positive Enterobacterales strains that were previously obtained from the cohort were fully genome-sequenced and comparatively analyzed. A metagenomic sequencing (MS) data set of the same samples was mined for contigs that harbored the tet(X4) resistance gene. The sequences of tet(X4)-containing contigs and plasmids sequences were compared. The presence of the resistance genes floR and estT (previously annotated as catD) was detected in the same cassette in 10 and 12 out…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
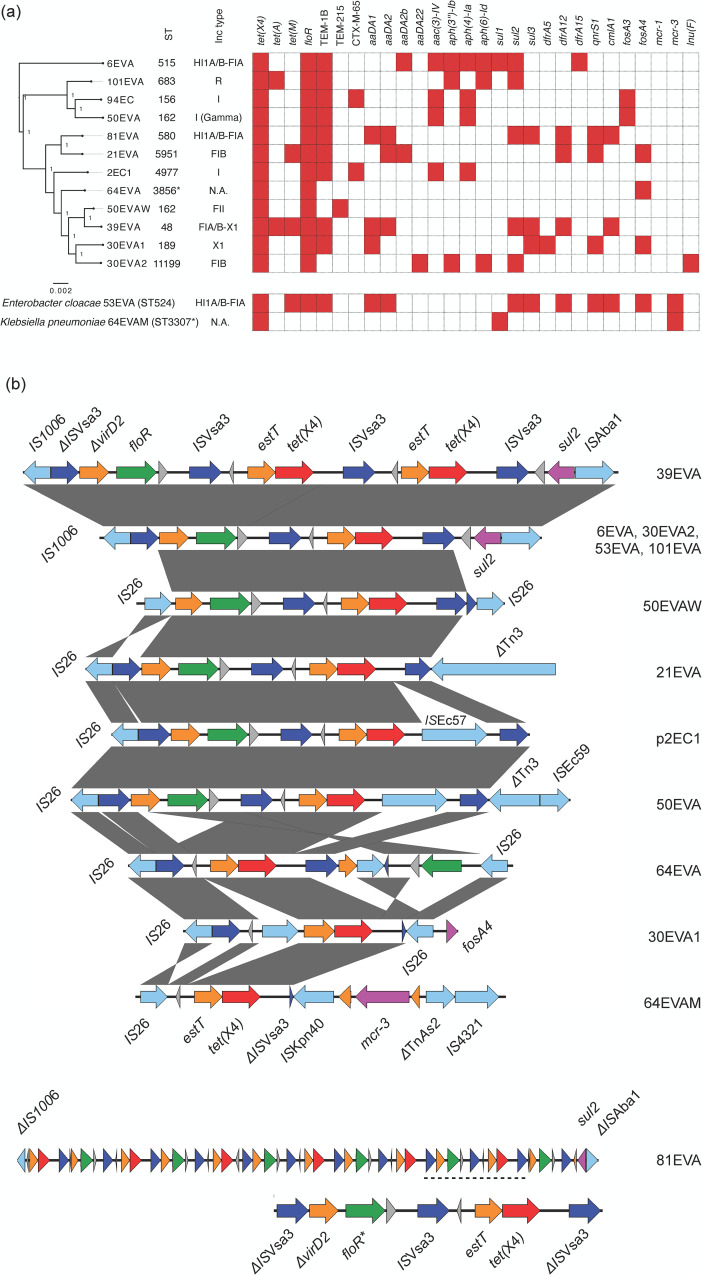 Fig 1
Fig 1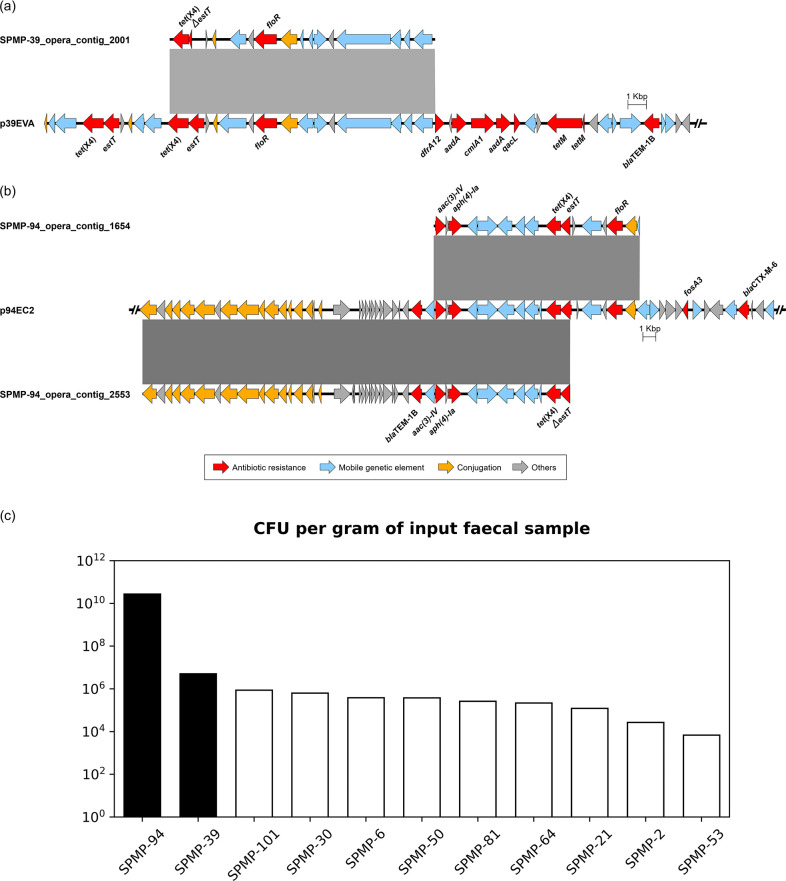 Fig 2
Fig 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Pharmaceutical and Antibiotic Environmental Impacts · Antibiotic Use and Resistance
INTRODUCTION
Genomic surveillance is one important measure to monitor and control the spread of multidrug resistant bacteria (MDR) in the human population. In general, the prevalence for MDR bacteria is determined through a cultivation-dependent approach; i.e., clinical or environmental samples are inoculated into selective agar plates supplemented with antibiotics to obtain MDR bacterial colonies, which will be confirmed with phenotypic and molecular assays. Selected colonies can be further subjected to whole-genome sequencing and comparative genomic analysis to determine their antibiotic resistance genes, plasmid sequences, and possible transmission clusters. This approach has allowed detailed insights into the genomic structure of many MDR bacteria by numerous studies, especially for the recently emerged tet(X) family-mediated tigecycline-resistant Enterobacterales (1, 2). On the other hand, shotgun metagenomic sequencing (MS) characterizes the microbial communities in clinical and environmental samples via an unbiased culture-independent approach in which the total DNA of the samples is extracted and sequenced. Subsequently, the antibiotic resistance genes of interest can be further analyzed using the contigs assembled from metagenomic sequencing data to determine their presence in the samples, as well as the associated plasmid types and host species. Shotgun metagenomic sequencing may therefore complement culture-based whole-genome sequencing approaches for rapid identification of MDR bacteria, especially when cultivation of bacteria is difficult, or a high-throughput screen is required.
The recently identified tet(X) family tigecycline resistance genes are variants of the initially described tet(X) (3). These new variants share 95% sequence identity with tet(X), which also confers high-level resistance to last-resort antibiotics such as tigecycline (4), posing a serious threat to public health (5). Among the various emerging tet(X) variants, tet(X4) has been identified in animals, healthy individuals, and patients in multiple provinces of China and other regions (6), and its successful transmission could be attributed to conjugative plasmids and ISCR2-mediated transposition (7). We previously reported that the prevalence of Tet(X4)-producing Enterobacterales in the gut microbiota of healthy individuals in Singapore is 10.1% and analyzed the sequences of two IncI1-type plasmids (p2EC1-1 and p94EC-2) that carry tet(X4) (1). Here, we further sequenced and characterized additional 12 tigecycline-resistant Enterobacterales strains isolated from human fecal samples in Singapore. We show that tet(X4) is associated with a diverse range of plasmid types and hosts and is potentially co-transferred with florfenicol resistance gene floR and tylosin resistance gene estT. The latter has only recently been characterized as a serine-dependent macrolide esterase (8, 9). We further leveraged on recently published high-quality metagenomic sequence data for the same fecal samples collected from the cohort to evaluate if contigs assembled from metagenomic sequencing data could reveal tet(X4) plasmid sequences (1, 10). Our findings suggest that metagenomic sequencing could complement culture-based surveillance for MDR bacteria if they are present at high abundance in clinical samples.
MATERIALS AND METHODS
Sample collection and DNA extraction
The collection and DNA extraction of fecal samples have been described previously (1, 10). In brief, feces from 109 individuals aged 48–76 years old of the Singapore Integrative Omics Study were collected in 2018 using a BioCollector (BioCollective) kit, according to the manufacturer’s instructions. Fecal samples were handled in a Coy anaerobic chamber containing N_2_ (75%), CO_2_ (20%), and H_2_ (5%) gas mixture. Homogenized samples were transferred to 50-mL screw-cap tubes prior to storage at –80°C. The QIAamp Power Fecal Pro DNA kit was used to extract gDNA for genomic (2 × 2 mL pure culture, OD_600_ = 0.17) and metagenomic (fecal material, ~0.5 g) sequencing. DNA for genomic sequencing was further purified using a Qiagen Genomic-tip 20/G kit as described in the manufacturer’s protocol (Qiagen, Germany). Cells from cultures were concentrated at 10,000 × g for 15 min before DNA extraction. DNA was quantified using a Qubit v.1.0 fluorometer with a broad-range assay kit (Life Technologies) and a NanoDrop-2000 (Thermo Fisher Scientific).
Genome sequencing and data analysis
Genomic DNA of previously isolated strains was extracted using Qiagen Genomic-tip 20/G as per manufacturer’s instructions. Whole-genome sequencing was performed using MinION and Illumina Novaseq, followed by genome assembly and polishing using Flye v.2.9 (11, 12) and Pilon v.1.24 (13), respectively. The assembled complete genomes were subjected to sequence typing by online MLST v.2.0 (14), phylogenetic analysis using the Harvest Suite (15), antibiotic resistance gene prediction by ResFinder v.4.1 (16), plasmid typing by PlasmidFinder v.2.0 (17), and identification of insertion sequences by ISFinder (18). Comparative sequence analysis was performed using EasyFig v.2.2.5 (19) running BLAST+ v.2.13.0 (20).
Metagenomic sequencing assembly and analysis
MS contigs are derived from the Singapore Platinum Metagenomes Project (SPMP) (10), which was conducted on DNA extracted from the same fecal samples that were also used for the cultivation-based analysis. Contigs containing the tet(X4) gene were identified using BLAST, and subsequent verification was performed using ResFinder with default settings (16, 20).
CFU counting
Colony-forming unit (CFU) counting experiment was done for our previous study (1). Briefly, frozen fecal samples were weighed and inoculated into Luria broth, followed by incubation at 37°C with 200-rpm shaking for 3 h. The fecal suspensions were then serially diluted in 0.9% NaCl and spotted onto MacConkey agar plates supplemented with 2-mg/L eravacycline dihydrochloride. The CFUs were enumerated after incubation at 37°C for 18 h, and the results were normalized to CFU per gram of input fecal sample.
GenBank accession numbers
MS short and long reads can be found under BioProject number PRJEB49168, and genomes sequences can be found under BioProject number PRJNA599529.
RESULTS AND DISCUSSION
Characterization of tet(X4)-carrying plasmids by whole-genome sequencing
Twelve Enterobacterales strains that are positive for tet(X4) were previously isolated from human fecal samples on MacConkey agar plates supplemented with eravacycline (1). Their genomes have been sequenced to complete-genome level by Illumina and Nanopore. In total, tet(X4) was carried by seven different plasmid types, including IncHI1A/B-IncFIA (n = 3), IncFIB (n = 2), IncI (gamma, n = 1), IncX1 (n = 1), IncFIA/B-IncI (n = 1), IncFII (n = 1), and IncR (n = 1), while two plasmids were non-typable (Fig. 1a). The host bacterial species include Escherichia coli (n = 10), Klebsiella pneumoniae (n = 1, isolate 64EVAM, ST3307*), and Enterobacter cloacae (n = 1, isolate 53EVA, ST524) (Fig. 1a). In particular, the 10 tet(X4)-positive E. coli strains belonged to 10 different sequence types (Fig. 1a). These results suggested that a broad range of E. coli strains with diverse genetic backgrounds had been associated with tet(X4) in Singapore, which is consistent with findings previously reported in other regions such as China, Thailand, and Pakistan (6, 7, 21).
(a) Antibiotic resistance gene profiling and phylogenetic analyses of tet(X4)-positive Enterobacterales strains. The phylogeny, sequence type, and incompatibility group of the tet(X4)-harboring plasmids are shown in the figure (N.A. indicates the plasmid is not typeable). The heat map shows the antibiotic resistance genes carried by tet(X4)-carrying plasmid. The presence of antibiotic resistance genes is indicated by red blocks. (b) Comparison of tet(X4) genetic environments. Open reading frames and their directions of transcription are indicated by colored arrows. Red: tet(X4); blue: ISVsa3 (corresponding to ISCR2); green: floR; magenta: other antibiotic resistance genes; light blue: mobile genetic elements and transposases; orange: genes with putative functions; gray: hypothetical proteins. Truncated genes are indicated by the symbol Δ, whereas floR contains a mutation indicated by an asterisk () for isolate 81EVA.*
We further characterized and compared the genetic environments of tet(X4) among the 10 plasmids (Fig. 1b). We also included two previously reported isolates, 2EC1 and 94EC, which carried tet(X4) and blaCTX-M-65 in our comparative genomic analysis (1). It was found that tet(X4) is closely associated with ISVsa3 by having at least one copy of ISVsa3 at its upstream, except for 64EVAM. This is consistent with previous studies showing tet(X4) is probably mobilized via ISVsa3 (ISCR2)-mediated transposition (1, 2). In addition, we found that other resistance genes are co-occurring with tet(X4); i.e., the estT is found in all 12 cassettes, while florfenicol resistance gene floR is located in the same cassette in 10 out of the 12 tet(X4)-carrying plasmids (Fig. 1b), including ΔISVsa3-ΔvirD2-floR-ISVsa3-estT-tet(X4)-ISVsa3 (39EVA, 6EVA, 30EVA2, 53EVA, 101EVA, and 81EVA); ΔISVsa3-ΔvirD2-floR-ISVsa3-estT-tet(X4)-ISEC57-ISVsa3 (50EVA); IS26-ΔvirD2-floR-ISVsa3-estT-tet(X4)-ISVsa3 (21EVA and 50EVAW); and IS26-ISVsa3-estT-tet(X4)-ISVsa3-orf-IS26-floR-IS26 (64EVA). For 30EVA1 and 64EVAM, although floR and tet(X4) are not in the same cassette, floR was either on the same plasmid as tet(X4) in 30EVA1 or carried by another plasmid in 64EVAM (Fig. 1a). Such close association was not found for other antibiotic resistance genes identified in the 12 strains (Fig. 1). The floR gene could confer resistance to florfenicol and chloramphenicol, while estT confers resistance against 16-membered ring-containing macrolide antibiotics, including tylosin, tilmicosin, and tildipirosin (22–24). Most of these antibiotics are commonly used as veterinary medicine in aquaculture, swine, cattle, and poultry (23, 25–27) . Similarly, the emergence of tet(X4) and other tet(X) variants was suggested to be related to the overuse of tetracycline in the food industry in China (2). However, Singapore lacks primary food industry and imports most of the agriculture products from other countries. The co-carriage of floR, estT, and tet(X4) by MDR plasmids isolated in healthy individuals in Singapore suggested that their origin might be linked to importation of animal products from other countries (28, 29). Nonetheless, we could not rule out that the emergence of tet(X4)-carrying MDR plasmids in Singapore could also arise due to the rampant inappropriate use of antimicrobials in various other sectors, while further studies should be carried out to track their origin.
Evaluation of shotgun metagenomic sequencing in detection of tet(X4)-carrying plasmids
In total, 11 fecal samples contain tet(X4)-positive Enterobacterales, and the tet(X4)-carrying plasmid sequences were analyzed in this study (Fig. 1) and in our previous study (1). To assess if shotgun metagenomic sequencing could detect tet(X4)-carrying plasmids, we further screened the contigs assembled from shotgun metagenomic sequencing for tet(X4). Interestingly, we found that tet(X4)-harboring contigs can only be detected in two fecal samples (subject SPMP-39 and SPMP-94). The sizes of the tet(X4)-harboring contigs (14–33 kbp) were shorter than the plasmids (101–134 kbp). A comparison of the tet(X4)-harboring contigs with the plasmid sequences revealed high homology of the contigs to the plasmid sequences (Fig. 2). This finding indicates that shotgun metagenomic sequencing may potentially aid in the detection of tet(X4) and its surrounding genetic environment.
Comparative analysis between metagenomic sequencing-assembled contigs and plasmid sequences containing tet(X4) genes derived from (a) SPMP-39 and (b) SPMP-94. Genes and their respective transcriptional directions are represented by arrows with colors indicating their functional classifications. Shown are truncated sequences of the plasmids. Truncated genes are indicated by the symbol Δ. The gray-shaded rectangles indicate regions (>4,500 bp) of (a) 98.8% and (b) >99.5% homology. (c) Colony-forming unit counts of tet(X4)-positive Enterobacterales from fecal samples. Contigs assembled from shotgun metagenomic sequencing data contained tet(X4)-carrying contigs for subjects SPMP-39 and SPMP-94 (black bars) but not for the other fecal samples (white bars). Of note, the fecal samples were incubated in lysogeny broth prior to inoculation onto selective agar plate (see Materials and Methods). This is to allow the tet(X4)-positive Enterobacterales to recover from frozen stock before being exposed to the antibiotic for accurate CFU counting. All fecal samples were incubated under the same conditions for the same period of time.
Enterobacterales is often present in low abundance in the human gut, which may potentially result in lower sensitivity for the detection of its associated antibiotic resistance genes when using shotgun metagenomic sequencing. We therefore wondered if the detection of tet(X4)-carrying contigs from shotgun metagenomic sequencing data is related to the abundance of the tet(X4)-positive Enterobacterales in the fecal samples. Interestingly, out of the three tet(X4)-harboring contigs identified, two were detected in subject SPMP-94, who uncoincidentally has a much higher CFU count—by four orders of magnitude—than subject SPMP-39 and the other nine samples for which MS failed to detect tet(X4)-containing contigs (Fig. 2c). Thus, these results suggest that shotgun metagenomic sequencing could detect tet(X4)-harboring plasmids when the bacteria containing the plasmid are present in high abundance in clinical samples.
Taken together, we report that tet(X4) is associated with a broad range of plasmids and host bacteria in the gut of healthy Singaporeans and is closely associated with florfenicol resistance gene floR and tylosin resistance gene estT. By comparing the contigs assembled from shotgun metagenomic sequencing, we show that this approach could complement culture-based detection of tet(X4) plasmids in human fecal samples when present at higher abundance. Further optimization is required if metagenomic sequencing should be used to discover MDR from clinical and environmental samples. However, selective cultivation currently remains the most reliable and cost-effective approach for detection of antibiotic-resistant bacteria.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ding Y, Saw WY, Tan LWL, Moong DKN, Nagarajan N, Teo YY, Seedorf H. 2020. Emergence of tigecycline- and eravacycline-resistant Tet(X 4)-producing Enterobacteriaceae in the gut microbiota of healthy Singaporeans. J Antimicrob Chemother 75:3480–3484. doi:10.1093/jac/dkaa 37232853333 · doi ↗ · pubmed ↗
- 2He T, Wang R, Liu D, Walsh TR, Zhang R, Lv Y, Ke Y, Ji Q, Wei R, Liu Z, et al.. 2019. Emergence of plasmid-mediated high-level tigecycline resistance genes in animals and humans. Nat Microbiol 4:1450–1456. doi:10.1038/s 41564-019-0445-231133751 · doi ↗ · pubmed ↗
- 3Guiney Jr DG, Hasegawa P, Davis CE. 1984. Expression in Escherichia coli of cryptic tetracycline resistance genes from bacteroides R plasmids. Plasmid 11:248–252. doi:10.1016/0147-619x(84)90031-36379711 · doi ↗ · pubmed ↗
- 4Moore IF, Hughes DW, Wright GD. 2005. Tigecycline is modified by the flavin-dependent monooxygenase Tet X. Biochemistry 44:11829–11835. doi:10.1021/bi 050606616128584 · doi ↗ · pubmed ↗
- 5Dong N, Zeng Y, Cai C, Sun C, Lu J, Liu C, Zhou H, Sun Q, Shu L, Wang H, Wang Y, Wang S, Wu C, Chan EW, Chen G, Shen Z, Chen S, Zhang R. 2022. Prevalence, transmission, and molecular epidemiology of tet(X)-positive bacteria among humans, animals, and environmental niches in China: an epidemiological, and genomic-based study. Sci Total Environ 818:151767. doi:10.1016/j.scitotenv.2021.15176734801490 · doi ↗ · pubmed ↗
- 6Mohsin M, Hassan B, Martins WM, Li R, Abdullah S, Sands K, Walsh TR. 2021. Emergence of plasmid-mediated tigecycline resistance tet(X 4) gene in Escherichia coli isolated from poultry, food and the environment in South Asia. Sci Total Environ 787:147613. doi:10.1016/j.scitotenv.2021.14761333992939 · doi ↗ · pubmed ↗
- 7Sun C, Cui M, Zhang S, Liu D, Fu B, Li Z, Bai R, Wang Y, Wang H, Song L, Zhang C, Zhao Q, Shen J, Xu S, Wu C, Wang Y. 2020. Genomic epidemiology of animal-derived tigecycline-resistant Escherichia coli across China reveals recent endemic plasmid-encoded tet(X 4) gene. Commun Biol 3:412. doi:10.1038/s 42003-020-01148-032737421 PMC 7395754 · doi ↗ · pubmed ↗
- 8Dhindwal P, Myziuk I, Ruzzini A. 2023. Macrolide esterases: current threats and opportunities. Trends Microbiol 31:1199–1201. doi:10.1016/j.tim.2023.08.00837689489 · doi ↗ · pubmed ↗