The P granule antibody KT3 recognizes epitopes in both PGL-1 and PGL-3
Brennan M Danlasky, Mario Martinez, Madeline Cassani, Devavrat Bodas, Andrea Putnam, Geraldine Seydoux

TL;DR
This paper shows that the KT3 antibody binds to two similar proteins, PGL-1 and PGL-3, likely due to a shared region.
Contribution
The study identifies that KT3 recognizes both PGL-1 and PGL-3 through a shared epitope in their disordered region.
Findings
KT3 antibody recognizes both PGL-1 and PGL-3 proteins.
The recognition is likely due to a shared epitope in the intrinsically disordered region of these proteins.
Abstract
The KT3 antibody is a commercially available antibody that recognizes the P granule protein PGL-3 (Takeda et al., 2008). Using immunostaining and western blotting of purified peptide fragments, we show that KT3 recognizes both PGL-3 and its paralog PGL-1 , likely through a shared epitope in the intrinsically disordered region.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
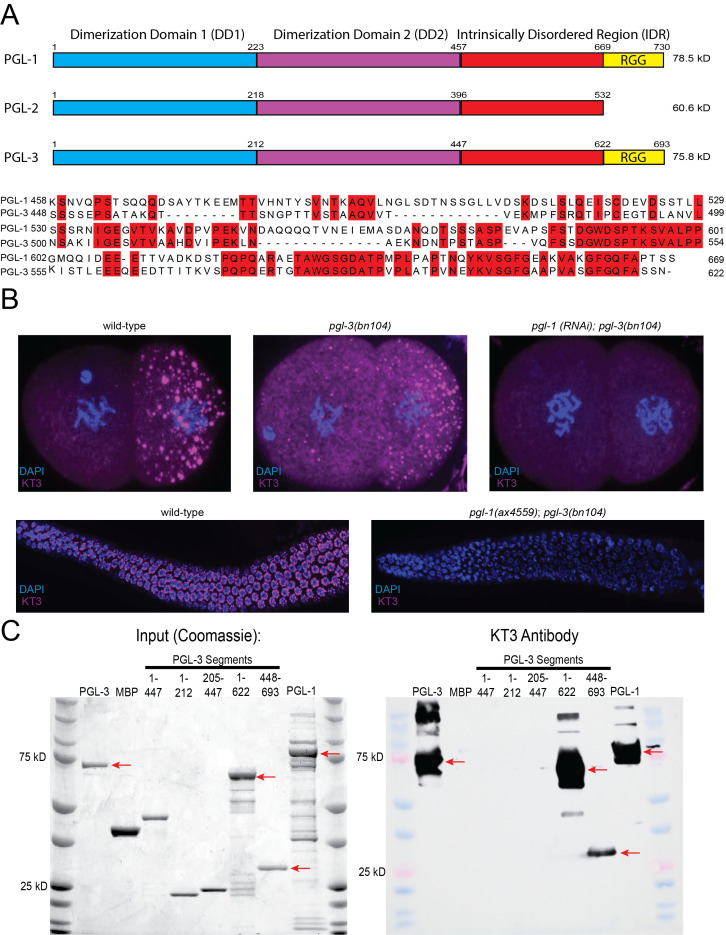 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGlycosylation and Glycoproteins Research · RNA Research and Splicing · Cancer-related gene regulation
Description
The KT3 antibody and other antibodies obtained via antigen subtraction are widely used reagents for studying *C. elegans * (Takeda et al. 2008, Hanazawa et al. 2011, Schmidt et al., 2021) *. * Antigen subtraction is a two round hybridoma production and screening method designed to raise monoclonal antibodies against random, relatively low abundance proteins present in a tissue lysate. The KT3 antibody was originally characterized (Takeda et al., 2008) as an antibody against PGL-3 , a 75kD P granule protein (Kawasaki et al., 2004) . In immunostaining experiments in embryos, KT3 recognizes P granules and an unidentified ( * pgl-3 * independent) epitope in muscle (Takeda et al., 2008) .
*C. elegans * has three PGL paralogs, each with two conserved N-terminal dimerization domains and a less conserved intrinsically disordered region (IDR) ( Fig. 1A, Kawazaki et al ., 2004). The PGL-1 and PGL-3 IDRs share some homology with each other, but not with the PGL-2 IDR. In immunostaining of embryos, we noticed that KT3 recognizes P granules in both wild-type and * pgl-3 ( bn104 ) * knockout embryos , suggesting that KT3 might recognize another P granule epitope besides PGL-3 ( Fig. 1B ). Consistent with this view, no P granule staining was observed in * pgl-3 ( bn104 ); pgl-1 (RNAi) * embryos and adult * pgl-3 ( bn104 ); pgl-1 ( ax4559 ) * germlines, suggesting that KT3 also recognizes PGL-1 , but not PGL-2 .
To further define KT3 specificity, we tested KT3 against recombinant PGL-1 , PGL-3 , and PGL-3 segments partially purified from *E. coli * ( Fig. 1C ). The KT3 antibody recognized full length PGL-1 and PGL-3 , as well as the PGL-3 IDR and a truncated PGL-3 lacking the RGG domain. KT3 did not recognize PGL-3 segments lacking the IDR.
We conclude that KT3 recognizes P granule proteins PGL-1 and PGL-3 , likely through a shared epitope in the IDR.
Methods
** CRISPR generation of PGL-1 deletion **
JH4230 * pgl-1 ( ax4559 ); pgl-3 ( bn104 ) * was generated via CRISPR/CAS9 genome editing as described in Paix et al., 2017.
Protein purification
MBP::6XHis::TEV:: PGL-3 was expressed in Rosetta (DE3) cells in terrific broth + ampicillin (100 μg/mL) to an OD600 of ~1.0 at 37°C and induced with 1 mM isopropyl β-D-1-thiogalactopyranoside at 16° C for 16 hr. Cells were resuspended in Buffer A (25 mM HEPES pH 7.4, 500 mM NaCl, 0.4 M L-Arginine, 20% (vol/vol) glycerol, 1 mM DTT) with added protease inhibitors, 5 μg/mL RNase A (USB), 0.25 U/μL RNase I (Ambion), lysed by sonication, spun at 13,000 rpm for 15 min. Lysates were filtered and incubated with amylose resin (New England Biolabs). The resin was washed 3X with Buffer B (25 mM HEPES pH 7.4, 20% (vol/vol) glycerol, 1 mM DTT, 500 mM NaCl). Protein was eluted in Buffer B + 20 mM Maltose. 6XHis::TEV::5XArg tag protease (expression plasmid obtained from addgene (PRK793) and purified as described (Tropea, Cherry, and Waugh 2009) was added at a ratio of ~ 1:5 mg TEV:PGL and incubated O/N at 16° C. After TEV cleavage MBP::6XHis and 6XHis::TEV::5XArg protease were removed by passing the cleavage reaction through a 5 mL HisTRAP column (GE Healthcare). The flowthrough containing cleaved, untagged PGL-3 was diluted to ~150 mM NaCl with Buffer B0 (containing 0 mM NaCl) and immediately loaded onto a HeparinTrap column (GE Healthcare). Proteins were eluted with a gradient of 150 to 1000 mM NaCl. PGL-3 mutants lacking the c-terminal RGG domain did not bind to the heparin column and were collected from the flowthrough and applied to a 5 mL HiTrap Q column (GE Healthcare). Proteins were eluted with a gradient of 150 to 1000 mM NaCl. Protein containing fractions were concentrated and flash-frozen in small aliquots in liquid nitrogen and stored at −80°C. Protocol adapted from Folkmann et al. 2021 and Putnam et al., 2019.
RNAi
L4 larvae were placed on IPTG-induced lawns of HT115 bacteria bearing L4440-based plasmid for 24 hours at 25°C followed by fixation, and then shifted to 20°C for one hour to avoid temperature dependent changes on P granules. The RNAi clone for * pgl-1 * came from the genomic RNAi feeding library (Medical Research Council Gene Services, Source BioScience, Nottingham, UK; Kamath et al., 2003).
Immunostaining
For embryo immunostaining, adult worms were placed into M9 on poly-l-lysine (0.01%)-coated slides and pressed under a coverslip to extrude embryos. For adult germline immunostaining, adult worms were placed in M9 buffer with 10 mM levamisole before worms were sliced open to extrude germlines. For both embryos and adult germlines, slides were laid on aluminum blocks pre-chilled by dry ice for more than 5 min. Coverslips were promptly removed to freeze-crack and permeabilize embryos and germlines. Both were then incubated overnight in methanol at −20°C. Prior to antibody treatment, fixed germlines were treated with 4% paraformaldehyde for one hour. Slides were blocked in 0.1% PBS Tween with 0.1% BSA for 1 hour, followed by an overnight incubation with 200 μL of KT3 primary antibody (1:100, DSHB). The following day, samples were incubated with a secondary antibody at room temperature for two hours (Jackson ImmunoResearch Labs Cat# 115-605-068 1:500). They were then mounted with Vectashield Antifade Mounting Media with DAPI.
Spinning Disk Confocal imaging
Imaging *C. elegans * embryos and germlines was carried out using a custom-built inverted Zeiss Axio Observer with CSU-W1 SoRa spinning disk scan head (Yokogawa), 4x relay lens (Yokogawa), fast piezo z-drive (Applied Scientific Instrumentation), and an iXon Life 888 EMCCD camera (Andor). Samples were illuminated with 405/637 nm solid-state laser (Coherent), using a 405/640 transmitting dichroic (Semrock) and a 624-40/692-40/525-30/445-45 nm bandpass filter (Semrock). Images were taken using Slidebook software using a 40x-1.3NA objective (Zeiss), with a 4x relay lens (Yokogawa) for embryos, and a 1x relay lens for germlines.
Western Blot
Recombinant proteins were diluted to 1 μM prior to denaturation and loading onto a 4–12% Bis-Tris pre-cast gels (Bio-Rad Hercules, CA). Western blot transfers onto PVDF membranes were run for 1 hour at 4°C. Membranes were blocked for 30 minutes in PBS with 5% milk. The KT3 antibody (DSHB) was diluted by 1:100 and incubated overnight. The secondary incubation occurred for two hours at room temperature (Thermofisher Scientific cat# 62-6720 1:4000, Jackson labs).
** PGL-1 and PGL-3 alignment **
The PGL-1 and PGL-3 C-termini were aligned by running amino acid sequences through MUSCLE (Edgar, 2004) , and then visualized via JALVIEW (Waterhouse et al. 2009) .
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Edgar RC 2004319 MUSCLE: multiple sequence alignment with high accuracy and high throughput.Nucleic Acids Res 3250305-10481792179710.1093/nar/gkh 34015034147 PMC 390337 · doi ↗ · pubmed ↗
- 2Folkmann Andrew W. Putnam Andrea Lee Chiu Fan Seydoux Geraldine 2021910 Regulation of biomolecular condensates by interfacial protein clusters Science 37365600036-80751218122410.1126/science.abg 707134516789 PMC 8627561 · doi ↗ · pubmed ↗
- 3Hanazawa M Yonetani M Sugimoto A 2011314 PGL proteins self associate and bind RN Ps to mediate germ granule assembly in C. elegans.J Cell Biol 19260021-952592993710.1083/jcb.20101010621402787 PMC 3063142 · doi ↗ · pubmed ↗
- 4Kamath RS Fraser AG Dong Y Poulin G Durbin R Gotta M Kanapin A Le Bot N Moreno S Sohrmann M Welchman DP Zipperlen P Ahringer J 2003116 Systematic functional analysis of the Caenorhabditis elegans genome using RN Ai.Nature 42169200028-083623123710.1038/nature 0127812529635 · doi ↗ · pubmed ↗
- 5Kawasaki I Amiri A Fan Y Meyer N Dunkelbarger S Motohashi T Karashima T Bossinger O Strome S 200461 The PGL family proteins associate with germ granules and function redundantly in Caenorhabditis elegans germline development.Genetics 16720016-673164566110.1534/genetics.103.02309315238518 PMC 1470885 · doi ↗ · pubmed ↗
- 6Paix A Folkmann A Seydoux G 201747 Precision genome editing using CRISPR-Cas 9 and linear repair templates in C. elegans.Methods 121-1221046-2023869310.1016/j.ymeth.2017.03.02328392263 PMC 6788293 · doi ↗ · pubmed ↗
- 7Putnam A Cassani M Smith J Seydoux G 201934 A gel phase promotes condensation of liquid P granules in Caenorhabditis elegans embryos.Nat Struct Mol Biol 2631545-999322022610.1038/s 41594-019-0193-230833787 PMC 6668929 · doi ↗ · pubmed ↗
- 8Schmidt H Putnam A Rasoloson D Seydoux G 202169 Protein-based condensation mechanisms drive the assembly of RNA-rich P granules.Elife 1010.7554/e Life.6369834106046 PMC 8238508 · doi ↗ · pubmed ↗