Expansion of the split hygromycin toolkit for transgene insertion in Caenorhabditis elegans
Megan J Moerdyk-Schauwecker, Erin K Jahahn, Zachariah I Muñoz, Kristin J Robinson, Patrick C Phillips

TL;DR
This paper expands a genetic tool for inserting transgenes in C. elegans, adding new chromosomal locations and natural isolates for better germline expression.
Contribution
The split hygromycin system is extended to new chromosomal sites and natural isolates, enabling more flexible transgene insertion.
Findings
Two new chromosomal locations permissive for germline expression were added to the split hygromycin system.
Engineered landing pads were created in three additional natural isolates of C. elegans.
Guide and repair template plasmids are now available for all expanded sites.
Abstract
Engineered sites for genetic transformation have simplified transgene insertion in Caenorhabditis elegans . These strategies include our split hygromycin system (Stevenson et al. 2020) which allows for integration-specific selection of transgenes. Here we have expanded the split hygromycin selection system to include two additional chromosomal locations, both of which are permissive for germline expression, as well as engineered landing pads in three additional natural isolates. Corresponding guide and empty repair template plasmids are also available for each of these sites.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
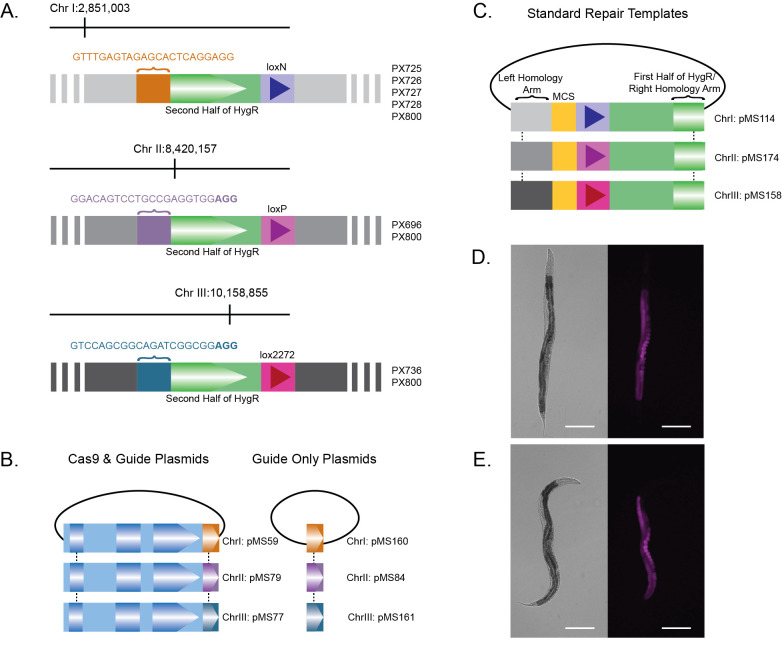 Figure 1
Figure 1|
|
|
|
|
|
|
|
NA |
Wild Isolate |
|
Upon request |
|
|
NA |
Wild Isolate |
|
CaeNDR |
|
|
NA |
Wild Isolate |
|
CaeNDR |
|
|
NA |
Wild Isolate |
|
CaeNDR |
|
|
|
|
|
CGC |
|
|
|
|
|
CGC |
|
|
|
|
This work |
CGC |
|
|
|
|
This work |
CGC |
|
|
|
|
This work |
CGC |
|
|
|
|
This work |
CGC |
|
|
|
|
This work |
CGC |
|
|
|
|
This work |
Upon request |
|
|
|
|
This work |
Upon request |
|
|
|
|
This work |
Upon request |
|
|
|
|
This work |
Upon request |
|
|
|
|
This work |
Upon request |
|
|
|
|
This work |
Upon request |
|
|
|
|
This work |
Upon request |
|
|
|
|
This work |
Upon request |
|
|
|
|
|
|
|
pMS87 |
Landing pad insertion plasmid for ChrI |
5'HA + synthetic guide site1 + 3'ΔHygR::
|
This work |
Addgene |
|
pMS110 |
Landing pad insertion plasmid for ChrIII |
5'HA + synthetic guide site3 + 3'ΔHygR:
|
This work |
Addgene |
|
pMS18 |
Cas9 + guide plasmid for inserting ChrI landing pad |
|
|
Addgene |
|
pMS62 |
Cas9 + guide plasmid for inserting ChrIII landing pad |
|
|
Addgene |
|
pMS114 |
Empty repair plasmid for ChrI |
5'HA + MCS + loxN +
|
This work |
Addgene |
|
pMS178 |
Empty repair plasmid for ChrII |
5'HA + MCS + loxP +
|
This work |
Addgene |
|
pMS158 |
Empty repair plasmid for ChrIII |
5'HA + MCS + lox2272 +
|
This work |
Addgene |
|
pMS59 |
Cas9 + guide plasmid targeting ChrI landing pad |
|
|
Addgene |
|
pMS79 |
Cas9 + guide plasmid targeting ChrII landing pad |
|
|
Addgene |
|
pMS77 |
Cas9 + guide plasmid targeting ChrIII landing pad |
|
|
Addgene |
|
pMS160 |
Guide only plasmid targeting chrI landing pad |
U6p::GTTTGAGTAGAGCACTCAGG |
This work |
Addgene |
|
pMS84 |
Guide only plasmid targeting chrII landing pad |
U6p::GGACAGTCCTGCCGAGGTGG |
|
Addgene |
|
pMS161 |
Guide only plasmid targeting chrIII landing pad |
U6p::GTCCAGCGGCAGATCGGCGG |
This work |
Addgene |
|
pMS143.1 |
Reporter for insertion into chrI landing pad |
5'HA +
|
This work |
Upon request |
|
pMS81 |
Reporter for insertion into chrII landing pad |
5'HA +
|
|
Addgene |
|
pMS144 |
Germline reporter for insertion into chrI landing pad |
5'HA +
|
This work |
Upon request |
|
pMS145 |
Germline reporter for insertion into chrIII landing pad |
5'HA +
|
This work |
Upon request |
|
pCFJ421 |
Array marker; source for cloning components |
|
(Frøkjær-Jensen et al. 2012) |
Addgene |
|
pMS30 |
Source for cloning components |
5'HA +
|
|
Upon request |
|
pMS63 |
Source for cloning components |
5'HA +
|
|
Upon request |
|
pMS70 |
Source for cloning components |
5'HA + synthetic guide site1 + 3'ΔHygR::
|
|
Upon request |
|
pMS72 |
Source for cloning components |
5'HA + synthetic guide site3 + 3'ΔHygR::
|
|
Upon request |
|
pZCS11 |
Empty vector for guide expression |
U6p::Scaffold |
|
Upon request |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms · CRISPR and Genetic Engineering · Insect Resistance and Genetics
Description
Several strategies have been developed utilizing engineered “safe harbor” or “landing pad” sites for transgene insertion in Caenorhabditis elegans (Silva-García et al. 2019; Nonet 2020, 2021, 2023; Stevenson et al. 2020, 2023; Yang et al. 2020; Mouridi et al. 2022; Malaiwong et al. 2023). These sites facilitate efficient introduction of transgenes by providing a means for selection of integrants and/or simplifying the required insertion template. We have previously demonstrated split hygromycin selection as a simple, rapid and highly generalizable system for transgene insertion in C. elegans (Stevenson et al. 2020) . This system splits the hygromycin resistance ( HygR ) gene between the landing pad site and the repair template, meaning that only individuals with an integrated transgene, and not array bearing individuals, have resistance to the drug hygromycin B and can therefore be identified by a simple live/dead screen. Here we expand this system (1) to additional chromosomal locations, allowing for generation of more complex transgenics; and (2) to several commonly used C. elegans natural isolates, allowing for generation and comparison of transgenics in multiple genetic backgrounds.
Previously we introduced a single copy, split hygromycin landing pad into chromosome II of N2 ( PX696 ) (Stevenson et al . 2020). Here, using the same self-excising cassette (SEC) based CRISPR strategy, we also introduced a single copy, split hygromycin landing pad into chromosome I of N2 at the same intergenic region occupied by the * ttTi4348 * transposon utilized in MosSCI (Frøkjær-Jensen et al. 2012) and into chromosome III of N2 between the * nac-1 * and * K08E5.5 * genes, thereby creating strains PX725 and PX736 respectively ( Figure 1A ). Both of these locations have also been previously utilized for direct transgene insertion (Kasimatis et al. 2018, 2022) . Each of these three landing pads contain the 3’ portion of the coding sequence for the HygR gene and a *unc-54 * 3’ UTR flanked by a lox site. This allows for repair template integration specific selection of transgenic individuals and for subsequent optional removal of *HygR * by expression of Cre recombinase (Stevenson et al. 2020) . A unique lox sequence is used for each landing pad site, allowing them to be used together without triggering interchromosomal recombination (Lee and Saito 1998; Langer et al. 2002) .
Each landing pad also contains a unique, high-efficiency synthetic guide site (Stevenson et al. 2020) , that is targeted for insertion of transgenic cargo. This guide site can be targeted by injection of either a plasmid expressing the guide plus Cas9 or a plasmid expressing guide alone plus an independent source of Cas9 ( Figure 1B ). This Cas9 can come from either a separate plasmid, purified protein or a strain expressing integrated Cas9 [for example (Zhang et al. 2018; Schwartz et al. 2021) ]. We demonstrated that the landing pads can be used in conjunction with integrated Cas9 by crossing PX696 and PX725 with the Cas9 bearing strain EG9882 to create PX783 and PX784 respectively. PX736 is incompatible with EG9882 due to both strains possessing a lox2272 site. A rpl-28p::mKate2 transcriptional reporter was then integrated into the PX783 and PX784 strains by CRISPR to generate PX798 and PX792 . In screening for integrants, we noticed a high frequency of removal of the HygR gene used for selection, even in the absence of heat shock, likely due to leaky expression of the
- hsp-16.41p::Cre* construct also present in EG9882 . As such, it may be possible to achieve higher integration efficiencies if integrated Cas9 strains lacking Cre are used.
For each landing pad a corresponding empty repair template plasmid was also made ( Figure 1C ). These plasmids each contain homology to the landing pad on either side of the cut site, a multiple cloning site (MCS) to facilitate linearization of the vector (although this can also be done by PCR), the * rps-0 * promoter proceeded by a lox sequence matching that in the landing pad, and the 5’ portion of the HygR coding sequence. While we created generic, empty repair templates, repair templates could also be customized for specific purposes. In principle, it should be possible to combine the split hygromycin selection with SEC (Dickinson et al. 2015) , thereby bypassing the need for an outside source of Cre for removal of the HygR gene. While it does indeed work to include the additional SEC components in the repair template, for reasons that are unclear, transgene insertion using this repair template is not only significantly lower than the base split hygromycin system, but also lower than that achieved using the original SEC protocol.
Since all three landing pads have been designed to be compatible with each other and are individually targetable, they can be combined to make more complex recombinants either through crosses or by sequentially inserting genes into a strain containing two or more landing pads, although in this case removal of the restored HygR gene is required to restore the drug-sensitive background prior to the next round of editing. In principle, it is also possible to take a co-CRISPR like approach and simultaneously target two or more sites as editing at one site increases the probability of editing at additional sites even when using large repair templates (Stevenson et al. 2023) . To facilitate these latter approaches, we have generated PX729 , which contains the chromosome I and II sites, and PX800 , which contains all three landing pad sites, in an N2 background.
One of the advantages of landing pads generated via CRISPR is that as long as the guide site sequence is conserved in the genome, the same reagents can be used to integrate a landing pad into any C. elegans genetic background. This creates a path for a more general study of background specific effects in individual alleles [i.e. epistasis (Phillips 2008) ]. Similarly, since the landing pad creates a standardized guide and homology arm set, transgenes can also be integrated into any particular landing pad using the same reagents regardless of genetic background. To demonstrate this and to create landing pads in several commonly used strains as a resource for the community, we integrated the chromosome I landing pad into MY16 , CB4856 and JU775 to create PX726 , PX727 and PX728 respectively. These strains have been used in multiple quantitative trait locus (QTL) mapping experiments (Andersen and Rockman 2022) , as well as being core component of the *Caenorhabditis * Intervention Testing Program (CITP) (Lucanic et al. 2017) .
Finally, one limitation to transgenesis in C. elegans is that transgene expression can be restricted in the germline, and this effect is based in part on chromosome location (Frøkjær-Jensen et al. 2012). Therefore, it is important to determine the germline permissiveness of any new transgene insertion site. The chromosome II landing pad has been previously shown to be permissive for germline expression (Stevenson et al. 2020) . To test the germline permissiveness of chromosome I and III landing pads, a mex-5p::mKate2 reporter was integrated into each. In both cases, robust fluorescence could be detected in the germline ( Figure 1D and E ), indicating that these landing pads are germline permissive. For the chromosome I landing pad, this is consistent with previous work using the * ttTi4348 * MosSCI site (Frøkjær-Jensen et al. 2012).
Overall, these new strains and reagents expand the *C. elegans * split hygromycin toolkit and allow integration of genetic elements at multiple independent sites in the genome and across multiple genetic backgrounds.
Methods
Worm maintenance
All strains were maintained at 15⁰C on nematode growth media (NGM) seeded with Escherichia coli OP50 unless otherwise specified.
Plasmids
Guide plus Cas9 plasmids have been previously described (Kasimatis et al . 2018, 2022; Stevenson et al . 2020). Guide only plasmids were generated by site directed mutagenesis of pZCS11 (Stevenson et al. 2023) using the Q5 site-directed mutagenesis kit [New England Biolabs (NEB), E0554S)]. Empty repair templates were generated from an insert-containing repair template using site directed mutagenesis to remove the insert and replace it with an MCS.
The landing pad insertion plasmids and insert-containing repair templates were generated by Gibson cloning using NEBuilder HiFi DNA assembly master mix (NEB, E2621). Cloning fragments were made either by restriction digest of existing vectors or by PCR using Q5 high-fidelity 2x master mix (NEB, M0492).
For the chromosome I landing pad insertion plasmid (pMS87), the backbone and homology arms were amplified from pMS30 (Kasimatis et al. 2018) and the split HygR landing pad and SEC with loxN sites added by PCR were amplified from pMS70 (Stevenson et al. 2020) . For the chromosome III landing pad insertion plasmid (pMS110), the backbone and homology arms were amplified from pMS63 (Kasimatis et al. 2022) and the split HygR landing pad and SEC with lox2272 sites added by PCR were amplified from pMS72 (Stevenson et al . 2020).
The chromosome II repair template containing rpl-28p::mKate2 (pMS81) has been previously described (Stevenson et al. 2020) . pMS81 was also used as the source of the plasmid backbone, split HygR and rpl-28p::mKate2 source for the corresponding chromosome I repair template (pMS143.1), while the genomic homology arm was amplified from pMS87 and the loxN site was added by synthesis.
For both the chromosome I and chromosome III mex-5p::mKate2 repair templates (pMS144 and pMS145 respectively), mex-5p was amplified from genomic DNA, mKate2 was amplified from pMS81 and the tbb-2 3’ UTR was amplified from pCFJ421 (Frøkjær-Jensen et al. 2012). The remaining components of pMS144 were amplified from pMS114. For pMS145 the genomic homology arm was amplified from pMS63 while with backbone and split HygR site were amplified from pMS81 with addition of the lox2272 site by PCR.
All plasmids were purified using the ZR plasmid miniprep classic kit (Zymo Research, D4016) and verified by sequencing prior to use. Plasmid sequences are available through Addgene or upon request.
Transgenic generation
To make single copy insertions of the landing pads, young adult hermaphrodites of the desired background strain were injected with a mixture consisting of 10ng/µl repair template plasmid (pMS87 or pMS110), 50ng/µl guide plus Cas9 plasmid (pMS18 or pMS62) and 2.5ng/µl pCFJ421 that served as an array marker. Following injection, worms were maintained at 25°C. Selection, screening, and SEC removal were done as in (Dickinson et al. 2015) .
For insertion of germline reporters into the landing pads, young adult PX725 or PX736 hermaphrodite were injected with a mixture consisting of 15ng/µl repair template plasmid (pMS144 or pMS145) and 50ng/µl guide plus Cas9 plasmid (pMS59 or pMS77). To test landing pad insertions using integrated Cas9, young adult PX783 or PX784 hermaphrodites were injected with a mixture consisting of 15ng/µl repair template plasmid (pMS81 or pMS143.1) and 25ng/µl guide only plasmid (pMS84 or pMS160)
In both cases, following injection, worms were maintained at 25°C and dosed with hygromycin B at a final concentration of 250µg/ml approximately 48 hours later. Five to seven days after dosing, candidate worms were singled and genotyped by PCR.
Genotyping of candidate individuals was done by single worm PCR using OneTaq quick-load 2x master mix with standard buffer (NEB, M0486). All transgene insertions were confirmed by Sanger sequencing. Sequences of genotyping/sequencing primers are available upon request. All strains are available through CGC, CaeNDR or upon request.
Microscopy
Microscopy images were obtained with the 10x objective on an Olympus IX inverted fluorescent microscope using the Olympus cellSens Dimension 2.3 capture software and an Andor Zyla sCMOS camera. Worms were mounted on a 2% agarose pad and immobilized using 100mM sodium azide. Fluorescent images were captured using the mCherry settings with an exposure time of 10 ms. Light images were captured using the DIC filter set. Fluorescent images were recolored using ImageJ2 version 2.9.0.
Reagents
**: **
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Andersen EC Rockman MV 202214 Natural genetic variation as a tool for discovery in Caenorhabditis nematodes.Genetics 22010016-673110.1093/genetics/iyab 15635134197 PMC 8733454 · doi ↗ · pubmed ↗
- 2Crombie TA Mc Keown R Moya ND Evans KS Widmayer SJ La Grassa V Roman N Tursunova O Zhang G Gibson SB Buchanan CM Roberto NM Vieira R Tanny RE Andersen EC 202415 Cae NDR, the Caenorhabditis Natural Diversity Resource.Nucleic Acids Res 52D 10305-1048 D 850D 85810.1093/nar/gkad 88737855690 PMC 10767927 · doi ↗ · pubmed ↗
- 3Dickinson DJ Pani AM Heppert JK Higgins CD Goldstein B 201563 Streamlined Genome Engineering with a Self-Excising Drug Selection Cassette.Genetics 20040016-67311035104910.1534/genetics.115.17833526044593 PMC 4574250 · doi ↗ · pubmed ↗
- 4El Mouridi S Alkhaldi F Frøkjær-Jensen C 2022825 Modular safe-harbor transgene insertion for targeted single-copy and extrachromosomal array integration in Caenorhabditis elegans.G 3 (Bethesda)12910.1093/g 3journal/jkac 18435900171 PMC 9434227 · doi ↗ · pubmed ↗
- 5Frøkjær-Jensen C Davis MW Ailion M Jorgensen EM 2012130 Improved Mos 1-mediated transgenesis in C. elegans.Nat Methods 921548-709111711810.1038/nmeth.186522290181 PMC 3725292 · doi ↗ · pubmed ↗
- 6Kasimatis KR Moerdyk-Schauwecker MJ Phillips PC 2018731 Auxin-Mediated Sterility Induction System for Longevity and Mating Studies in Caenorhabditis elegans . G 3 (Bethesda)882655266210.1534/g 3.118.20027829880556 PMC 6071612 · doi ↗ · pubmed ↗
- 7Kasimatis KR Moerdyk-Schauwecker MJ Lancaster R Smith A Willis JH Phillips PC 2022214 Post-insemination selection dominates pre-insemination selection in driving rapid evolution of male competitive ability.P Lo S Genet 1821553-7390 e 1010063 e 101006310.1371/journal.pgen.101006335157717 PMC 8880957 · doi ↗ · pubmed ↗
- 8Langer SJ Ghafoori AP Byrd M Leinwand L 2002715 A genetic screen identifies novel non-compatible lox P sites.Nucleic Acids Res 30140305-10483067307710.1093/nar/gkf 42112136089 PMC 135742 · doi ↗ · pubmed ↗