The CRISPR/Cas9 system forms a condensate in the yeast nucleus
Sara Medina-Suárez, Félix Machín

TL;DR
The study shows that the CRISPR/Cas9 system forms a condensate in the nucleus of yeast cells, which could help improve gene editing techniques.
Contribution
The novel contribution is the observation that Cas9 forms a condensate in the yeast nucleus, which co-localizes with the nucleolus.
Findings
Cas9 accumulates in the nucleus of yeast cells over time.
Cas9 forms a non-uniform condensate in the nucleoplasm.
The condensate co-localizes with the nucleolus and targets the site to its periphery.
Abstract
CRISPR/Cas9 gene editing technology has revolutionized genetic engineering. However, the nuclear dynamics of Cas9 in eukaryotic cells, particularly in the model organism Saccharomyces cerevisiae , remains poorly understood. Here, we constructed yeast strains expressing fluorescently tagged Cas9 variants, revealing their accumulation in the nucleus over time. Notably, Cas9 was non-uniformly distributed in the nucleoplasm during the initial hours, suggesting the formation of a condensate. This condensate often co-localizes with the nucleolus and associates the target site to its periphery. Our findings provide insights into Cas9 nuclear dynamics in yeast, advancing our understanding of CRISPR/Cas9-based genetic manipulation.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
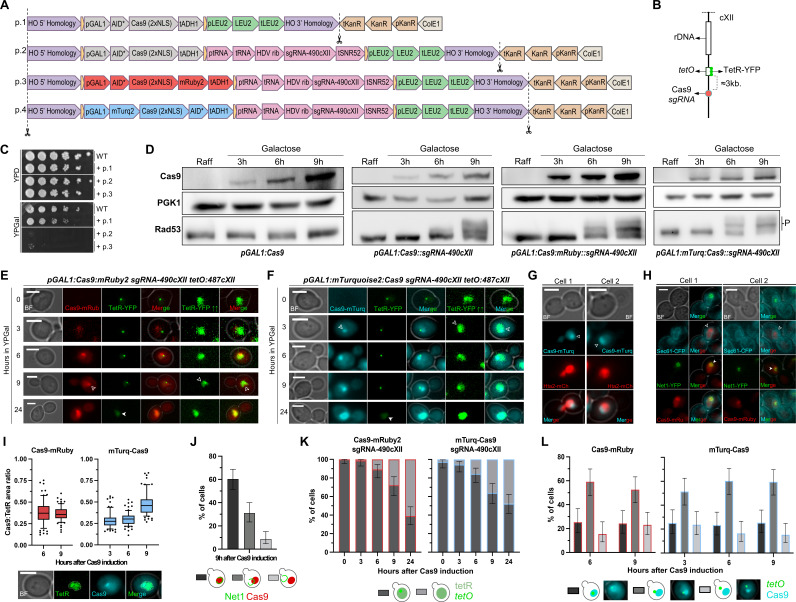 Figure 1
Figure 1|
|
|
|
|
|
FM1249 |
|
F.Machín lab |
1C |
|
FM2664 |
FM1249;
|
This work |
1C, 1D, 1K |
|
FM2643 |
FM1249;
|
This work |
1C, 1D |
|
FM2644 |
FM1249;
|
This work |
1C, 1D, 1E, 1I, 1K, 1L |
|
FM2710 |
FM1249;
|
This work |
1D, 1F, 1I, 1K, 1L |
|
FM3203 |
|
This work |
1G |
|
FM3205 |
|
This work |
1H, 1J |
|
|
|
|
|
pFM431 (p.1) |
pFM427 as the single TU (
|
MoClo plasmid to insert
|
|
pFM432 (p.2) |
pFM428 as the first TU (
|
MoClo multigene plasmid to insert
|
|
pFM433 (p.3) |
pFM429 as the first TU (
|
MoClo multigene plasmid to insert
|
|
pFM434 (p.4) |
pFM430 as the first TU (
|
MoClo multigene plasmid to insert
|
|
pFM422 |
CAS9-3b-stop PCR product inserted in the pYTK001 part plasmid |
Part Plasmid with
|
|
pFM423 |
CAS9-3b-nostop PCR product inserted in the pYTK001 part plasmid |
Part Plasmid with
|
|
pFM424 |
sgRNA-CXII490-F/R dsDNA inserted in the pYTK050 plasmid |
RNA pol III transcriptional unit for
|
|
pFM425 |
Synthetic
|
Part Plasmid carrying
|
|
pFM426 |
AID-4a PCR product inserted in the pYTK001 part plasmid |
Part Plasmid carrying
|
|
|
|
|
Cas9-3b-F |
GCATCGTCTCATCGGTCTCATTCTGACAAGAAGTATTCTATCGGACTG |
|
Cas9-3b-R-stop |
ATGCCGTCTCAGGTCTCAGGATCAGGATCCTTATACCTTTCTCTTC |
|
Cas9-3b-R-nostop |
TGCCGTCTCAGGTCTCAGGATCCTACCTTTCTCTTCTTTTTTGGATCTACC |
|
sgRNA-cXII490-F |
GACTTTCCGCCCCAGCCAAACTCTCC |
|
sgRNA-cXII490-R |
AAACGGAGAGTTTGGCTGGGGCGGAA |
|
AID-4a-F |
GCATCGTCTCATCGGTCTCAATCCCCTAAAGATCCAGCCAAACCTC |
|
AID-4a-R |
ATGCCGTCTCAGGTCTCAGCCATTACTTCACGAACGCCGCCGC |
|
|
|
|
|
|
|
anti-Pgk1 |
Primary |
Mouse monoclonal |
Thermo Fisher Sci. |
22C5D8 |
|
anti-Rad53 |
Primary |
Mouse monoclonal |
Abcam |
ab166859 |
|
anti-CRISPR-Cas9 |
Primary |
Rabbit monoclonal |
Abcam |
ab189380 |
|
anti-Mouse (HRP) |
Secondary |
Goat polyclonal |
Promega |
W4021 |
|
anti-Rabbit (HRP) |
Secondary |
Goat polyclonal |
Abcam |
ab97051 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCRISPR and Genetic Engineering · Genetics, Aging, and Longevity in Model Organisms · RNA and protein synthesis mechanisms
Description
The advent of the Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) and CRISPR-associated protein 9 (Cas9) system has deeply transformed genetic engineering and genome editing, enabling precise and targeted modifications of DNA sequences in a wide range of organisms (Doudna and Charpentier 2014) . Cas9 is an endonuclease responsible for generating DNA double-strand breaks at specific target sites defined by a short guide RNA (sgRNA). However, while Cas9 has been extensively employed for genome editing, much remains to be elucidated regarding its intracellular behavior and dynamic localization within eukaryotic cells. Microscopy techniques have been invaluable for visualizing the spatiotemporal distribution of proteins within living cells. Studies of Cas9 localization in mammalian cells have been mostly focused on its use to visualize specific loci. In this context, an endonuclease-dead mutant (dCas9) is used, and the distribution includes a pool of dCas9 bound to the target sequence(s) together with another pool that is uniformly distributed throughout the nucleoplasm but enriched at nucleoli (Chen et al., 2013) . However, this pattern is not universal as, for example, only the target-bound pool is present in Caenorhabditis elegans embryos (Memar et al., 2022) . Of note, the localization of Cas9 in Saccharomyces cerevisiae cells, a classical model organism for eukaryotic genetics, remains comparatively less explored and, to the best of our knowledge, has only been studied in relation to the optimization of nucleocytoplasmic shuttling strategies (Roggenkamp et al., 2018) . Now, we have addressed this crucial knowledge gap by a time course exploration of Cas9 localization within the yeast nucleus.
To this end, we constructed four yeast strains expressing yeast-optimized Cas9 variants assembled using the Golden Gate modular cloning (MoClo) method (Lee et al., 2015) . One strain expresses only the basic Cas9 protein under the inducible GAL1 promoter ( Figure 1A, p . 1). A second strain also co-expresses an sgRNA against a genomic region 3 Kb away from a tetO array inserted at the 487 Kb coordinate in chromosome XII ( sgRNA-490cXII ) ( Figure 1A, p .2; Figure 1B ). This tetO array is a tool to detect this particular genomic region as a spot through its specific binding to the TetR reporter, which in turn is fused to the fluorescent protein YFP (yellow/green fluorescence) (Machín et al., 2005). Importantly, not all TetR-YFP molecules bind the tetO array; an unbound pool freely and uniformly circulates in the nucleus. The third strain is a derivative of the latter in which Cas9 is fused at the C-terminus to the fluorescent protein mRuby2 (red fluorescence) ( Figure 1A, p . 3), while the fourth strain has the fluorescent protein mTurquoise2 (cyan fluorescence) fused to the N-terminus of Cas9 ( Figure 1A, p . 4). Functional analysis by spot assays showed that co-expression of Cas9 constructs and sgRNA-490cXII completely inhibited cell growth ( Figure 1C, p .2 or p.3 in YPgal), whereas neither Cas9 alone (p.1 in YPgal) nor sgRNA-490cXII alone (p.2 or p.3 in YPD) were toxic. Correspondingly, evidence of Cas9-mediated DNA damage, as measured by hyperphosphorylation of the DNA damage checkpoint reporter Rad53, was only observed when the Cas9 variants were co-expressed with sgRNA-490cXII ( Figure 1D ). The timing of Rad53 hyperphosphorylation in a time course of Cas9 expression showed a delay of approximately 3 h relative to the appearance of Cas9 (6 h and 3 h from induction, respectively) ( Figure 1D ).
Microscopic detection of the fluorescently tagged Cas9 variants showed that Cas9-mRuby2 was not visible 3 h after Cas9 induction, although Cas9 was clearly detected by Western blot at the same time point ( Figure 1D and 1E). By contrast, there was good agreement between microscopy and Western blots for mTurquoise2-Cas9 detection ( Figure 1D and 1F). The delay in fluorescence detection of Cas9-mRuby2 is likely related to the slow maturation of red fluorescent proteins (Balleza et al., 2018) . Both Cas9-mRuby2 and mTurquoise2-Cas9 accumulated in the yeast nucleus over time, and an intense signal covered the entire nucleoplasm by 24 h ( Figure 1E and 1F). Interestingly, however, the distribution of Cas9 was not uniform during the first 9 h, with Cas9 confined to less than 25%-50% of the nucleoplasmic area, as reported by the freely circulating pool of NLS-TetR-YFP, the nuclear DNA histone Hta2, and the nuclear envelope marker Sec61 ( Figure 1E -I). The fact that Cas9 is not uniformly distributed across the nucleoplasm strongly suggests that the Cas9 sgRNA partner forms a condensate, as has been proposed for the yeast nucleolus (Lawrimore et al., 2021) . Based on this similarity and the fact that Cas9 is enriched in human nucleoli (Chen et al., 2013) , we also examined whether the Cas9 condensate associates with the yeast nucleolus. We found partial co-localization with the nucleolar marker Net1 ( Figure 1H,J ). In ~60% of cells the Cas9 condensate perfectly colocalizes with Net1, whereas in ~30% of cells Cas9 and Net1 partially co-localized. Only in ~8% of cells did Net1 and Cas9 clearly occupy distinct nuclear subdomains. Thus, there appears to be a tendency for Cas9 to condensate in the yeast nucleolus.
Finally, we asked whether the Cas9 condensate nucleates around the expected cut site in the genome. If this were the case, we would expect to see Cas9 surrounding the tetO /TetR spot. Note that the percentage of cells with visible spots decreased over the time course of Cas9/ sgRNA-490cXII co-expression ( Figure 1K ), likely as a result of resection of the Cas9-generated double-strand break (Mosbach et al., 2020) . Of those cells where the spot was visible, approximately 25% had the spot surrounded by Cas9 ( Figure 1L ). This percentage is similar to the subnuclear area covered by the Cas9 condensate and is therefore consistent with the expected percentage by chance. However, the tetO spot did not follow a random location in the remaining cells. In fact, in up to 60% of cells overall (~80% of cells with the tetO not surrounded by Cas9), the tetO spot is located at the periphery of the Cas9 condensate ( Figure 1L ), with no differences in distribution between Cas9-mRuby2 and mTurquoise2-Cas9. This result suggests that the genomic region to be cut may stably contact the periphery of the Cas9 condensate. However, since the cutting site was designed near the ribosomal DNA array ( Figure 1B ), further work will be required to discern the actual cause of the Cas9-nucleolus-target site association.
In conclusion, Cas9 is not homogeneously distributed in the yeast nucleoplasm, but forms a condensate that occupies one third of the nuclear area. Interestingly, this condensate does not assemble onto the target site in the genome, but around the nucleolus, although this site is attached to the condensate periphery. Our investigation fills a knowledge gap that not only advances our understanding of CRISPR/Cas9-based genetic manipulation in yeast, but may also contribute to a broader understanding of Cas9 behavior across eukaryotic species.
Methods
Yeast strains and plasmids
Most yeast strains used stem from FM1249, which is derived from the YPH499 background. FM1249 carries several genetic modifications, including: (i) a fluorescent tag for the distal part of the rDNA array on the right arm of chromosome XII ( tetO:487 /NLS-TetR-YFP system; NLS, nuclear location signal), (ii) the cdc14-1 thermosensitive allele, which arrests cells in early anaphase at the restrictive temperature of 37 ºC (Machín et al., 2016), and (iii) an integrated construct to constitutively express the auxin-inducible degron system (OsTIR1) (Morawska and Ulrich 2013) . FM3203 and FM3205 are wild-type strains from the W303 background that carry fluorescent tags for nuclear reporters apart from expressing Cas9.
Different multigene plasmids carrying transcriptional units (TU) for expressing a yeast-optimized Cas9 under the control of the GAL1 promoter ( GAL1p ) were generated by the MoClo Yeast Toolkit (Lee et al., 2015) ( Figure 1A ). The ADH1 terminator ( ADH1t ) was used in all TUs. The toolkit already provides a cloning module for Cas9 as the sole open reading frame between the promoter and the terminator (type 3 module, pYTK036). In order to add tags at both the N- and C-terminus, we made a Cas9 type 3b module (see plasmid table in Reagents) by PCR using a set of primers with the corresponding tails (see primer table in Reagents). In this manner, Cas9 was fused to fluorescent proteins in the corresponding TUs ( Figure 1A ). Likewise, the minimal auxin-inducible degron ( AID* ) sequence was tagged to the second terminus of Cas9 to make Cas9 degradable. The AID* sequence, obtained from the plasmid pKan-AID*-9myc (addgene code #99522; #2188 in the Urlich collection) (Morawska and Ulrich 2013) , was converted into a 4a module by PCR, whereas an AID* 3a module was ordered as a synthetic gene (gBlocks HiFi Gene fragments, IDT). In these multigene plasmids, a second TU was assembled to include an sgRNA designed to cut at coordinate 490 Kb on the chromosome XII right arm, just 3 Kb away from the tetO:487 fluorescent tag. Finally, the multigene cassette was designed to be flanked by sequences homologous to the ho locus on yeast chromosome IV. In all cases, the different transcriptional units were assembled using pYTK095 as a backbone (bacterial AmpR-ColE1 type 678 module). If the final plasmid has only one TU, it is flanked by LS and RE connectors; if it has two, the first TU is flanked by LS and R1 connectors, while the second is flanked by L1 and RE.
Integration of Cas9 variants and sgRNA was achieved by releasing the cassette by cutting with NotI-HF (NEB, R3189L) ( Figure 1A ), followed by standard transformation procedures (Knop et al., 1999) .
Experimental conditions
Strains were grown overnight in air orbital incubators at 25 ºC in YP media (10 g·L ^−1^ yeast extract, 20 g·L ^−1^ peptone) with raffinose 2% (w/v) as carbon source, then galactose 2% (w/v) was added to log-phase asynchronous cultures whose OD 600 was previously adjusted to ~ 0.4. Samples were taken at different time points during 24 h for Western blotting and/or wide field fluorescence microscopy. Neither the early anaphase arrest property of the cdc14-1 allele nor AID*-mediated conditional degradation by auxin in FM1249-based strain was tested in this work. At 25º C, Cdc14-1 is functional in this background. The W303-based strains carry a wild type CDC14 gene.
Western blotting and microscopy were carried out as reported before (Matos-Perdomo et al., 2022) . A fully motorized Leica DMI6000B microscope with an ultra-sensitive DFC350 digital camera and a 63X/1.30 immersion objective was used for wide field fluorescence microscopy. A stack of 20 z-focal plane images (~0.2 µm depth) was collected for each field. Images were taken from freshly harvested cells and a minimum of 70 cells were quantified per experimental data point. The AF6000 (Leica) and Fiji-ImageJ (NIH) software were used for image processing and quantification. Two-dimensional maximum projections of the focal area in the z-slices were used for analysis.
For viability spot assays, cultures were adjusted to OD 600 = 0.5 and then serially diluted 5-fold in 96-well plates. A 48-pin replica plater was used to spot ~3 µL onto the corresponding plates, which were incubated at 25 °C for 3–4 days before taking photographs.
Data display
Graphpad Prism 10 was used to present data as either bar graphs or box plots. For bar graphs, error bars represent 95% confidence intervals calculated using the Wilson/Brown method. In box plots, the median line represents the median, the box limits represent the 25th and 75th percentiles, the whiskers extend to the 10th and 90th percentiles, and the dots represent outliers.
Reagents
**: **
**: **
**: **
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Doudna JA Charpentier E 20141128 Genome editing. The new frontier of genome engineering with CRISPR-Cas 9.Science 34662130036-80751258096125809610.1126/science.125809625430774 · doi ↗ · pubmed ↗
- 2Chen B Gilbert LA Cimini BA Schnitzbauer J Zhang W Li GW Park J Blackburn EH Weissman JS Qi LS Huang B 20131219 Dynamic imaging of genomic loci in living human cells by an optimized CRISPR/Cas system.Cell 15570092-86741479149110.1016/j.cell.2013.12.00124360272 PMC 3918502 · doi ↗ · pubmed ↗
- 3Memar N Sethi A Luehr S Lambie EJ Conradt B 20221213 In vivo labeling of endogenous genomic loci in C. elegans using CRISPR/d Cas 9.Micro Publ Biol 202210.17912/micropub.biology.00070136606081 PMC 9807462 · doi ↗ · pubmed ↗
- 4Roggenkamp E Giersch RM Schrock MN Turnquist E Halloran M Finnigan GC 201832 Tuning CRISPR-Cas 9 Gene Drives in Saccharomyces cerevisiae.G 3 (Bethesda)83999101810.1534/g 3.117.30055729348295 PMC 5844318 · doi ↗ · pubmed ↗
- 5Lee ME De Loache WC Cervantes B Dueber JE 201551 A Highly Characterized Yeast Toolkit for Modular, Multipart Assembly.ACS Synth Biol 4997598610.1021/sb 500366 v 25871405 · doi ↗ · pubmed ↗
- 6Machín F Torres-Rosell J Jarmuz A Aragón L 2005117 Spindle-independent condensation-mediated segregation of yeast ribosomal DNA in late anaphase.J Cell Biol 16820021-952520921910.1083/jcb.20040808715657393 PMC 2171579 · doi ↗ · pubmed ↗
- 7Balleza E Kim JM Cluzel P 20171120 Systematic characterization of maturation time of fluorescent proteins in living cells.Nat Methods 1511548-7091475110.1038/nmeth.450929320486 PMC 5765880 · doi ↗ · pubmed ↗
- 8Lawrimore J Kolbin D Stanton J Khan M de Larminat SC Lawrimore C Yeh E Bloom K 202157 The r DNA is biomolecular condensate formed by polymer-polymer phase separation and is sequestered in the nucleolus by transcription and R-loops.Nucleic Acids Res 4980305-10484586459810.1093/nar/gkab 22933836082 PMC 8096216 · doi ↗ · pubmed ↗