Computing on actin bundles network
Andrew Adamatzky, Florian Huber, Joerg Schnauss

TL;DR
This paper proposes a novel approach to computing using actin bundles by leveraging excitation wave propagation to implement Boolean logic circuits, aligning with biological gate distributions.
Contribution
It introduces a relaxed model of actin network computing based on excitation waves on actin bundles, enabling circuit implementation with arbitrary electrode arrangements.
Findings
Feasible implementation of Boolean gates on actin networks.
Matching of gate frequency distributions with biological substrates.
Potential for bio-inspired computing architectures.
Abstract
Actin filaments are conductive to ionic currents, mechanical and voltage solitons. These travelling localisations can be utilised in making the actin network executing specific computing circuits. The propagation of localisations on a single actin filament is experimentally unfeasible, therefore we propose a `relaxed' version of the computing on actin networks by considering excitation waves propagating on actin bundles. We show that by using an arbitrary arrangement of electrodes it is possible to implement two-inputs-one-output circuits. Frequencies of the Boolean gates' detection in actin network match an overall distribution of gates discovered in living substrates.
Click any figure to enlarge with its caption.
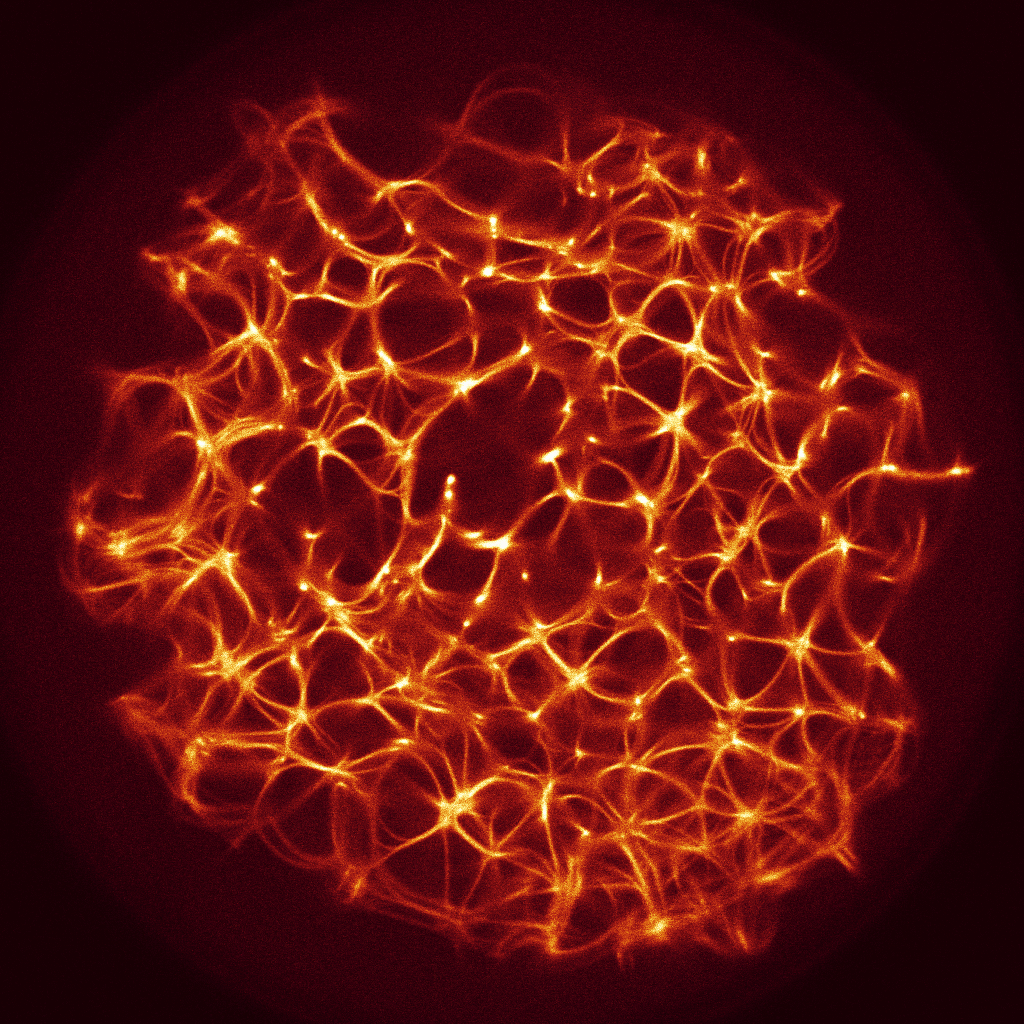 Figure 1
Figure 1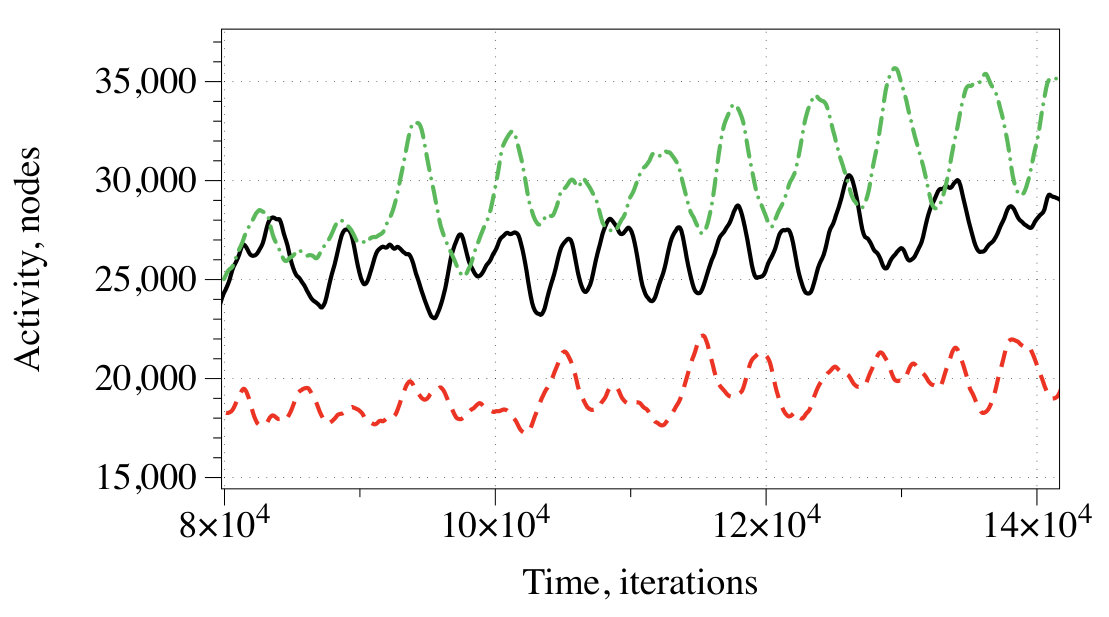 Figure 2
Figure 2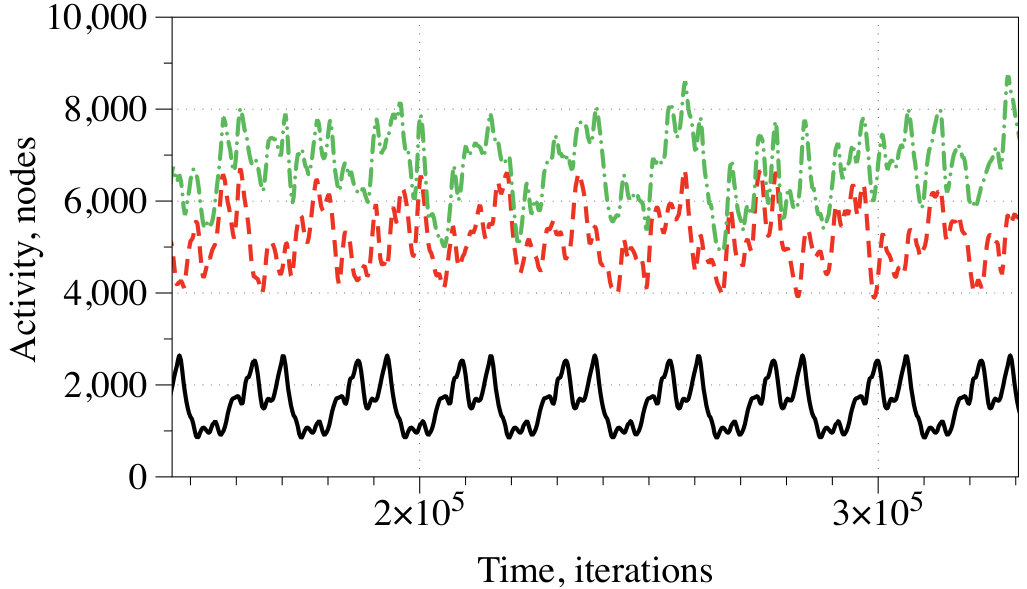 Figure 3
Figure 3 Figure 4
Figure 4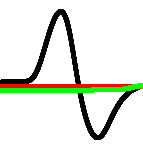 Figure 5
Figure 5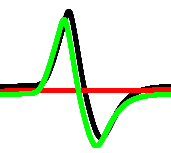 Figure 6
Figure 6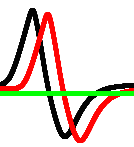 Figure 7
Figure 7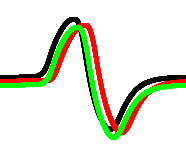 Figure 8
Figure 8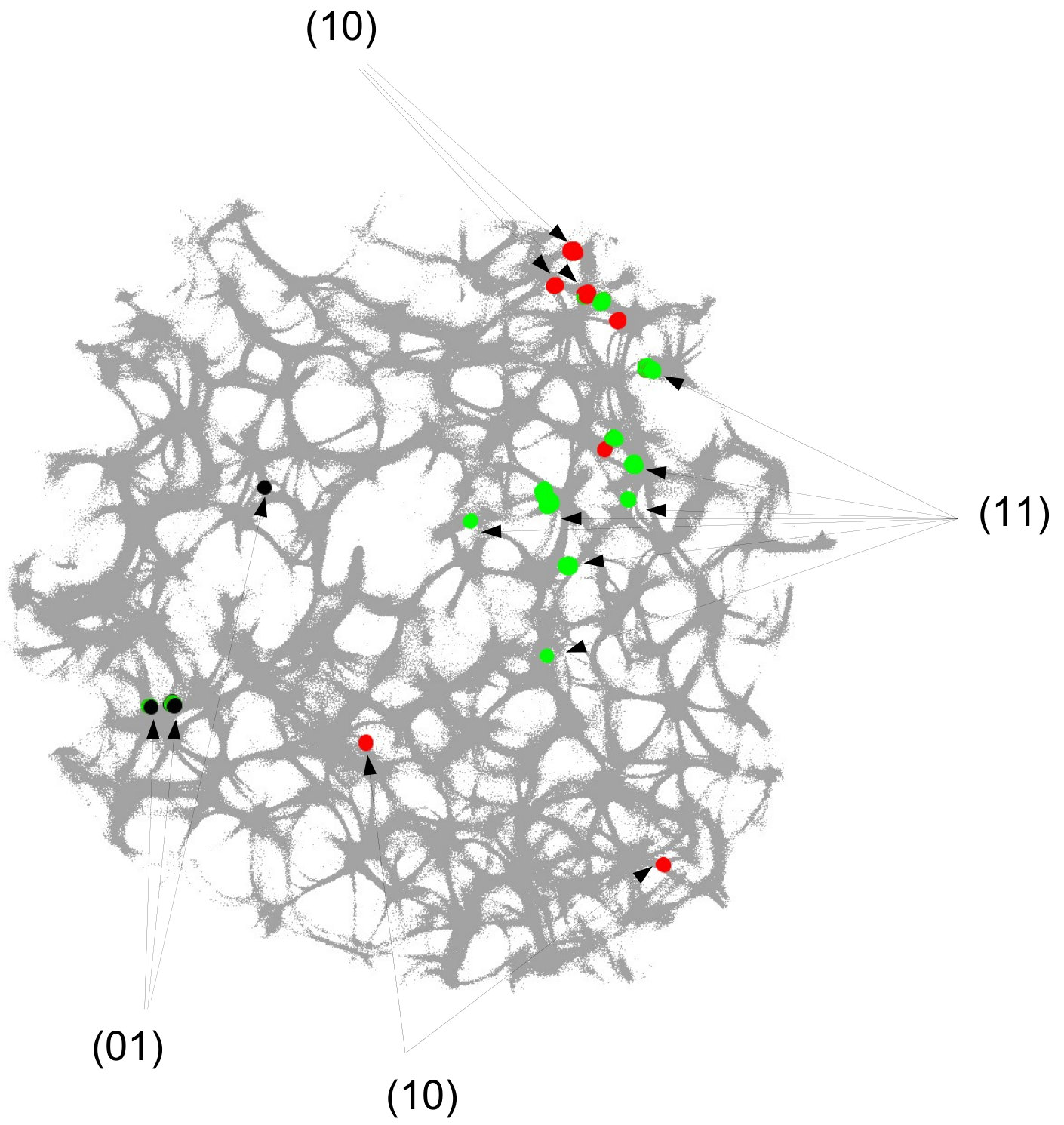 Figure 9
Figure 9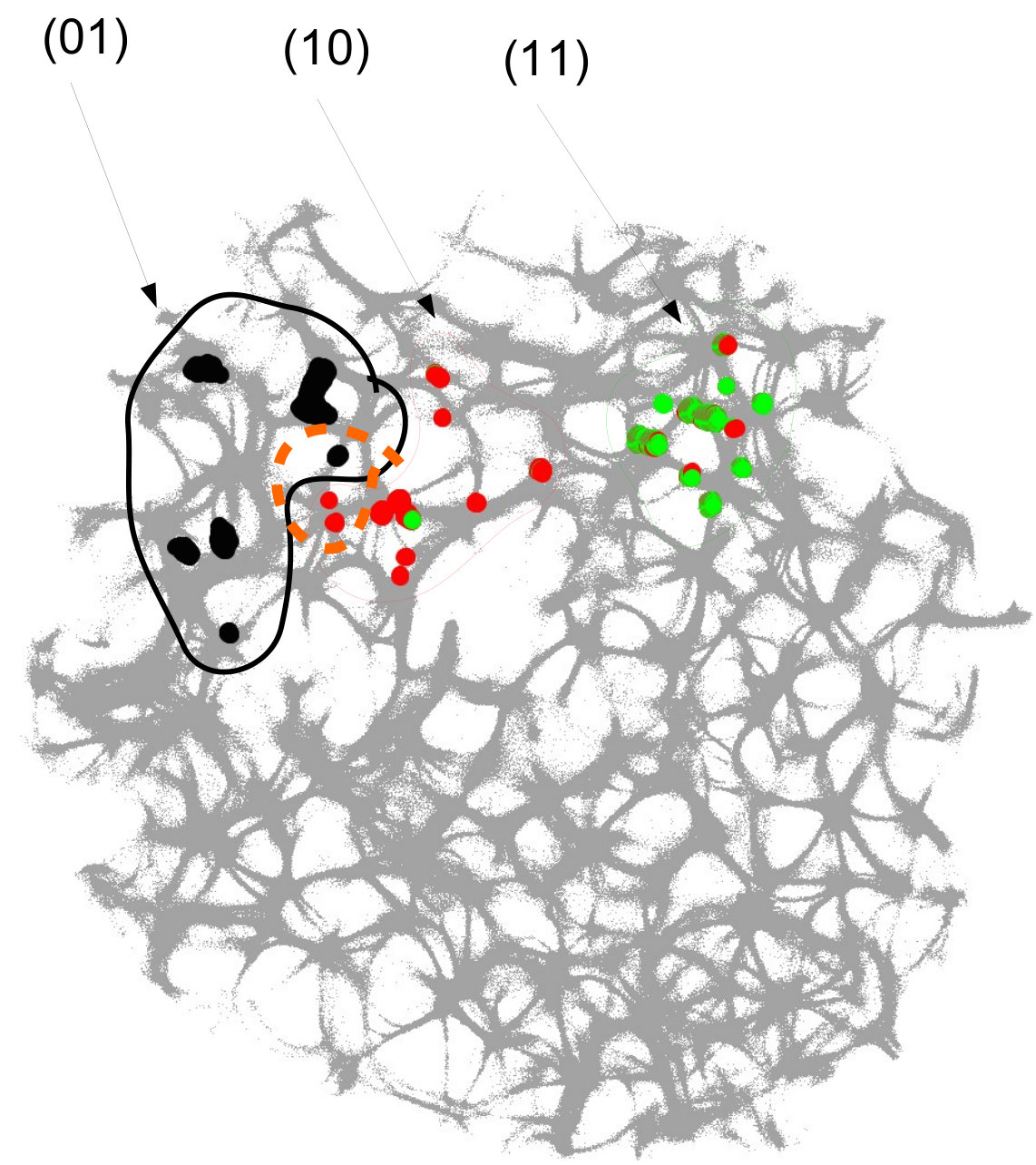 Figure 10
Figure 10 Figure 11
Figure 11 Figure 12
Figure 12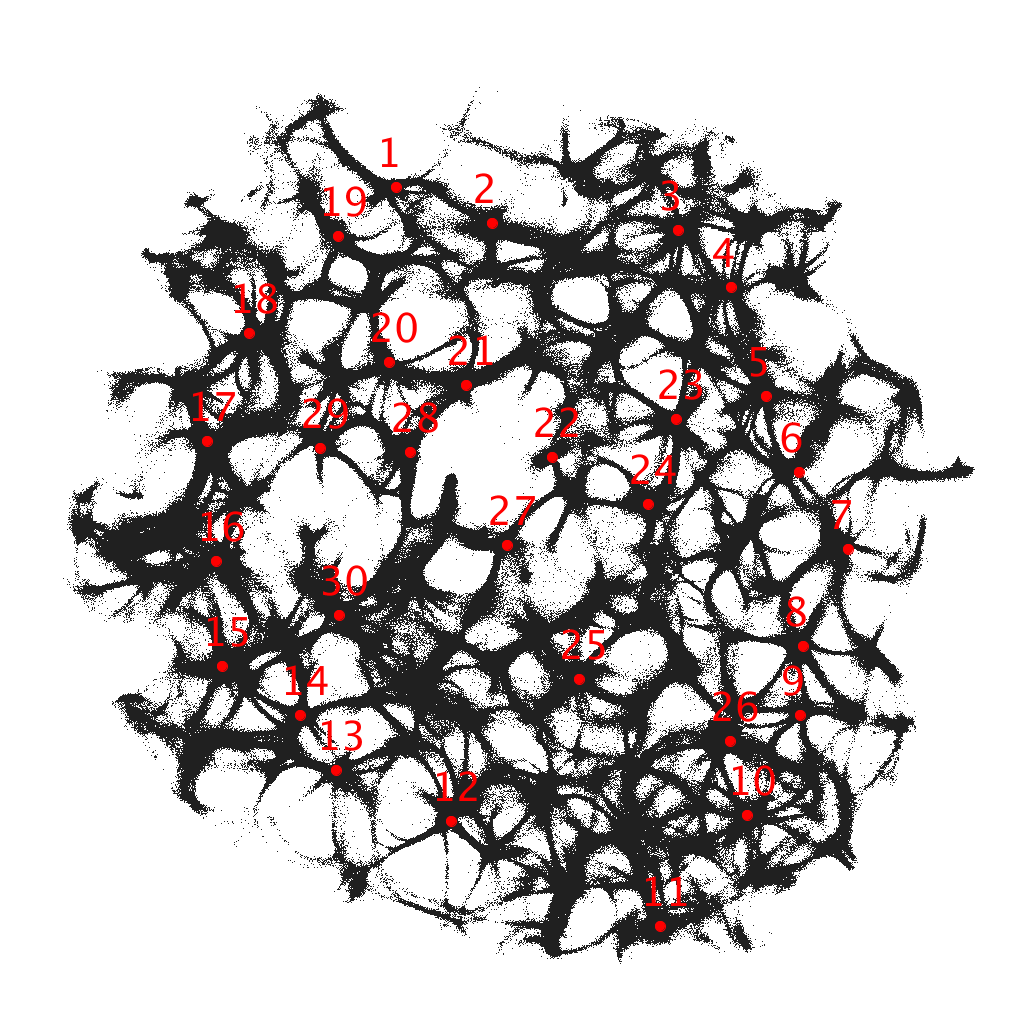 Figure 13
Figure 13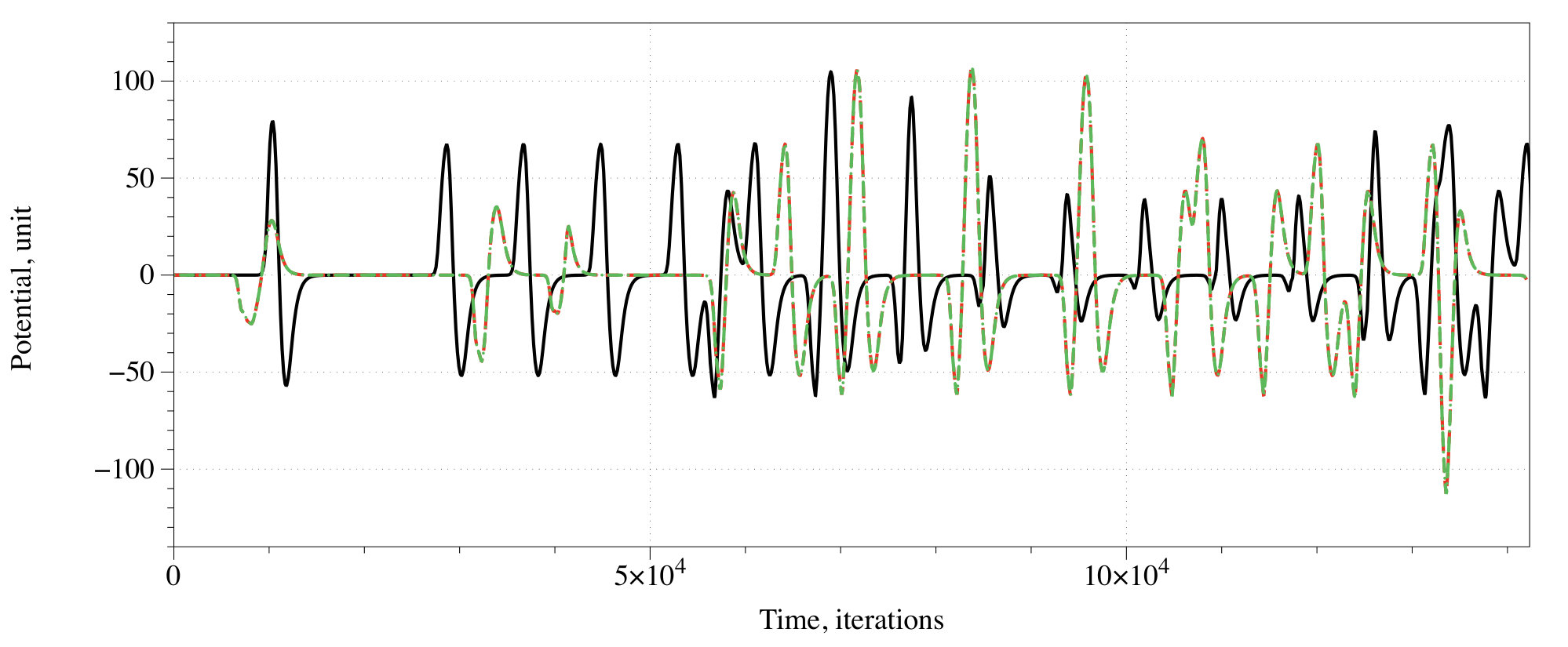 Figure 14
Figure 14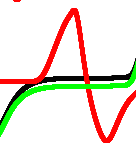 Figure 15
Figure 15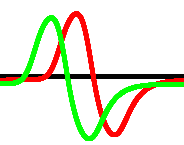 Figure 16
Figure 16 Figure 17
Figure 17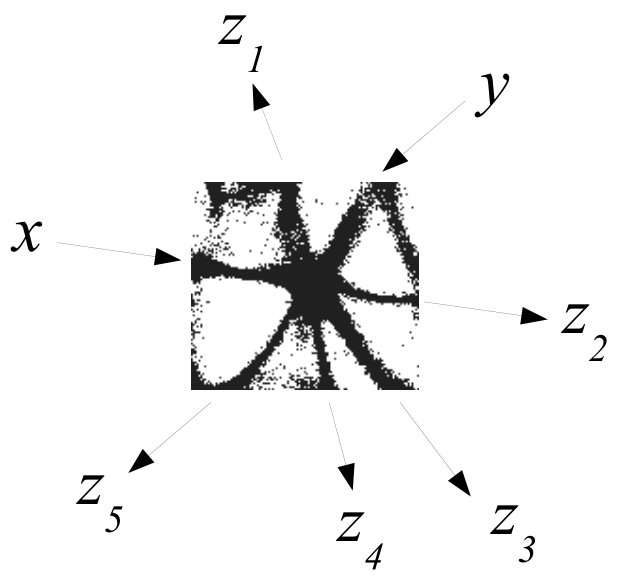 Figure 18
Figure 18 Figure 19
Figure 19 Figure 20
Figure 20 Figure 21
Figure 21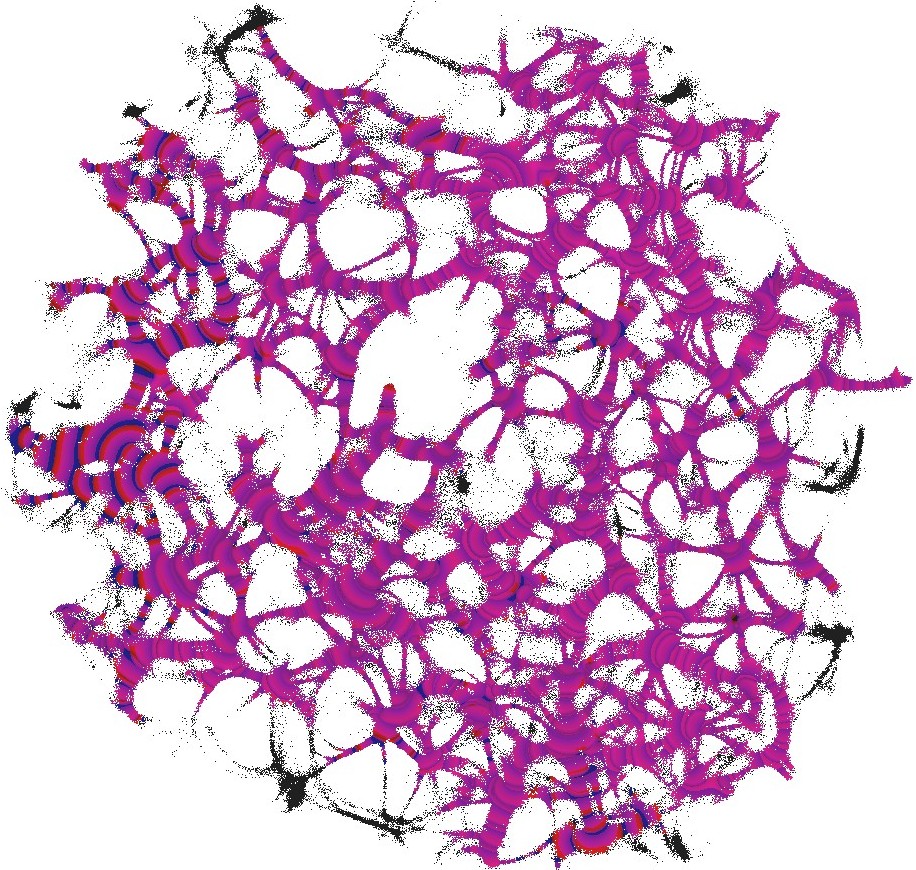 Figure 22
Figure 22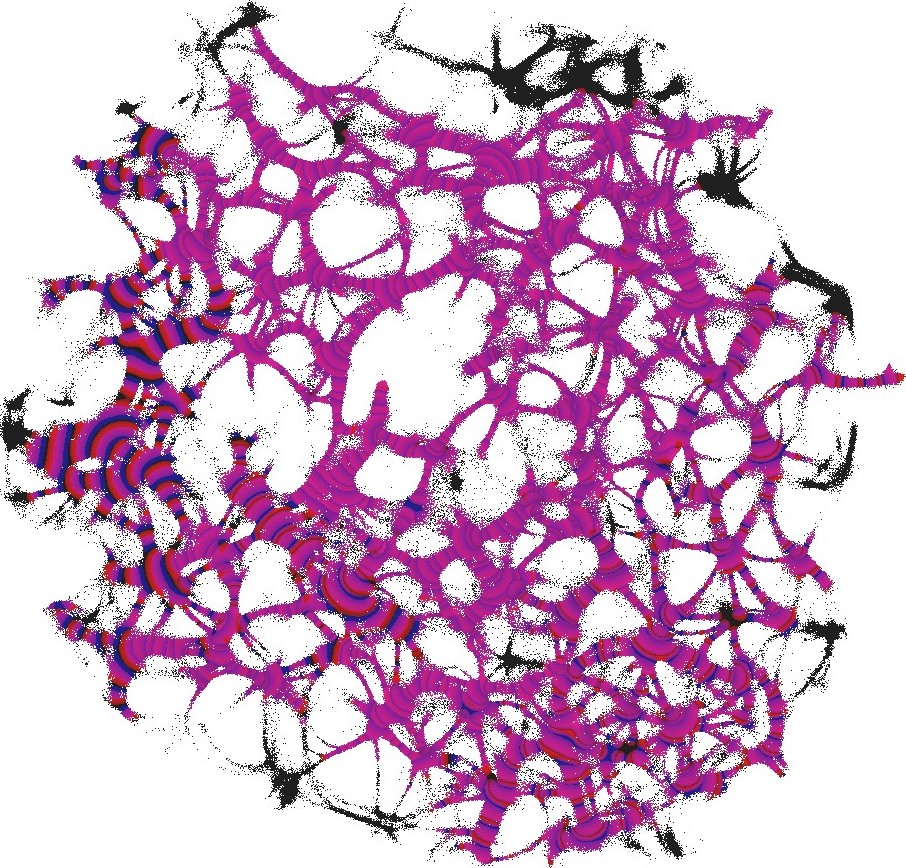 Figure 23
Figure 23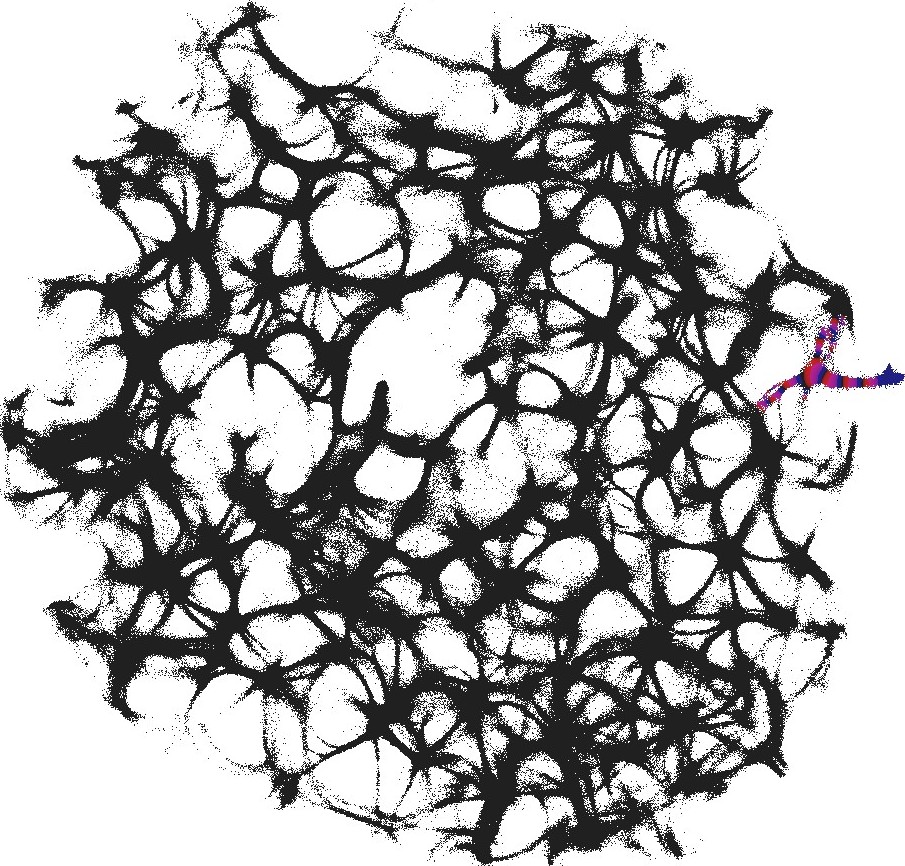 Figure 24
Figure 24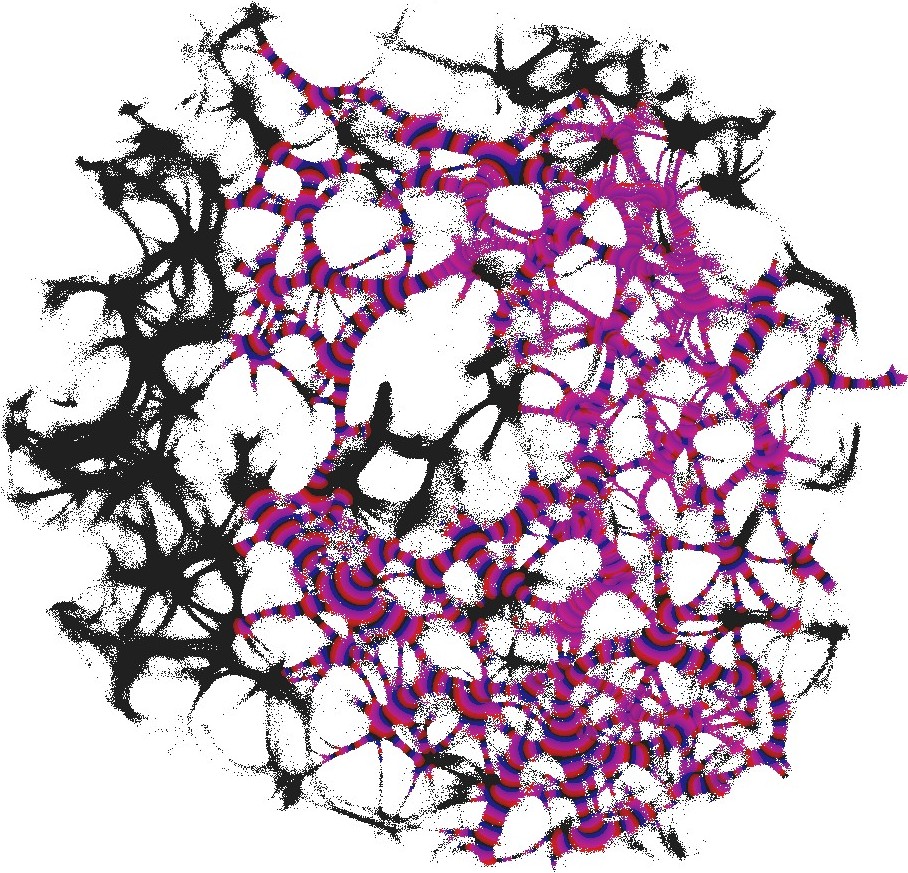 Figure 25
Figure 25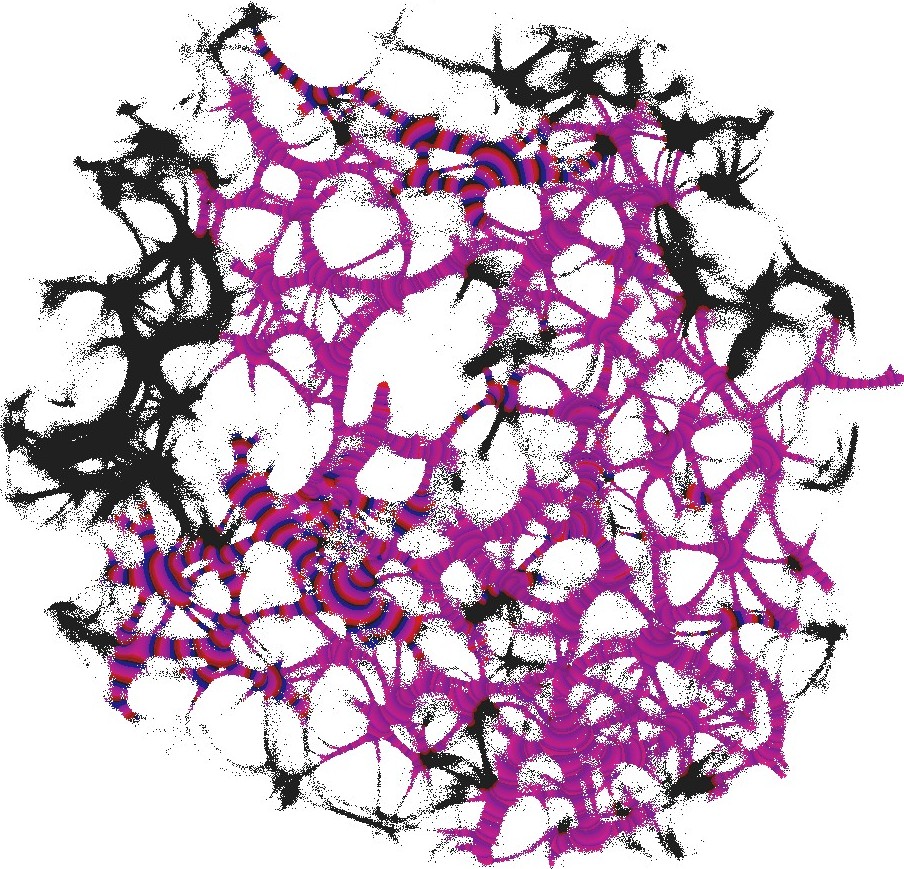 Figure 26
Figure 26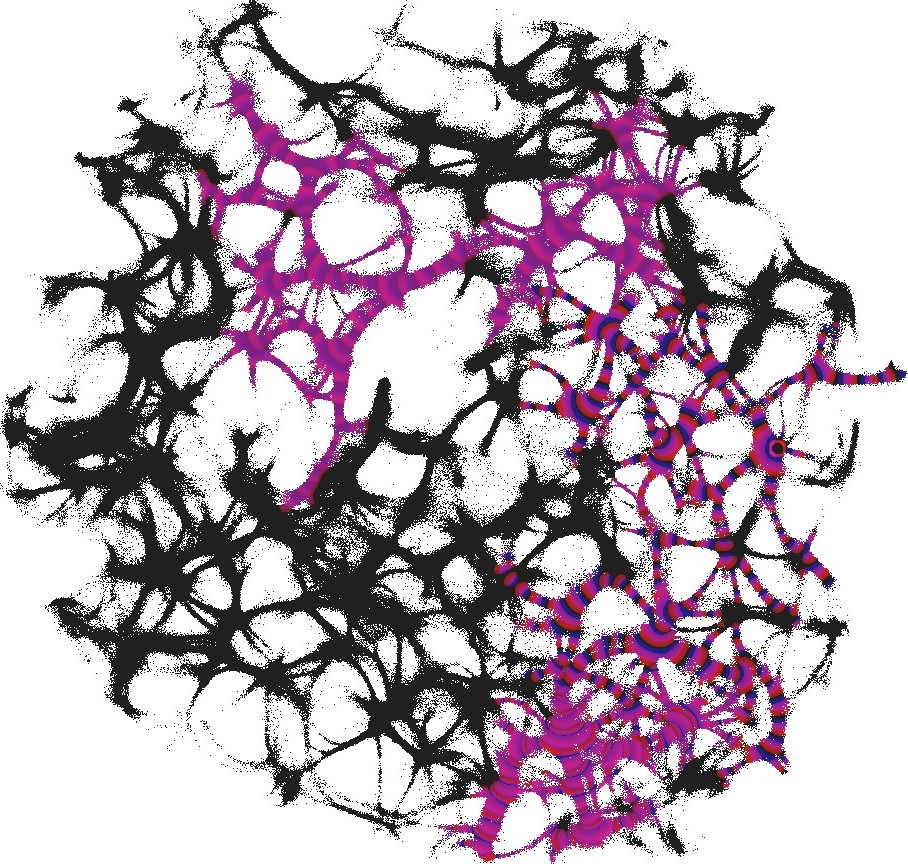 Figure 27
Figure 27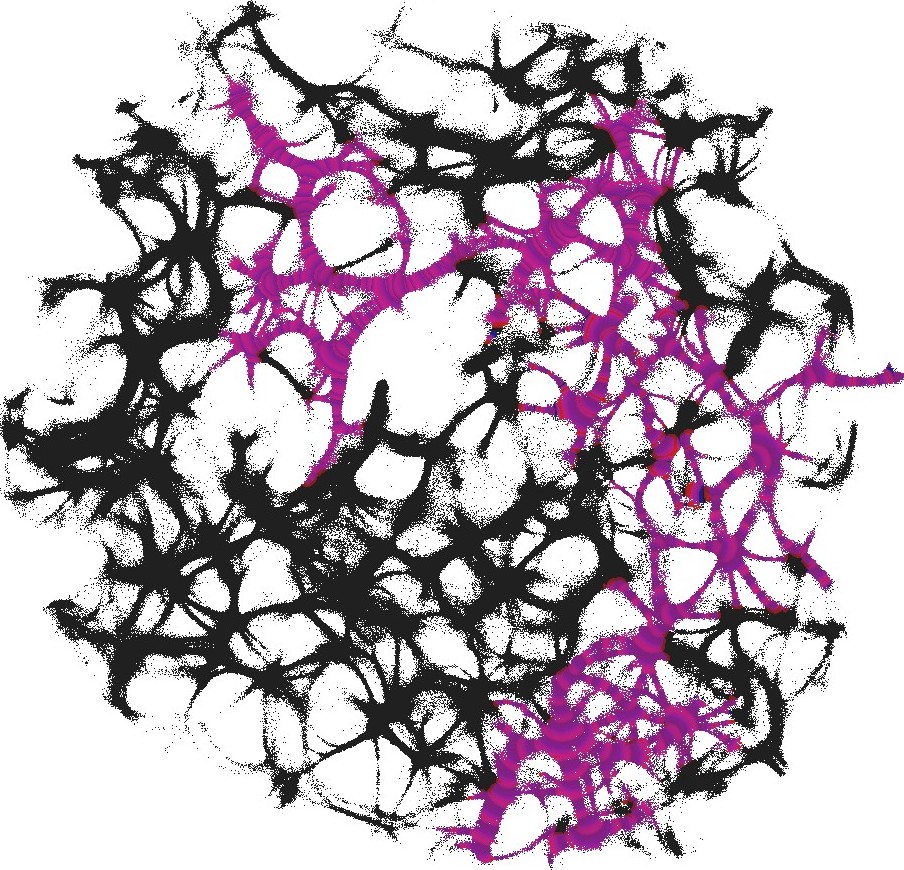 Figure 28
Figure 28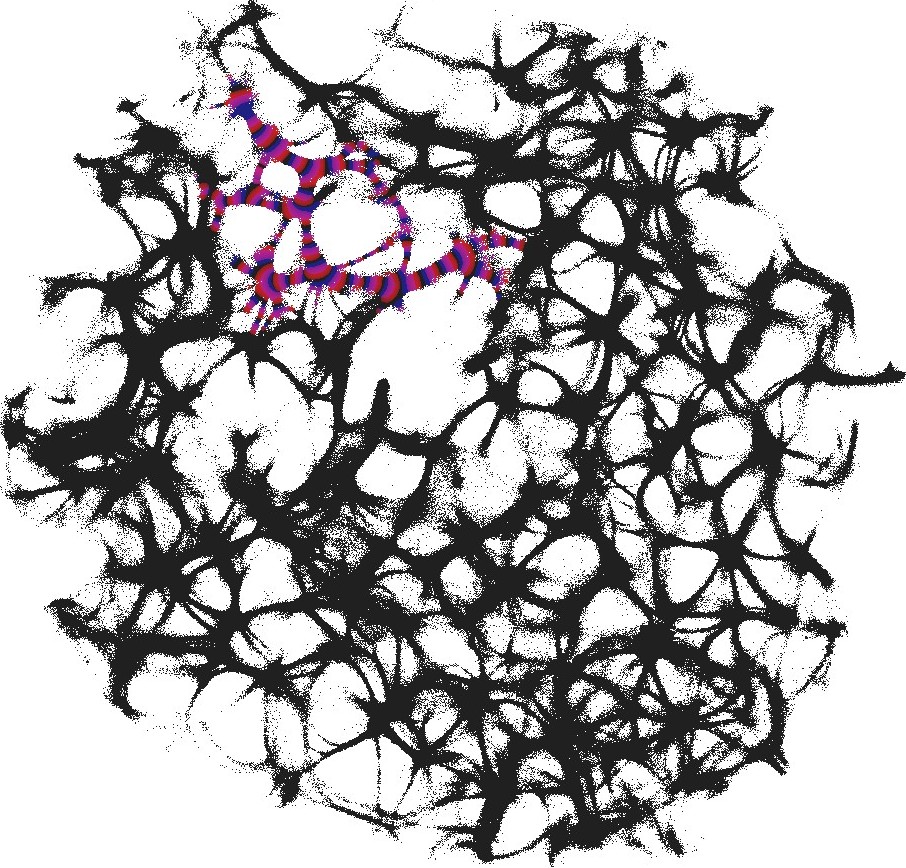 Figure 29
Figure 29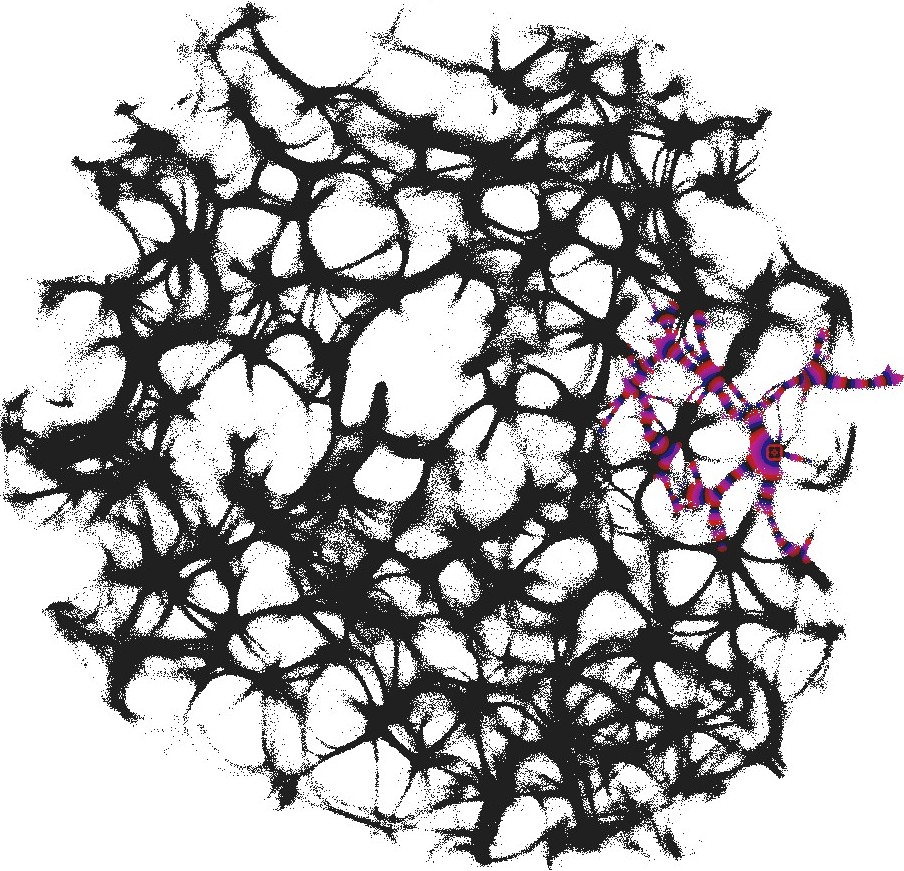 Figure 30
Figure 30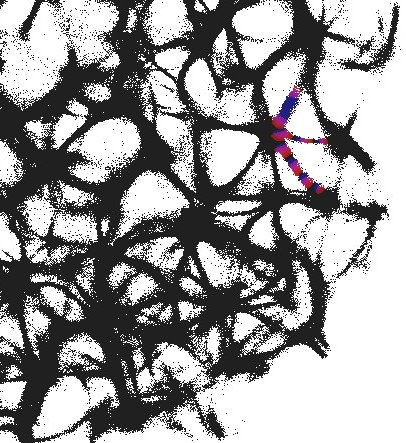 Figure 31
Figure 31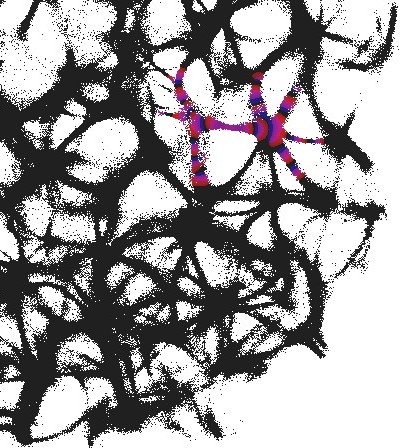 Figure 32
Figure 32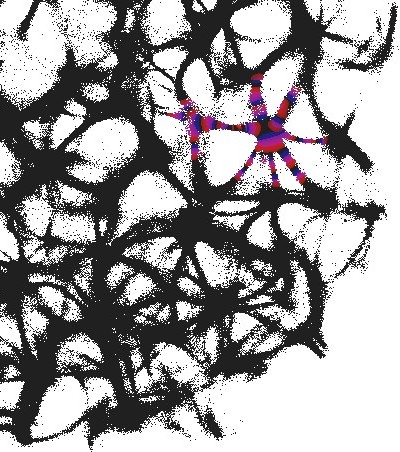 Figure 33
Figure 33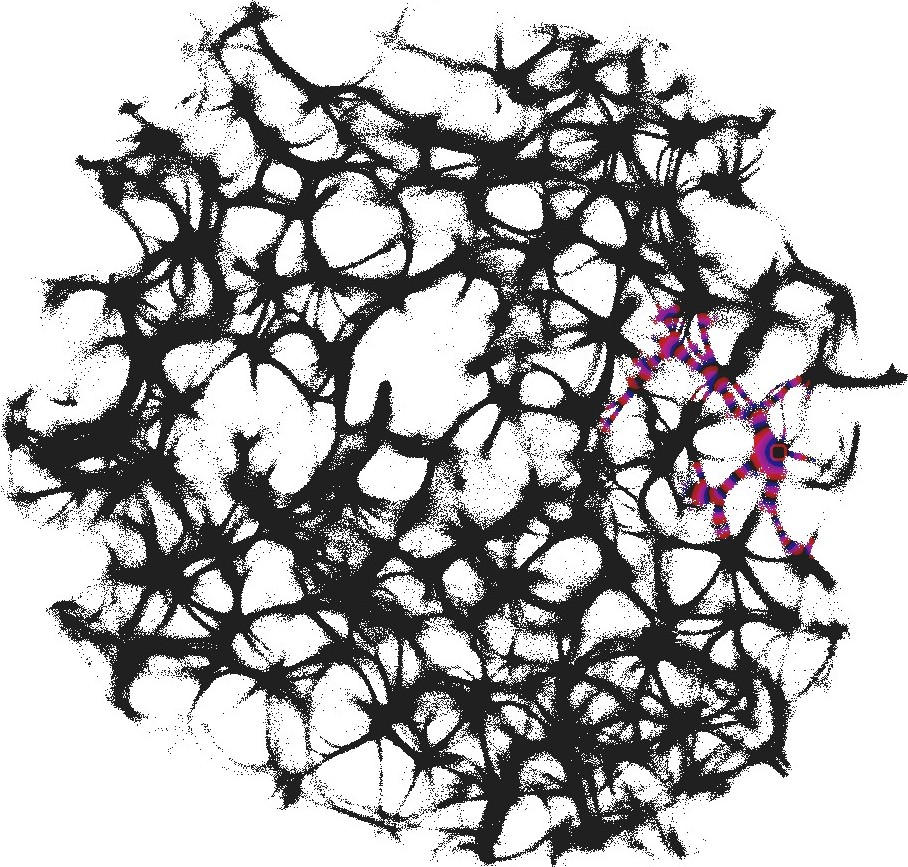 Figure 34
Figure 34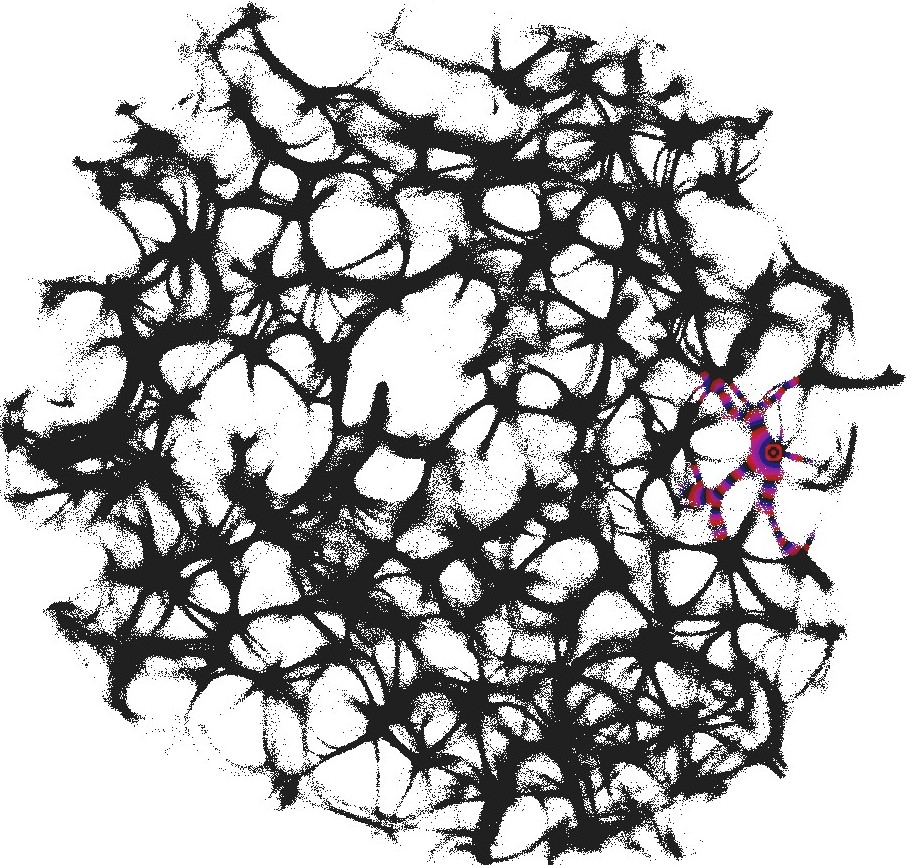 Figure 35
Figure 35| spikes | gate | notations |
|---|---|---|
| or | ||
| select | ||
| xor | ||
| select | ||
| not-and | ||
| and-not | ||
| and |
| () | Total | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 0 | 7 | 1 | 3 | 2 | 5 | 3 | 21 |
| 2 | 0 | 8 | 1 | 3 | 2 | 5 | 3 | 22 |
| 3 | 2 | 2 | 0 | 6 | 5 | 3 | 4 | 22 |
| 4 | 2 | 3 | 0 | 6 | 5 | 3 | 3 | 22 |
| 5 | 2 | 2 | 0 | 6 | 4 | 3 | 4 | 21 |
| 6 | 1 | 4 | 1 | 5 | 4 | 2 | 5 | 22 |
| 7 | 1 | 4 | 1 | 4 | 2 | 2 | 2 | 16 |
| 8 | 0 | 3 | 1 | 6 | 3 | 1 | 2 | 16 |
| 9 | 1 | 2 | 0 | 7 | 3 | 1 | 0 | 14 |
| 10 | 1 | 2 | 0 | 8 | 4 | 0 | 0 | 15 |
| 11 | 1 | 3 | 0 | 4 | 3 | 3 | 3 | 17 |
| 12 | 2 | 7 | 0 | 3 | 0 | 3 | 0 | 15 |
| 13 | 3 | 10 | 0 | 1 | 0 | 3 | 0 | 17 |
| 14 | 2 | 11 | 0 | 1 | 0 | 4 | 0 | 18 |
| 15 | 0 | 11 | 0 | 4 | 0 | 3 | 0 | 18 |
| 16 | 0 | 11 | 0 | 3 | 1 | 4 | 0 | 19 |
| 17 | 2 | 9 | 0 | 2 | 0 | 2 | 0 | 15 |
| 18 | 3 | 8 | 0 | 2 | 0 | 2 | 0 | 15 |
| 19 | 1 | 5 | 0 | 5 | 2 | 2 | 2 | 17 |
| 20 | 2 | 8 | 1 | 1 | 1 | 5 | 3 | 21 |
| 21 | 2 | 7 | 0 | 3 | 1 | 5 | 3 | 21 |
| 22 | 2 | 4 | 0 | 7 | 2 | 0 | 3 | 18 |
| 23 | 3 | 5 | 1 | 6 | 2 | 1 | 4 | 22 |
| 24 | 1 | 5 | 1 | 7 | 2 | 1 | 4 | 21 |
| 25 | 2 | 7 | 0 | 3 | 2 | 3 | 1 | 18 |
| 26 | 2 | 2 | 0 | 6 | 5 | 2 | 1 | 18 |
| 27 | 2 | 6 | 1 | 5 | 2 | 2 | 3 | 21 |
| 28 | 1 | 10 | 0 | 3 | 0 | 5 | 0 | 19 |
| 29 | 0 | 9 | 0 | 4 | 0 | 4 | 0 | 17 |
| 30 | 2 | 9 | 0 | 3 | 0 | 3 | 1 | 18 |
| Average | 1.43 | 6.13 | 0.30 | 4.23 | 1.90 | 2.73 | 1.80 | 18.53 |
| St dev | 0.94 | 3.06 | 0.47 | 1.96 | 1.67 | 1.46 | 1.65 | 2.57 |
| Median | 2.00 | 6.50 | 0.00 | 4.00 | 2.00 | 3.00 | 2.00 | 18.00 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeural dynamics and brain function · Advanced Memory and Neural Computing · Neuroscience and Neural Engineering
Computing on actin bundles network
Andrew Adamatzky
Unconventional Computing Laboratory, Department of Computer Science, University of the West of England, Bristol, UK
Florian Huber
Netherlands eScience Center, Science Park 140, 1098 XG Amsterdam, The Netherlands
Jörg Schnauß
Soft Matter Physics Division, Peter Debye Institute for Soft Matter Physics, Faculty of Physics and Earth Science, Leipzig University, Germany & Fraunhofer Institute for Cell Therapy and Immunology (IZI), DNA Nanodevices Group, Leipzig, Germany
Abstract
Actin filaments are conductive to ionic currents, mechanical and voltage solitons. These travelling localisations can be utilised in making the actin network executing specific computing circuits. The propagation of localisations on a single actin filament is experimentally unfeasible, therefore we propose a ‘relaxed’ version of the computing on actin networks by considering excitation waves propagating on actin bundles. We show that by using an arbitrary arrangement of electrodes it is possible to implement two-inputs-one-output circuits. Frequencies of the Boolean gates’ detection in actin network match an overall distribution of gates discovered in living substrates.
Keywords: actin, computing, waves, logical gates
1 Introduction
An idea of implementation of computation by using collisions of signals travelling along one-dimensional non-linear geometries can be traced back to mid 1960s when Atrubin developed a chain of finite-state machines executing multiplication [7], Fisher designed prime numbers generators in cellular automata [9] and Waksman proposed the eight-state solution for a firing squad synchronisation problem [46]. In 1986, Park, Steiglitz, and Thurston [23] designed a parity filter cellular automata with soliton-like dynamics of localisation. Their design led to a construction of a 1D particle machine, which performs computation by colliding particles in 1D cellular automata, i.e. computing embedded in a bulk media [36]. Being inspired by translating the purely theoretical ideas of collision-based computing [1, 2] into nano-computing at the subcellular level, we consider actin and tubuline networks being ideal candidates for the computing substrates. The idea of subcellular computing on cytoskeleton networks has been firstly announced by Hameroff and Rasmussen in a context of microtubule automata in 1980s [13, 27, 12]. Also, Priel, Tuszynski and Cantiello analysed how information processing could be realised in actin-tubulin networks of neuron dendrites [26]. In the present paper we focus on actin.
Actin is a crucial protein presented in all eukaryotic cells in forms of monomeric, globular actin (G-actin) and filamentous actin (F-actin) [37, 19, 39]. Under the appropriate conditions, G-actin polymerises into filamentous actin forming a double helical structure [16]. Within a filament, actin monomers display slightly changed shapes compared to their free, globular configuration [22]. Signals in actin networks could be represented by travelling localisations. The existence of the travelling localisations — defects, ionic waves, solitons — in cytoskeleton polymer networks is supported by (bio)-physical models [42, 43, 44, 41, 25, 45, 29, 31, 30, 18].
Computational studies demonstrated that it is feasible to consider implementing Boolean gates on a single actin filament [35] and on an intersection of several actin filaments [34] via collisions between solitons and to use a reservoir-computing-like approach to discover functions on a single actin unit [3] and filament [4]. In 2016, for instance, we demonstrated that it is possible to implement logical circuits by linking the protein chains [34]. In such a setup, Boolean values are represented by localisations travelling along the filaments and computation is realised via collisions between localisations at the junctions between the chains. We have shown that and, or and not gates can be implemented in such setups. These gates can be cascaded into hierarchical circuits, as we have shown on an example of nor [34].
The theoretical models developed so far address processing of information on a single actin unit or a chain of few units. Whilst being attractive from a computing point, the model might be difficult to implement in experimental laboratory conditions. Therefore, we developed an alternative version of the computing on actin networks by considering excitation waves propagating on actin bundles. Not a single actin filament is considered but an overall ‘density’ of the conductive material formed by the actin bundles arranged by crowding effects without the need of additional accessory proteins [32, 33]. First results of the innovative approached are presented below.
2 Model
FitzHugh-Nagumo (FHN) equations [10, 21, 24] give us a qualitative approximation of the Hodgkin-Huxley model [8] of electrical activity of living cells:
[TABLE]
where is a value of a trans-membrane potential, a variable accountable for a total slow ionic current, or a recovery variable responsible for a slow negative feedback, a value of an external stimulation current. The current through intra-cellular spaces is approximated by , where is a conductance. Detailed explanations of the ‘mechanics’ of the model are provided in [28], here we shortly repeat some insights. The term governs a passive spread of the current. The terms and describe the ionic currents. The term has two stable fixed points and and one unstable point , where is a threshold of an excitation.
We integrated the system using the Euler method with the five-node Laplace operator, a time step and a grid point spacing , while other parameters were , , , . We controlled excitability of the medium by varying from 0.09 (fully excitable) to 0.013 (non excitable). Boundaries are considered to be impermeable: , where is a vector normal to the boundary.
We used still images of the actin network, produced in laboratory experiments on formation of regularly spaced bundle networks from homogeneous filament solutions [17] as a conductive template. We have chosen this particular because it results from an experimental protocol which reliably produces regularly spaced aster-based networks formed due to cross-linking and bundling mechanisms in the absence of molecular motor-driven processes or other accessory proteins [17]. These structures effectively form very stable and long-living 3d networks, which can be readily imaged with confocal LSM and subsequently displayed as 2d structures. Thus, these networks can form a hardware of future cytoskeleton based computers [6].
The actin network (Fig. 1) was projected onto a nodes grid. Original image , , where and (Fig. 1) was converted to a conductive matrix (Fig. 1) derived from the image as follows: if , and .
The parameter determines excitability of the medium and thus determines a range of the network coverage by excitation waves fronts. This is illustrated in Fig. 2.
To show dynamics of both and , we calculated a potential at an electrode location as . Locations of electrodes are shown in Fig. 1.
The numerical integration code written in Processing was inspired by [14, 24, 28]. Time-lapse snapshots provided in the paper were recorded at every 150th time step, and we display sites with ; videos and figures were produced by saving a frame of the simulation every 100th step of the numerical integration and assembling them in the video with a play rate of 30 fps. Videos are available at https://zenodo.org/record/2561273.
3 Results
Input Boolean values are encoded in excitation as follows. We earmark two sites of the network as dedicated inputs, and , and represent logical True, or ‘1’, as an excitation. If then site corresponding to is excited, if the site is not excited. There are several ways to represent output values: presence/absence of an excitation wave-front at a dedicated output site, patterns of spike activity in the network and frequencies of the activity in dedicated output domains. We present four prototypes of logical: structural gate (exact timing of collisions between excitation wave-fronts is determined geometrically), frequency-based gate (Boolean values of outputs are encoded into frequencies of excitation), integral activity gates (an activity of the whole network is encoded into Boolean values) and spiking gates (where logical are values are represented by spikes or their combinations and search for the gates is done by using many output electrodes scattered in the network).
3.1 Structural gate
An example of an interaction gate is shown in Fig. 3. The gate is a junction of seven actin bundles, we call them ‘channels’ (Fig. 3). We earmark two channels as inputs and , and five other channels as outputs . To represent we excite channel , to represent we excite channel . When only channel is stimulated the excitation wave-fronts propagate into channels and (Fig. 3(a)). When only channel is stimulated, the excitation is recorded in channels , , (Fig. 3(b)). When both channels are excited, and , the excitation propagates into all channels (Fig. 3(c)). Thus, the following functions are implemented on the output channels , , . The channel is a selector function. The channels and realise disjunction. The channels and implement conjuction. An advantage of the interaction gate is that it is cascadable, i.e. many gates can be linked together without decoders or couplers. A disadvantage is that functioning of the gate is determined by exact geometrical structure of the actin bundle network, which might be difficult to control precisely.
3.2 Frequency based gates
For each pair of inputs we calculated a frequency matrix , , where each entity with coordinates show how often a node of was excited. At every iteration of the simulation at every node we updated the frequency as follows: if . When the simulation ends, the frequencies in all nodes were normalised as . For each of we selected domains of higher frequency as having entities . These domains are shown in Fig. 4. This unique mapping allows to implement any two-input-one-output logical gates by placing electrodes in the required unique domains. For example, by placing electrodes in domains which represent outputs for both input pair and input pair (black and red discs in Fig. 4), we can realise a xor gate.
While in excitable mode, , domains corresponding to different input tuples are somewhat dispersed in the network (Fig. 4), the sub-excitable medium, , shows compact and well spatially separated domains (Fig. 4). Moreover, for we even observe a localised domain (shown by orange dashed contour in Fig. 4), where input tuples and are displayed and thus the xor gate is realised.
3.3 Overall level of activity
At every iteration we measured the activity of the network as a number of conductive nodes with . A stimulation of the resting network evokes travelling wave-fronts, which collide with each other and may annihilate or form new wave-fronts in the result of the collisions. The wave-fronts also travel along cycling pathways in the network. Typically, e.g. after iterations for and iterations for the system falls in one of the limit cycle of the overall level of activity with a range of superimposed oscillations (Fig. 5). We found no evidence that shapes of the superimposed spikes in activity reflect the exact combination of inputs, however, an average level of activity definitely does. A correspondence between input tuples and average level of activity , as a percentage of a total number of conductive nodes is the following:
[TABLE]
By selecting an interval of as a logical True we can implement a range of gates. Consider the scenario : for , for , for , . In the scenario , a range of gates, implementable via assigning an activity interval to True, is limited to for and for .
3.4 Spiking gate
A spiking activity of the network shown in Fig. 1, with in a response to stimulation via electrodes and recorded from electrodes is shown in Fig. 6. Assume each spike represents logical True and that spikes being less than iterations closer to each other happen at the same moment. Then a representation of gates by spikes and their combination will be as shown in Tab. 1.
By selecting some specific intervals of recordings we can realise several gates in a single site of recording. In this particular case we assumed that spikes are separated if a difference between their occurrence time is more than iterations. An example is shown in Fig. 7.
To estimate logical richness of the network, we calculated frequencies of logical gates’ discoveries. For each of the recording sites we calculated a number of gates realised during iterations (Tab. 2). In terms of ‘frequency’ of appearance of gates during the simulation, the gates can be arranged in the following hierarchy, from most frequently to least frequent: select {and-not, not-and} and or xor.
The model can realise two-inputs-two-outputs logical gates when we consider values of two recording electrodes at the same specified interval. For example, a one-bit half-adder: one output is and and another output is xor, and Toffoli gate: one output is select and another xor.
4 Discussion
In numerical experiments we demonstrated that logical gates can be implemented in actin bundle networks by various ways of mapping excitation dynamics of the network onto output space. We illustrate the approach with detailed constructions of structural, frequency-based and overall activity based gates. We concluded our study with a comprehensive analysis of spiking gates, where we constructed a frequency of gates hierarchy. The frequency hierarchy of gates discovered select {and-not, not-and} {or, and } xor matches, with some variations, hierarchies of gates discovered in living slime mould [15], living plants [5], Belousov-Zhabotinsky chemical medium [40]. The gate select is dominating because it reflects a reachability of the recording site from the stimulation site: excitation from one electrode reaches a recording site, while an excitation originated from another electrode does not. The gates and-not and not-and represent the scenario when an excitation wave-front propagating from one input site blocks, for instance by its refractory tail, the wave-front propagating from another input site. Gate and symbolises the situation when wave-fronts originated on both input sites must meet up at some point of their travel to traverse areas with lower excitability, for example loci where a narrow channel enter sudden expansion. When excitation wave-fronts from both input sites can reach a recording side without annihilating each other, the site implements gate or. The gate xor reflects the situation when wave-fronts originated at different input sites cancel each other. Modelling results obtained in the paper are encouraging: they show that a computation can be implemented in an actin bundles network by recording excitation dynamics of the network at few arbitrarily selected domains. Thus, we believe, our further tasks will aim at experimental laboratory implementation of the gates. Based on our previous experimental work described in [17], we can extend this approach by specifically biasing the architectural design of these networks. Actin in its natural environment has many accessory proteins such as crosslinkers, which directly impact the properties of the bundle structures [38]. We have recently been able to mimic the properties of these naturally occurring accessory proteins with DNA-based, artificial constructs, which can alter the properties of actin structures in a programmable fashion [20]. The geometrical design of these constructs can be readily tuned by choosing different numbers of binding domains and by altering the underlying DNA template connecting them. These templates can be designed to favour specific binding angles and the number of bundles per junction. They would only act as a molecular precursor for the architecture of the actin system without interfering in the bundle formation and information transport themselves. With the ability to tune the properties of the junctions we gain control over the computing potential. Theoretically, it would be even possible to mix different types of structural proteins to allow parallel processing of information [11].
Author contributions statements
A.A., F.H., J.S. undertook the research and wrote the manuscript.
Competing interests
The authors declare they have no competing interests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1[1] A. Adamatzky. Collision-based computing in biopolymers and their automata models. International Journal of Modern Physics C , 11:1321–1346, 2000.
- 2[2] Andrew Adamatzky, editor. Collision-based Computing . Springer, 2002.
- 3[3] Andrew Adamatzky. Logical gates in actin monomer. Scientific reports , 7(1):11755, 2017.
- 4[4] Andrew Adamatzky. On discovering functions in actin filament automata. ar Xiv preprint ar Xiv:1807.06352 , 2018.
- 5[5] Andrew Adamatzky, Simon Harding, Victor Erokhin, Richard Mayne, Nina Gizzie, Frantisek Baluška, Stefano Mancuso, and Georgios Ch Sirakoulis. Computers from plants we never made: Speculations. In Inspired by nature , pages 357–387. Springer, 2018.
- 6[6] Andrew Adamatzky, Jack Tuszynski, Joerg Pieper, Dan V Nicolau, Rossalia Rinalndi, Georgios Sirakoulis, Victor Erokhin, Joerg Schnauss, and David M Smith. Towards cytoskeleton computers. A proposal. In Andrew Adamatzky, Selim Akl, and Georgios Sirakoulis, editors, From parallel to emergent computing . CRC Group/Taylor & Francis, 2019.
- 7[7] AJ Atrubin. A one-dimensional real-time iterative multiplier. IEEE Transactions on Electronic Computers , 3:394–399, 1965.
- 8[8] Go W Beeler and H Reuter. Reconstruction of the action potential of ventricular myocardial fibres. The Journal of physiology , 268(1):177–210, 1977.