Lack of detection of SARS-CoV-2 in wildlife from Kerala, India in 2020–21
Arun Zachariah, Sajesh P. Krishnankutty, Jishnu Manazhi, Vishnu Omanakuttan, Sam Santosh, Adam Blanchard, Rachael Tarlinton

TL;DR
This study found no evidence of SARS-CoV-2 in various wildlife species from Kerala, India, between 2020 and 2021.
Contribution
The study contributes new geographical data on the absence of SARS-CoV-2 in wildlife in a region with limited prior monitoring.
Findings
SARS-CoV-2 was not detected in lung or gut tissues of 237 wildlife animals from Kerala.
Testing used both pan-coronavirus and Sarbecovirus-specific PCR methods.
Species included bats, primates, civets, and big cats, with no positive results.
Abstract
Spillover of SARS-CoV-2 into a variety of wild and domestic animals has been an ongoing feature of the human pandemic. The establishment of a new reservoir in white-tailed deer in North America and increasing divergence of the viruses circulating in them from those circulating in the human population has highlighted the ongoing risk this poses for global health. Some parts of the world have seen more intensive monitoring of wildlife species for SARS-CoV-2 and related coronaviruses but there are still very large gaps in geographical and species-specific information. This paper reports negative results for SARS-CoV-2 PCR based testing using a pan coronavirus end point RDRP PCR and a Sarbecovirus specific E gene qPCR on lung and or gut tissue from wildlife from the Indian State of Kerala. These animals included: 121 Rhinolophus rouxii (Rufous Horsehoe Bat), six Rhinolophus bedommei (Lesser…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
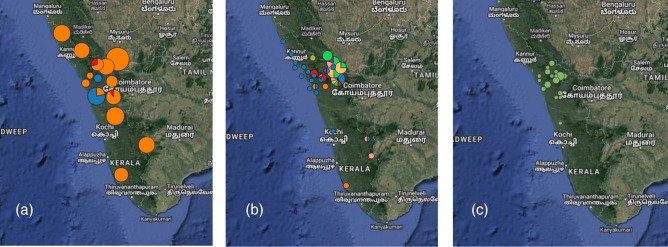 Fig. 1
Fig. 1|
Species |
Lung sample |
Gut sample |
Total no. of animals |
No. of sites |
Sample collection dates |
|---|---|---|---|---|---|
|
| |||||
|
Rufous horseshoe bat |
121 |
98 |
121 |
13 |
May 2021–June 2022 |
|
Lesser woolly horseshoe bat |
6 |
– |
6 |
4 |
May 2021–June 2022 |
|
Fulvous fruit bat |
15 |
– |
15 |
3 |
May 2021–June 2022 |
|
| |||||
|
Bonnet macaque |
47 |
– |
47 |
22 |
Feb 2019–Feb 2022 |
|
| |||||
|
Common palm civet |
35 |
– |
35 |
19 |
Feb 2018–May 2021 |
|
Small Indian civet |
5 |
– |
5 |
4 |
March 2018–March 2020 |
|
Common mongoose |
5 |
– |
4 |
4 |
Sep 2021–March 2022 |
|
Tiger |
10 |
– |
10 |
5 |
April 2020–April 2022 |
|
Leopard |
8 |
– |
8 |
6 |
January 2019–March 2022 |
|
Leopard cat |
4 |
– |
4 |
4 |
January 2021–January 2022 |
|
Jungle cat |
2 |
– |
2 |
2 |
September–May 2021 |
|
Wild dog |
2 |
– |
2 |
2 |
May 2019–Nov 2021 |
|
Sloth |
1 |
– |
1 |
1 |
Archive sample |
|
|
402 |
- —BBSRC
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSARS-CoV-2 and COVID-19 Research · Animal Virus Infections Studies · SARS-CoV-2 detection and testing
Data Summary
Demographic information for the samples described is presented in full in the supplementary information, available in the online version of this article. No other new data or code was generated as part of this project.
Introduction
There have been numerous reports of SARS-CoV-2 spill over from the human pandemic into multiple species. Prominent events with large numbers of animals in multiple sites and spill over back into the human population include domestic cats (Felis Cattus) [1, 2], farmed American mink (Neogale vison) [3, 4] and Syrian hamsters (Mesocricerus auratus) [5, 6]. SARS-CoV-2 has also established ongoing transmission in wild white-tailed deer (Odocoileus virginianus) in the USA, with infection back into the human population confirmed. Worryingly the variants found in the deer population have begun to significantly diverge from those in the human population creating an unpredictable reservoir of novel variants [7–9]. It would also appear from laboratory studies that the range of species able to be infected by SARS-CoV-2 is very dependent on the strain of virus and it is likely that as it continues to evolve in people that the species range of susceptibility will not be stable [10, 11].
There have been a very large number of reports of other species either able to be infected experimentally or with infection detected in sporadic case reports. These are reviewed in [12] but include a large number of cricetid rodents, felids, mustelids, other small carnivores and primates. Many of these reports have been from animals held in zoological collections where they are in close contact with humans, and it is not clear whether these species in their natural environment are at risk or not. Indeed there is a marked contrast in disease transmission between farmed mink at high population density, with almost 100 % of animals infected in a very short period of time in some outbreaks [13] and the sporadic reports, despite intense monitoring, in wild animals, which are largely solitary [14–17]. These behavioural considerations may be as important as biological barriers to which species the virus establishes in.
In addition, we also still have very large gaps in knowledge of the distribution of sarbecoviruses in bats from the Rhinolophoidea; horseshoe bats and roundleaf bats, their natural hosts. There has been intensive sampling of bats in SE Asia, driven by the original SARS-CoV outbreak in 2006 [18]. This effort has established that Sarbecoviruses (SARS like betacoronaviruses) are largely only found in Rhinolophoidid bats. There are however about 180 species of these bats spread across Eurasia and Africa with coronaviruses detected in about 30 of them [19]. Central and South Asia alongside Sub-Saharan Africa are notable absences in Sarbecovirus detection studies [20] with that gap only just beginning to be filled [21–25].
There has been remarkably little study of SARS-CoV-2 in animals in India despite the countries devastating human pandemic [26]. One study in Gujarat (a north western state) of 413 domestic animals of a variety of species reported 23.79 % of animals qPCR positive on nasal or rectal swabs, the positive animals being dogs, cattle and buffalo with sequence confirmation of one canine isolate [27]. Sequencing effort was targeted in areas with a large number of human cases potentially explaining the very high qPCR positivity in this study. A serological study of 320 captive Bengal tigers, Asiatic lions and leopards from eight Indian states demonstrated that 48 (15 %) of these animals had seroconverted to SARS-CoV-2 by October 2021. A small number of Indian elephants (24) and 40 spotted and swamp deer were all seronegative [28]. There have also been reports of PCR positive Asiatic lions in zoos [29] with 2/18 animals in Uttar Pradesh (northern India) and 1/20 in Rajasthan (north west India) qPCR positive on nasal or rectal swabs, with sequence confirmation of the isolates, other felids housed at these institutions did not test positive. Four out of 24 Asiatic lions in Chennai (Tamil Nadu state, south east India) were also found to be qPCR positive and sequence confirmed in a zoo [30], two of these animals died. The only report in a wild animal in India is a solitary juvenile Asiatic leopard found dead in Uttar Pradesh with qPCR positivity and sequence confirmation in [31], this was the only animal out of more than 500 qPCR screened samples positive. In all these cases the felid infections were consistent with the circulating human variants at the time.
India’s size and number of climate zones mean that biodiversity is very high with pressures from the world’s largest human population and known problems with illegal wildlife trade and human/wildlife conflict contributing to multiple zoonotic disease outbreaks [32–34]. The western ghats rainforest along the west coast of India is a biodiversity hotspot with 133 mammal species recorded. It is also an area of intense human wildlife interaction and conflict, with large species such as tigers and elephants causing considerable destruction in human settlements. Consequent to this, zoonotic disease outbreaks are frequent, with the Kyasanur forest and its eponymous virus part of this ecosystem. Surveillance systems and monitoring in this region are however seriously under-resourced with little systematic surveillance of either animals or their viruses [34].
This study sought to partially bridge these gaps with targeted trapping and testing of Rhinolophus bats and opportunistic testing of carnivore and primate species either found dead (roadkill) or culled as part of nuisance animal control activities in the state of Kerala in south west India.
Methods
Sample collection
A total of 260 animals from 13 species (Table 1) were targeted for coronavirus monitoring. For the two horseshoe bat species, palm civets and common mongoose, the targeted numbers were calculated in Epitools [35], two stage sampling for demonstration of disease freedom (cluster size unknown) based on assumption of 5 % prevalence of Coronavirus and 50 % of populations affected. Prior assumptions were based on previous studies of rodent coronaviruses in wild populations [36]. This gave an estimate of seven clusters with 17 individuals in each cluster to be samples (119 animals per species). The species targeted were the two most common horseshoe bats in this environment (others are rare) and the most common small carnivore predators of bats in these sites.
Bats were trapped using mist or harp nets, Subject to inhalational anaesthesia with isoflurane with throat and cloacal swabs collected. Later bats were euthanized by extending the anaesthesia and tissue samples were collected by necropsy. Samples were stored in RNAlater for nucleic acid extraction. Small carnivores (common palm civet, small Indian civet, common mongoose, leopard cat, jungle cat), bonnet macaques and larger carnivores (Bengal tiger, leopard, sloth bear and wild dog), samples were collected as part of routine necropsy procedures from dead animals in the study area. All the carcasses were fresh (within 12 h of death) and samples were preserved in RNA later and stored at −80 degrees Celsius. All procedures were conducted under the supervision of an experienced wildlife veterinarian.
Ethical approval was granted by the University of Nottingham School of Veterinary Medicine and Science Committee for Animal Research and Ethics (CARE). Permission for field work in forest areas for scientific research and sample collection was as per the permit number KFDHQ- 1979/ 2021-CWW / WL 10 issued by the Chief Wildlife Warden, Kerala state, India.
RNA extraction, reverse transcriptase (RT) and RNA-dependent RNA polymerase (RDRP) gene coronaviruses generic conventional PCR
All sample processing and PCR was performed in India at the Kerala state forest department and SciGenom labs, Kerala.
RNA extraction from lung tissue, faecal samples, rectal and oronasal swabs, and cell culture supernatant as positive control, was carried out using the Invitrogen Viral RNA extraction kit as per manufacturer’s instructions. The positive control sample used throughout this study was cDNA from the OC43 Coronavirus ATCC strain VR1558. RT was performed with the Applied Biosystems cDNA reverse transcription kit as per manufacturer’s instructions. All cDNA products were stored at −20 °C for conventional PCR. An endpoint SARS-CoV-2 specific PCR assay [36, 37] was used to amplify the RDRP gene with the Takara R050 A PrimeSTAR GXL taq according to manufacturer’s instructions.
RNA and cDNA quality control was assessed via partial amplification of 108 bp of the beta actin gene using a published conventional PCR protocol [38]. Primers were F: CAGCACAATGAAGATCAAGATCATC and R: CGGACTCATCGTACTCCTGCTT.
Results
No animal sample tested positive for SARS-CoV-2. Locations of samples are shown in Fig. 1.
(a) Locations of bat samples, orange = rufous horseshoe bat, red = lesser woolly horseshoe bat, blue = fulvous fruit bat, pie chart sizes are proportional to the number of animals at each location. (b) Locations of carnivore samples, blue = common palm civet, red = small Indian civet, orange = common mongoose, bright green = tiger, yellow = leopard, pink = leopard cat, purple = jungle cat, light green = wild dog, grey = sloth bear, pie chart sizes are proportional to the number of animals at each location. (c) Locations of bonnet macaque samples, circles are proportional to the number of animals at each location (maps drawn in QGIS v 3.3.1).
Discussion
This study found no evidence of widespread circulation of SARS-CoV-2 or related coronaviruses in Indian wildlife. Some of the species tested here, such as bonnet macaques, palm civets and mongoose are very commonly found in and around human habitation and represent significant pest or nuisance species in terms of aggressive interactions with humans and potential zoonoses or cross species transmission to and from domestic animals [39–41]. These species are high risk for SARS-CoV-2 spill over and it is at least reassuring that these animals tested negative. Though with the large caveats that sampling was PCR based, a small number of animals and could easily have missed infections. Follow up work with serological testing for SARS-CoV-2 antibody (indicating previous infection) would be an extremely useful follow up to this project, with of course the caveat that widely available serological assays have not been validated for these species, making results difficult to interpret [28].
Studies of felids in zoo (captive) populations in India have demonstrated a high rate of seroconversion [28] and PCR positive animals have been detected in zoos [29, 30] and in one wild leopard [31]. Our results here, while a small number of opportunistic samples, add to evidence that SARS-CoV-2 is not a widespread issue in wild Indian felids [31].
A completely negative finding in the two horseshoe bat species was unexpected, particularly as these species are the natural hosts of SARS-like viruses and the PCR assays used in this study should have detected known horseshoe bat sarbecoviruses. Our similar study of UK horseshoe bats did however demonstrate that presence or absence of sarbecoviruses can be very species specific with lesser horseshoe bats having a 44 % positivity rate on faecal or rectal swab samples but no detection at all in greater horseshoe bats [42]. Studies in SE Asia present with very different results with high positivity rates and sarbecoviruses detected in multiple species [43]. Of note the species in which SARS-CoV-2 like sarbecoviruses and recombinant viruses are commonly found, R. sinicus, R. ferrumequinum, R. pusillus, and R. affinis are either rare (R. pusillus) or not found in Kerala. These species are all cave roosting bats that form large colonies which may be a key factor in facilitating sarbecovirus diversity and cross species transmission.
Our sampling numbers and targets should have been able to detect sarbecoviruses in rufous horseshoe bats where target numbers were achieved. Target numbers were not achieved in other species, primarily due to extreme adverse weather conditions (flooding) in Kerala during the sampling period. Most known roost sites for the lesser woolly horseshoe bat (which frequently roosts in sites such as drain coverts) were found abandoned. Trapping success rates for small carnivores were also less than optimal. Nonetheless we present our negative results in the interest of providing the only data to date on Indian horseshoe bat populations. This adds to data indicating that sarbecovirus spill-over out of the horseshoe bat population may be a distinctly regional (SE Asian) phenomena [19, 43].
Supplementary Data
Supplementary material 1Click here for additional data file.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Piewbang C Poonsin P Lohavicharn P Wardhani SW Dankaona W et al SARS-Co V-2 transmission from human to pet and suspected transmission from pet to human, Thailand J Clin Microbiol 202260 e 010582210.1128/jcm.01058-2236314788 PMC 9667761 · doi ↗ · pubmed ↗
- 2Tyson GB Jones S Logan N Mc Donald M Marshall L et al SARS-Co V-2 seroprevalence and cross-variant antibody neutralization in cats, United Kingdom Emerg Infect Dis 2023291223122710.3201/eid 2906.22175537141617 PMC 10202862 · doi ↗ · pubmed ↗
- 3Oude Munnink BB Sikkema RS Nieuwenhuijse DF Molenaar RJ Munger E et al Transmission of SARS-Co V-2 on mink farms between humans and mink and back to humans Science 202137117217710.1126/science.abe 590133172935 PMC 7857398 · doi ↗ · pubmed ↗
- 4Wasniewski M BouéF Richomme C Simon-Lorière E Van der Werf S et al Investigations on SARS-Co V-2 and other coronaviruses in mink farms in France at the end of the first year of COVID-19 pandemicbio Rxiv 20232023.02.02.52674910.1101/2023.02.02.52674937624818 PMC 10456147 · doi ↗ · pubmed ↗
- 5Kok K-H Wong S-C Chan W-M Wen L Chu AW-H et al Co-circulation of two SARS-Co V-2 variant strains within imported pet hamsters in Hong Kong Emerg Microbes Infect 20221168969810.1080/22221751.2022.204092235135441 PMC 8890519 · doi ↗ · pubmed ↗
- 6Yen H-L Sit THC Brackman CJ Chuk SSY Gu H et al Transmission of SARS-Co V-2 delta variant (AY.127) from pet hamsters to humans, leading to onward human-to-human transmission: a case study Lancet 20223991070107810.1016/S 0140-6736(22)00326-935279259 PMC 8912929 · doi ↗ · pubmed ↗
- 7Caserta LC Martins M Butt SL Hollingshead NA Covaleda LM et al White-tailed deer (Odocoileus virginianus) may serve as a wildlife reservoir for nearly extinct SARS-Co V-2 variants of concern Proc Natl Acad Sci U S A 2023120 e 221506712010.1073/pnas.221506712036719912 PMC 9963525 · doi ↗ · pubmed ↗
- 8Mc Bride D Garushyants S Franks J Magee A Overend S et al Accelerated evolution of SARS-Co V-2 in free-ranging white-tailed deer Res Sq 2023 rs.3.rs-257499310.21203/rs.3.rs-2574993/v 137640694 PMC 10462754 · doi ↗ · pubmed ↗