Impact of the purposeful kinesis on running waves
Nurdan \c{C}abuko\u{g}lu

TL;DR
This paper investigates how purposeful kinesis influences the behavior of population running waves, demonstrating through numerical experiments that kinesis can prevent extinction and affect wave amplitude and predator kinesis.
Contribution
It provides a numerical analysis of purposeful kinesis effects on running waves, including monotonic and Allee effect dependencies, highlighting benefits for population survival.
Findings
Kinesis can prevent extinction despite higher diffusion.
Wave amplitude decreases with prey kinesis reduction.
Predator kinesis reduction causes sharp amplitude increase.
Abstract
The basic model of purposeful kinesis was developed recently (Gorban and Cabukoglu, Ecol. Complex. 2018, 33, 75 83) on the basis of the let well enough alone idea. According to this model the diffusion drops while the reproduction coefficient is increasing. That is, species prefer to stay in a good condition and the population gives birth; otherwise, in the bad situation individuals want to run away because of the fatal conditions. In this study, we analyse the impact of the purposeful kinesis model on running waves. The running waves in the population with kinesis are studied using numerical experiments. Both monotonic and non monotonic (Allee effect) dependence of the reproduction coefficient on the population density are studied. The possible benefits of the purposeful kinesis are demonstrated: with the higher diffusion, while the population without kinesis ends up with extinction,…
Click any figure to enlarge with its caption.
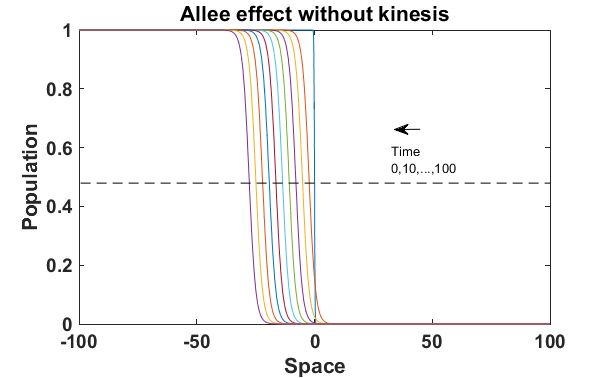 Figure 2
Figure 2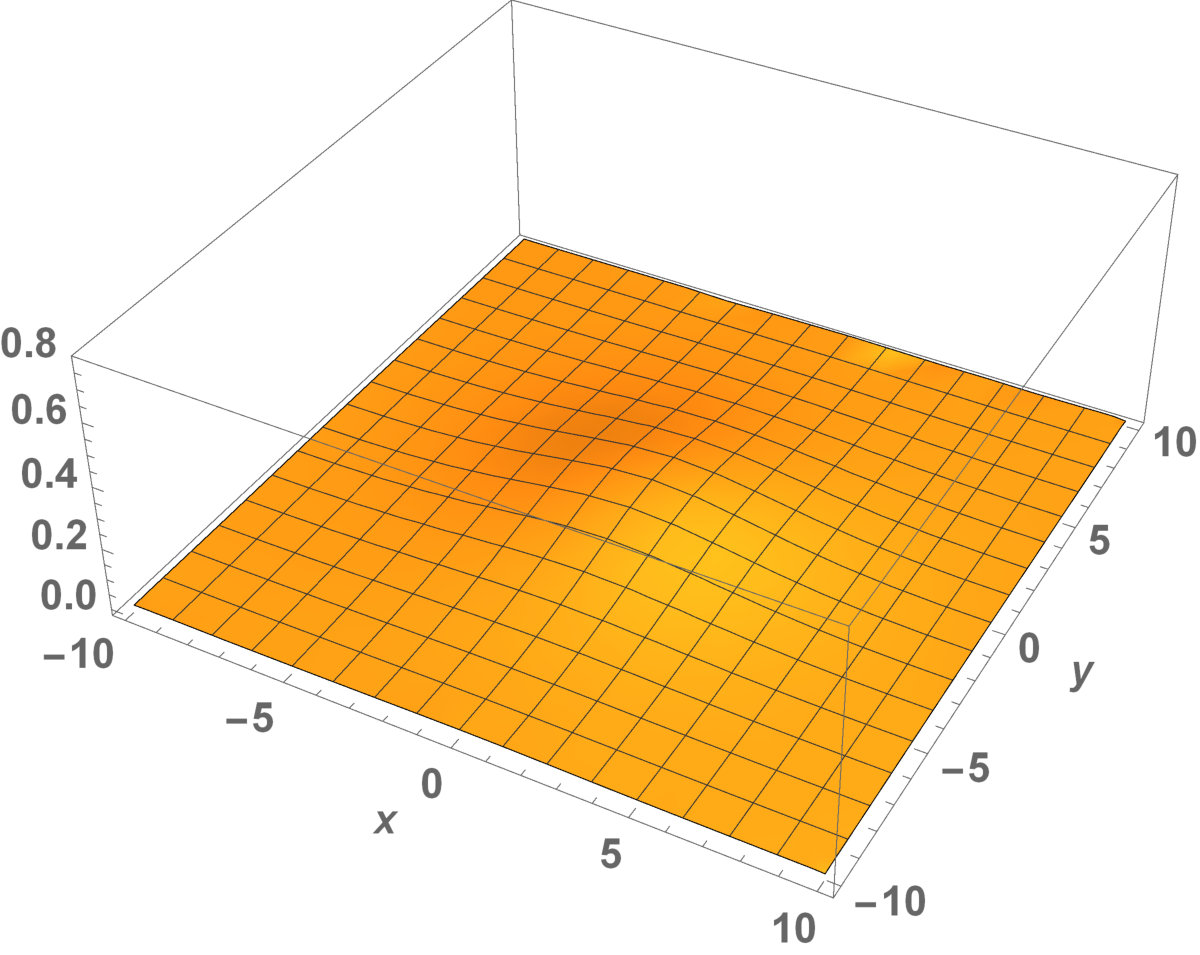 Figure 2
Figure 2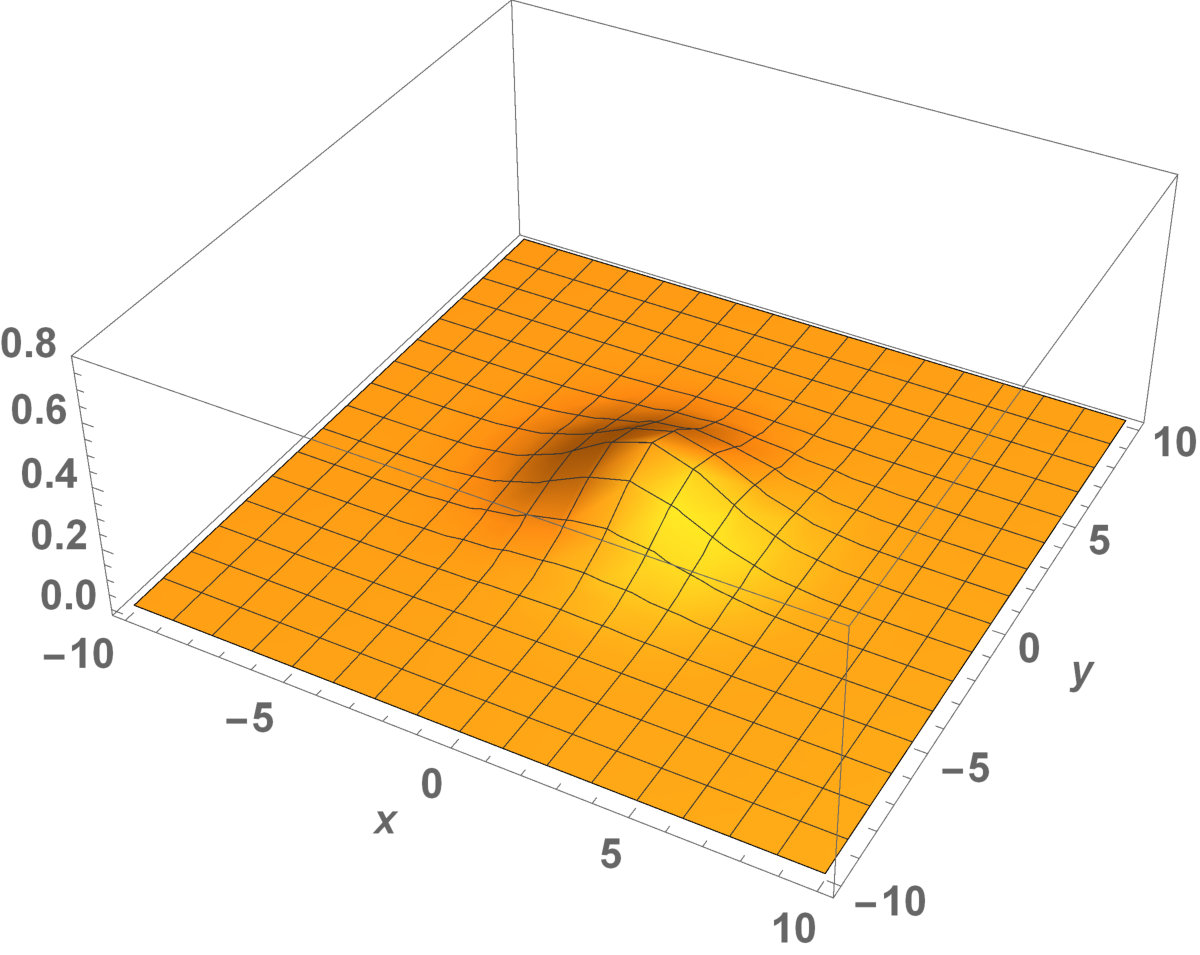 Figure 3
Figure 3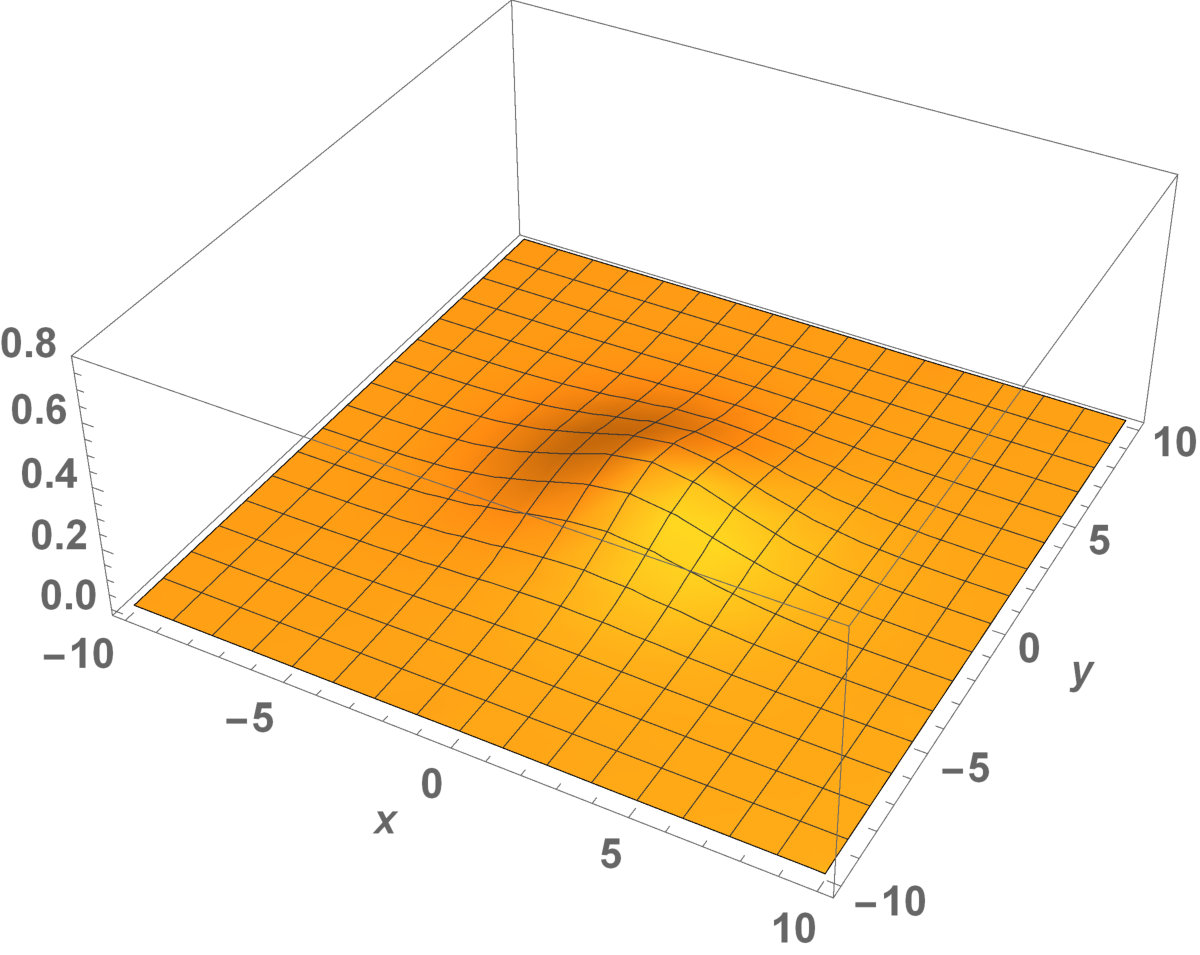 Figure 4
Figure 4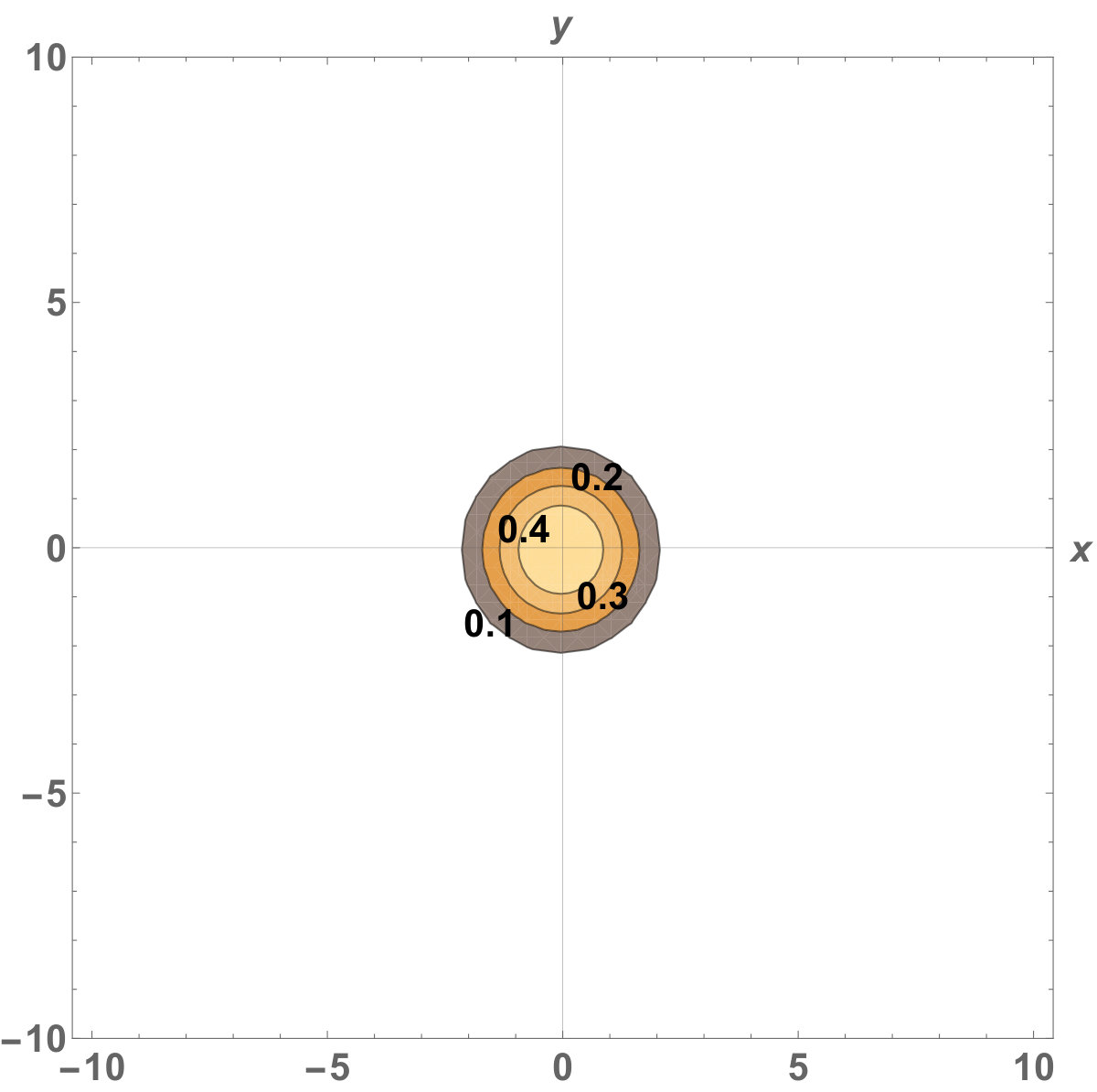 Figure 5
Figure 5 Figure 6
Figure 6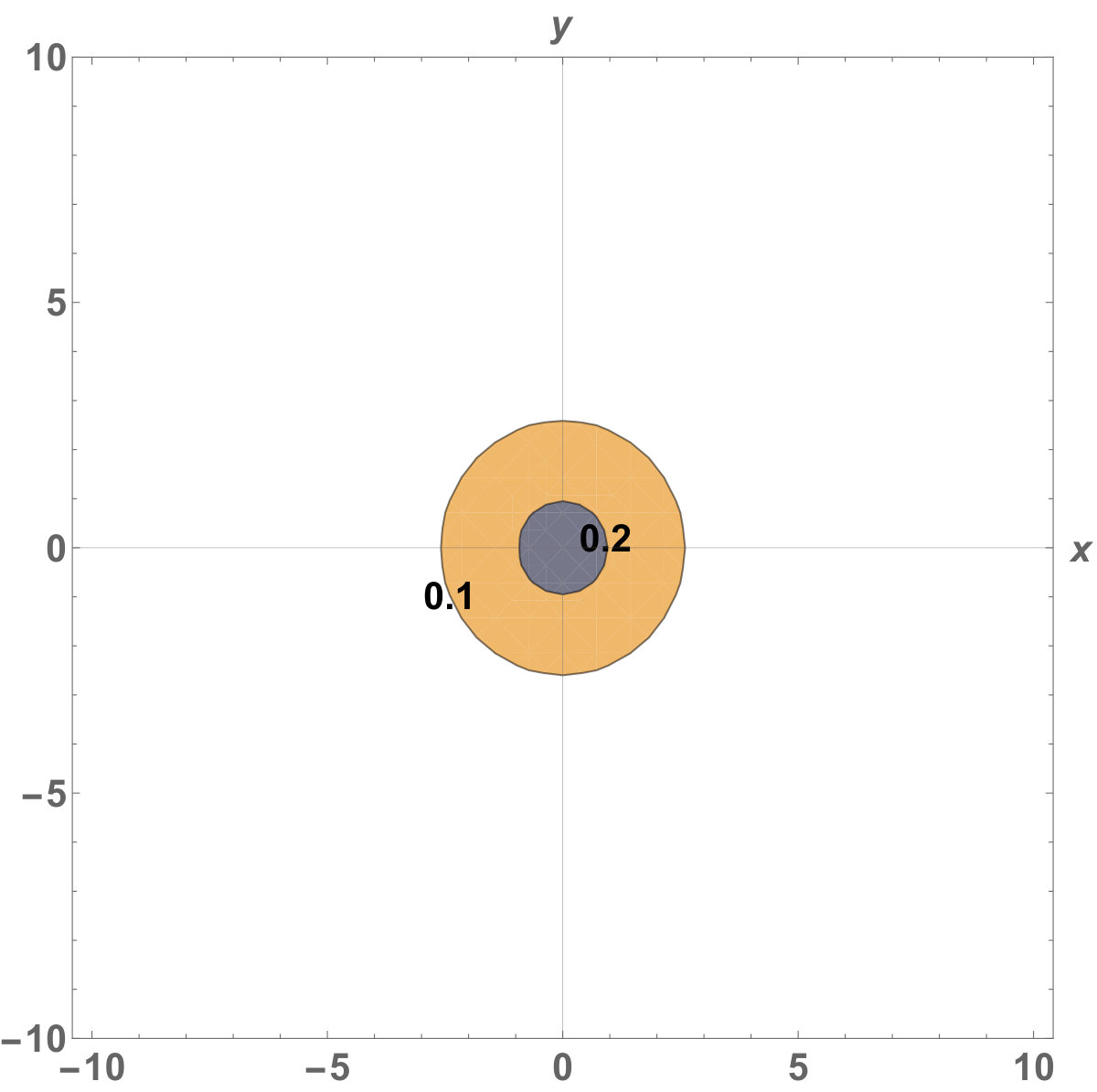 Figure 7
Figure 7 Figure 8
Figure 8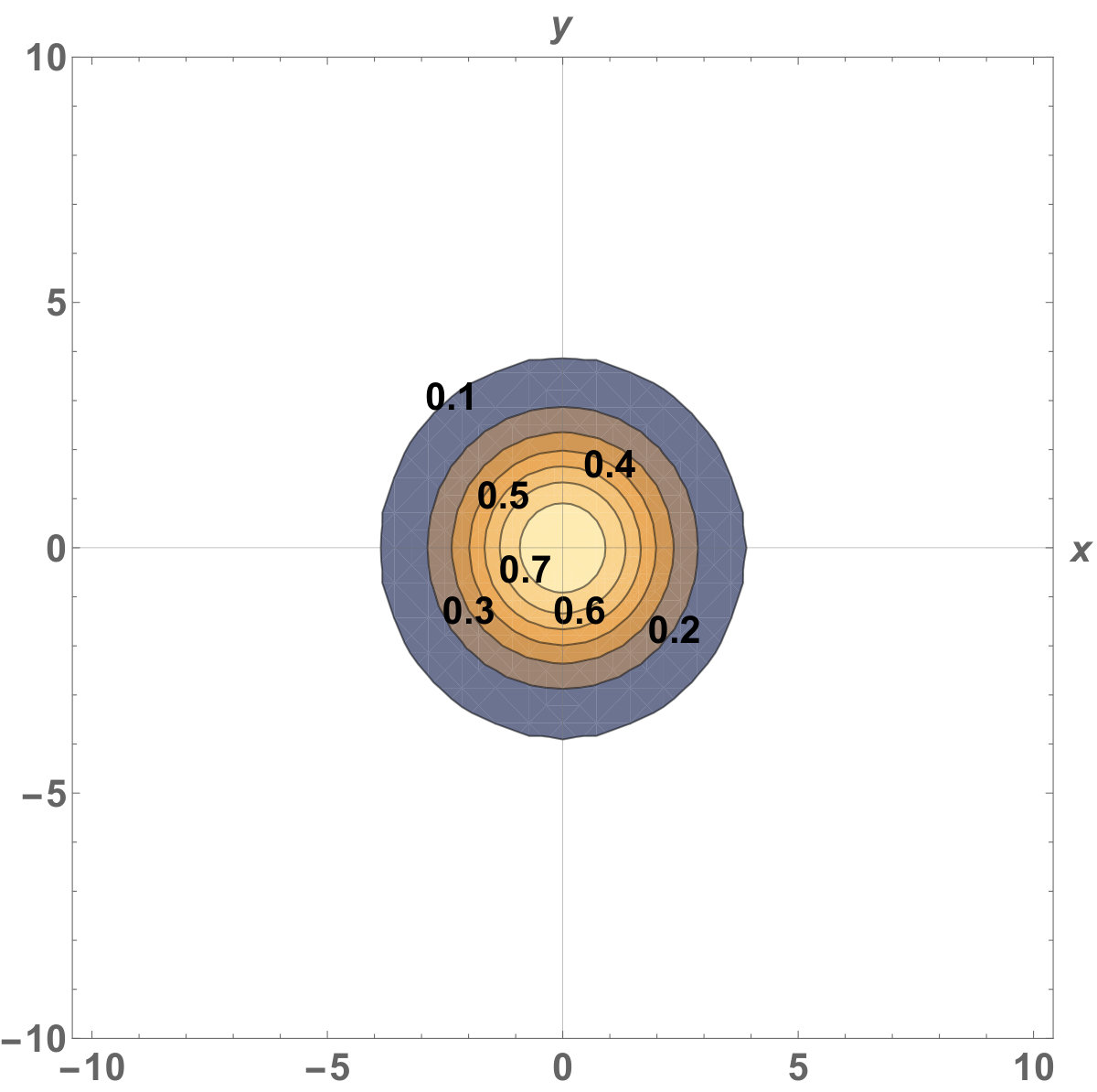 Figure 9
Figure 9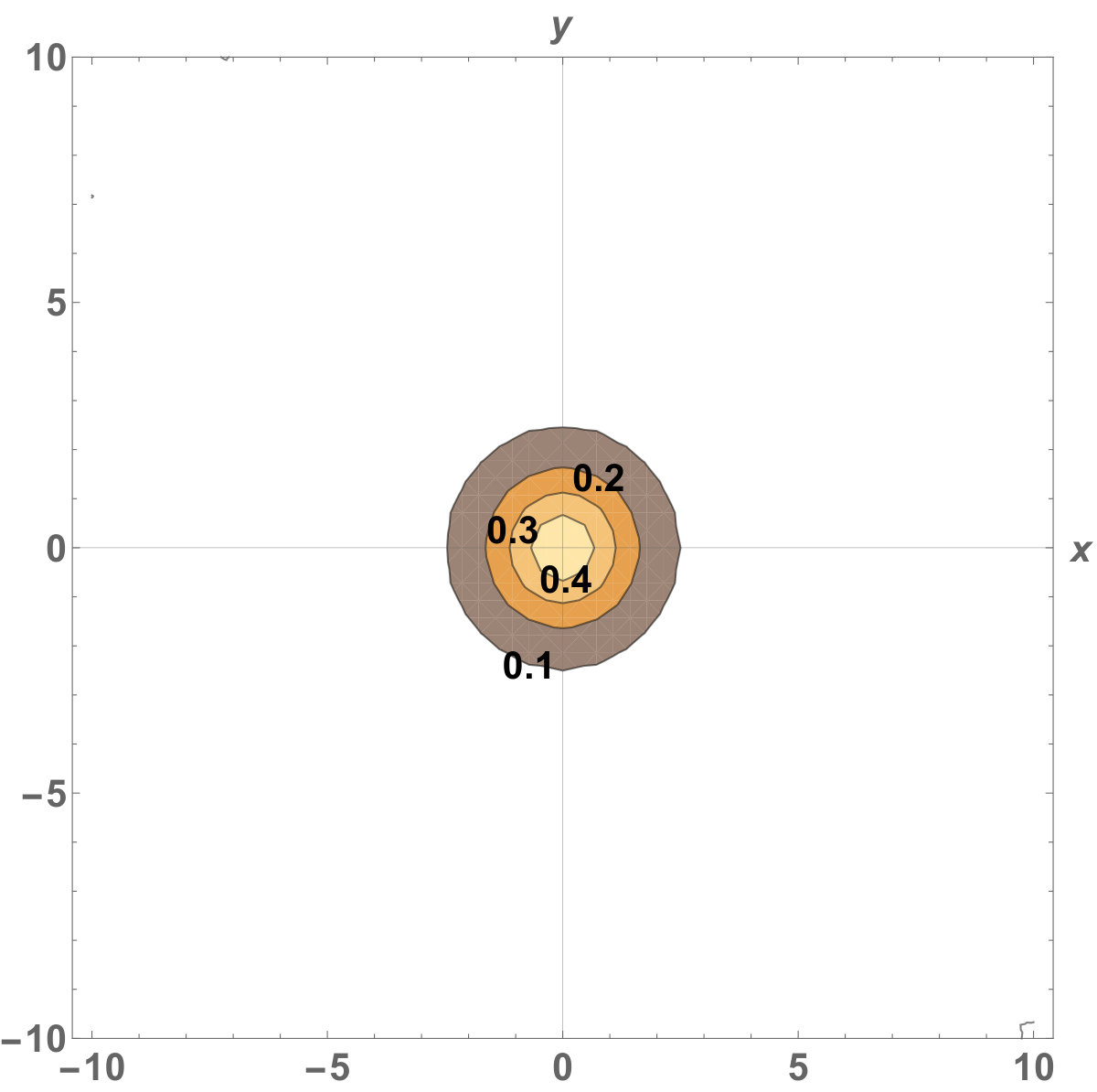 Figure 10
Figure 10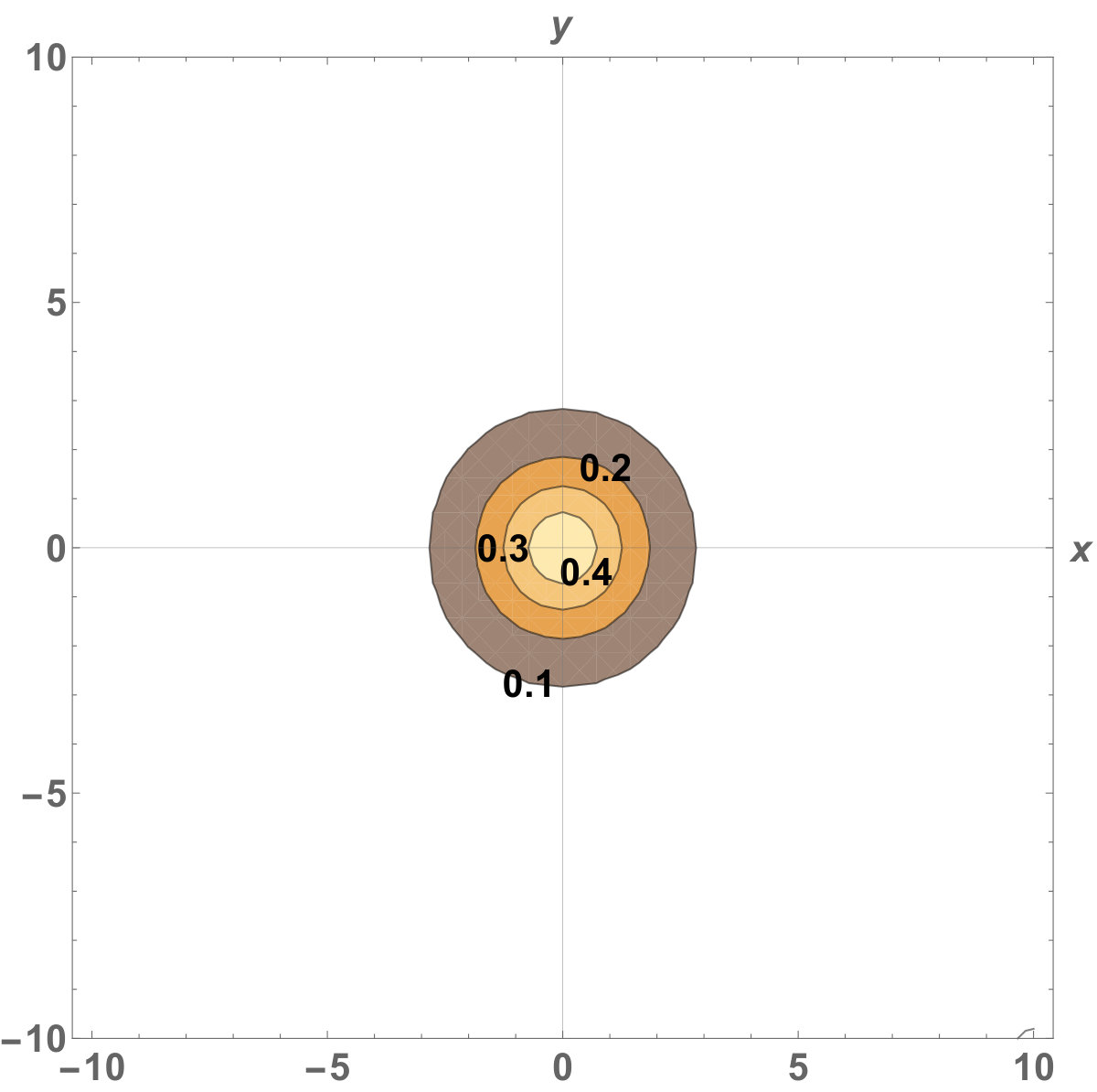 Figure 11
Figure 11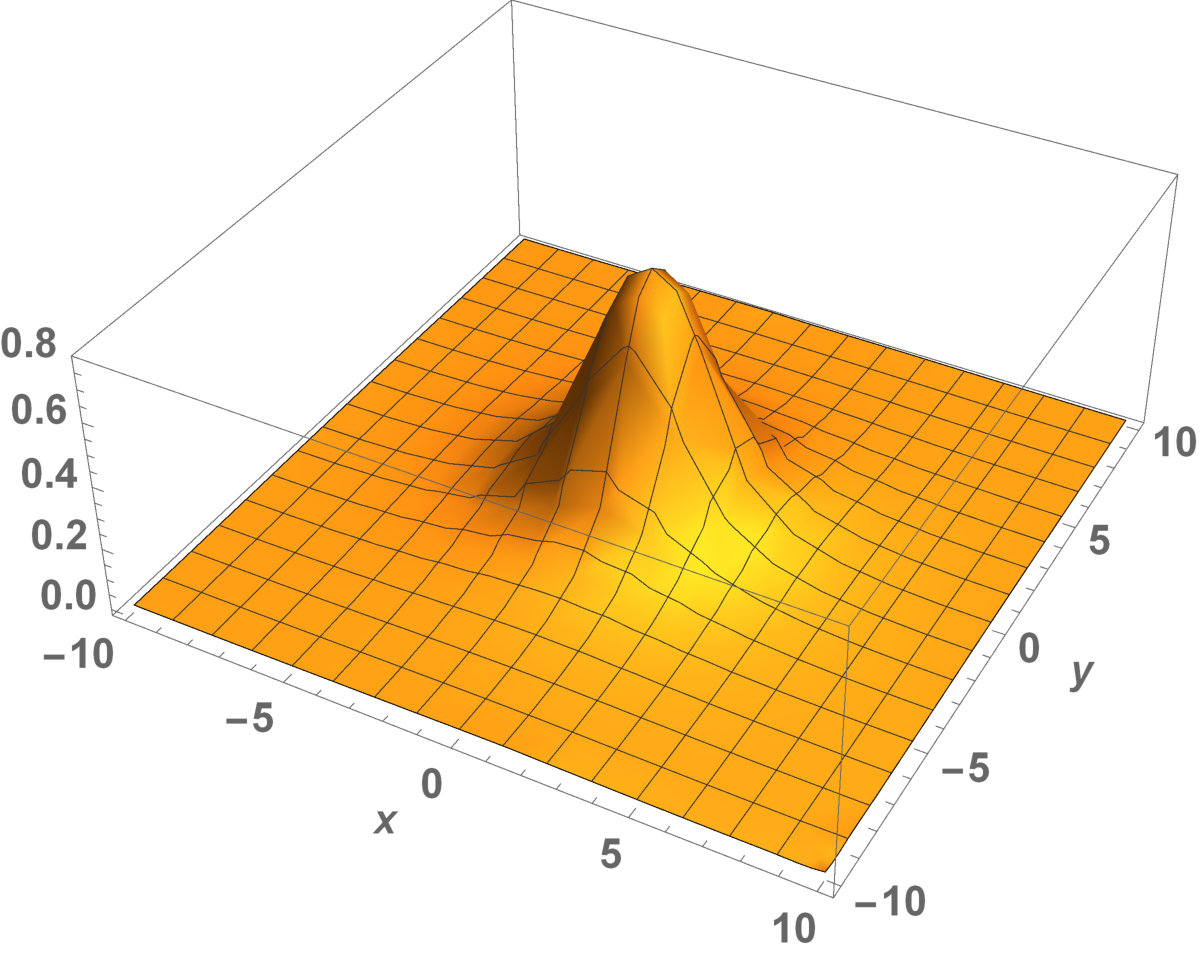 Figure 12
Figure 12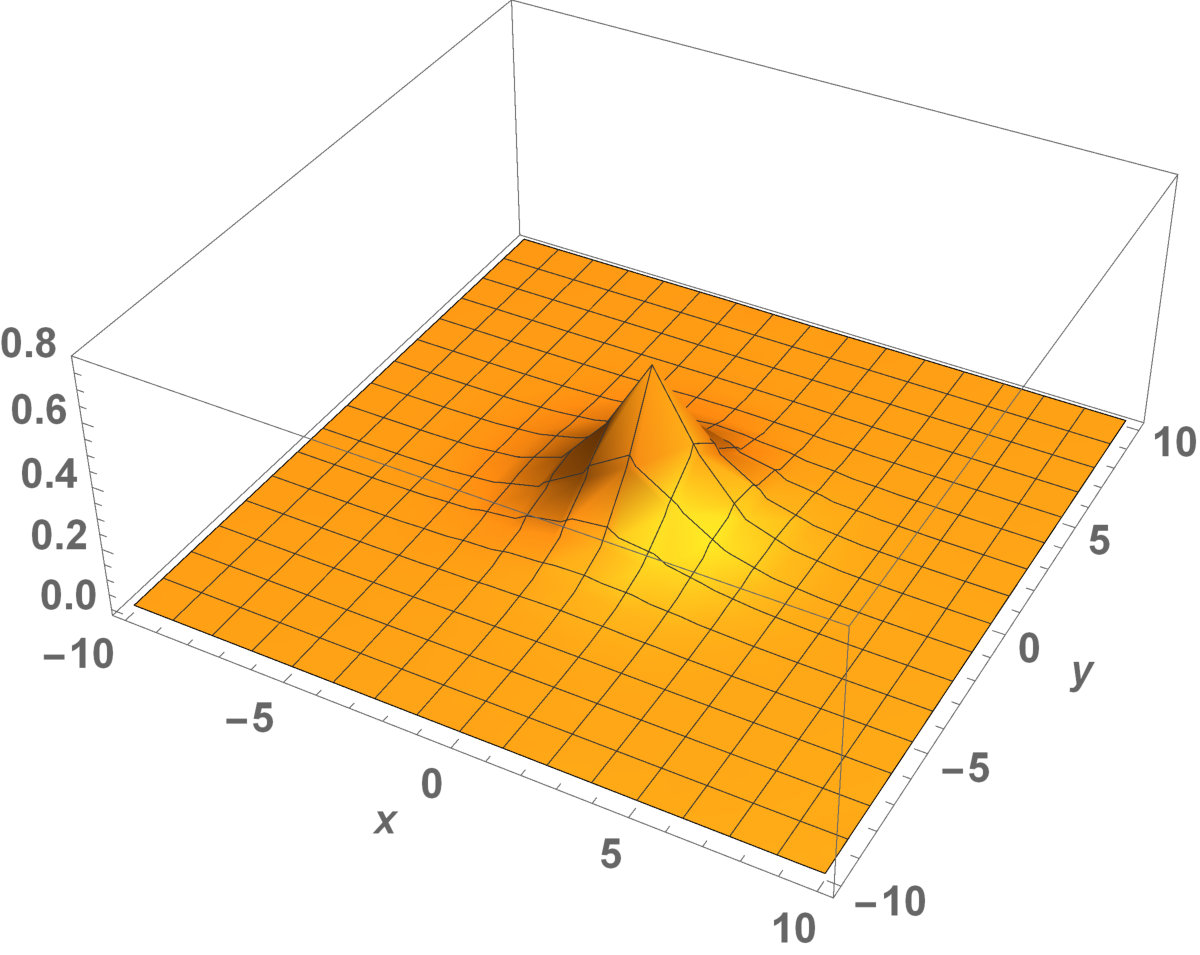 Figure 13
Figure 13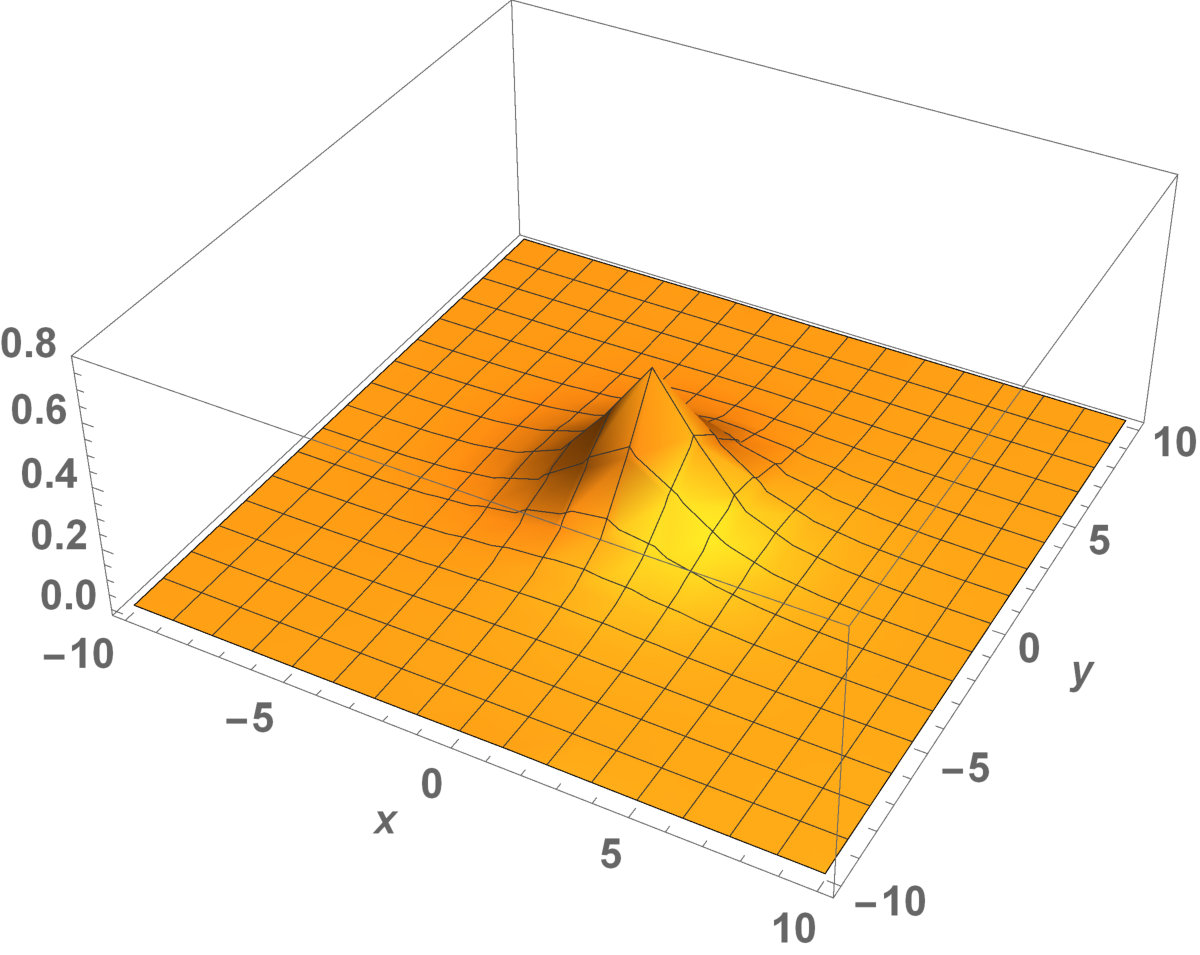 Figure 14
Figure 14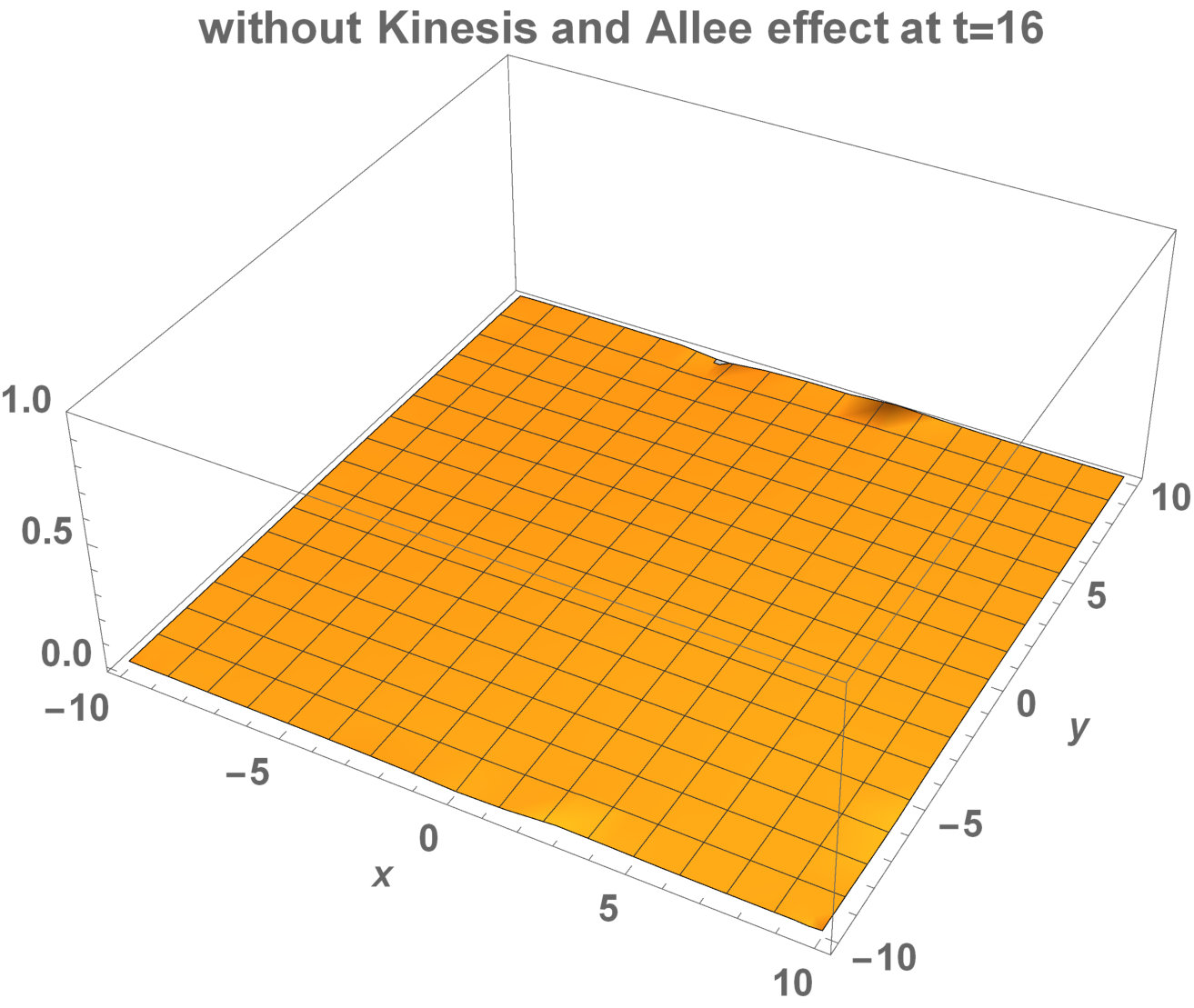 Figure 15
Figure 15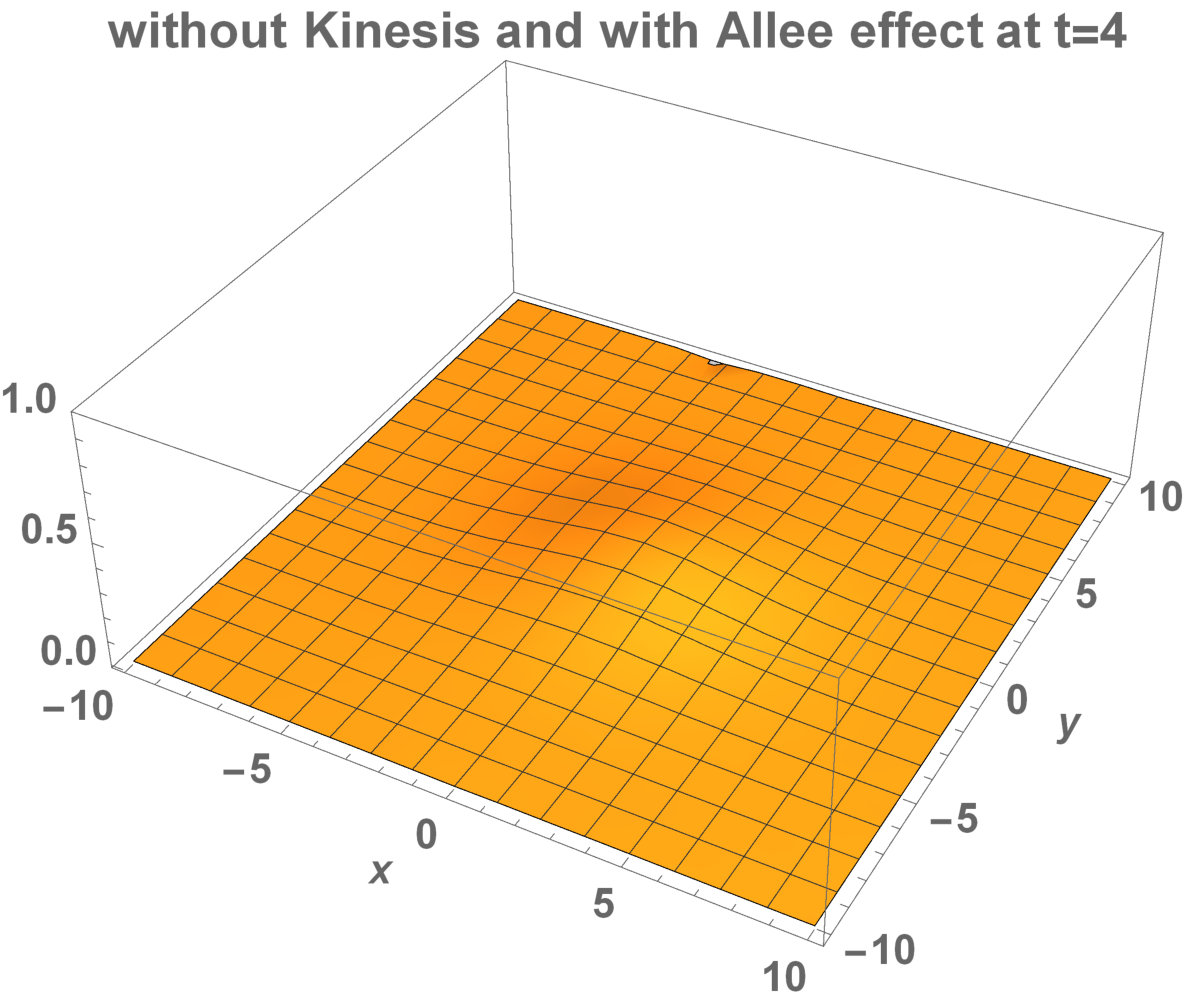 Figure 16
Figure 16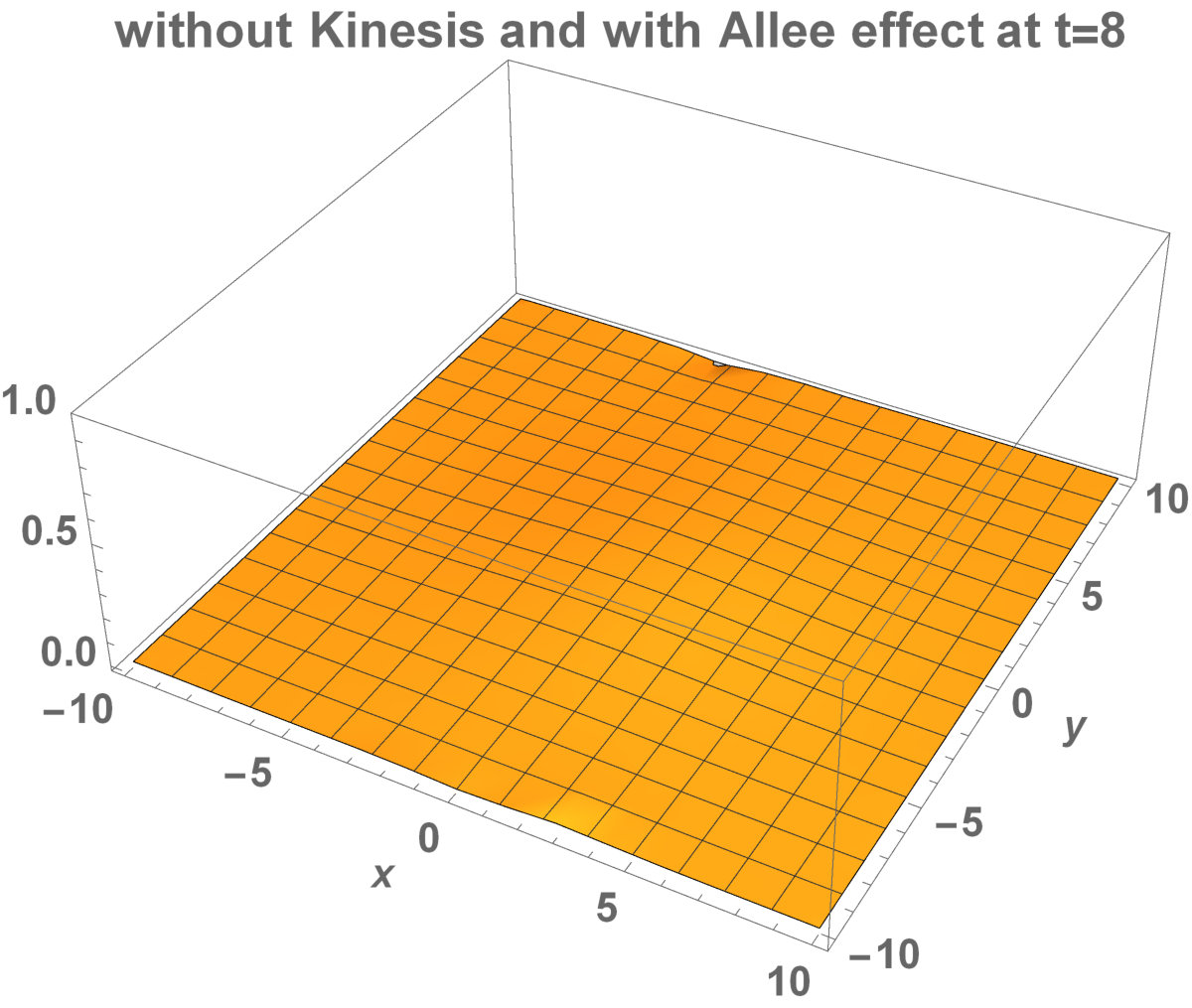 Figure 17
Figure 17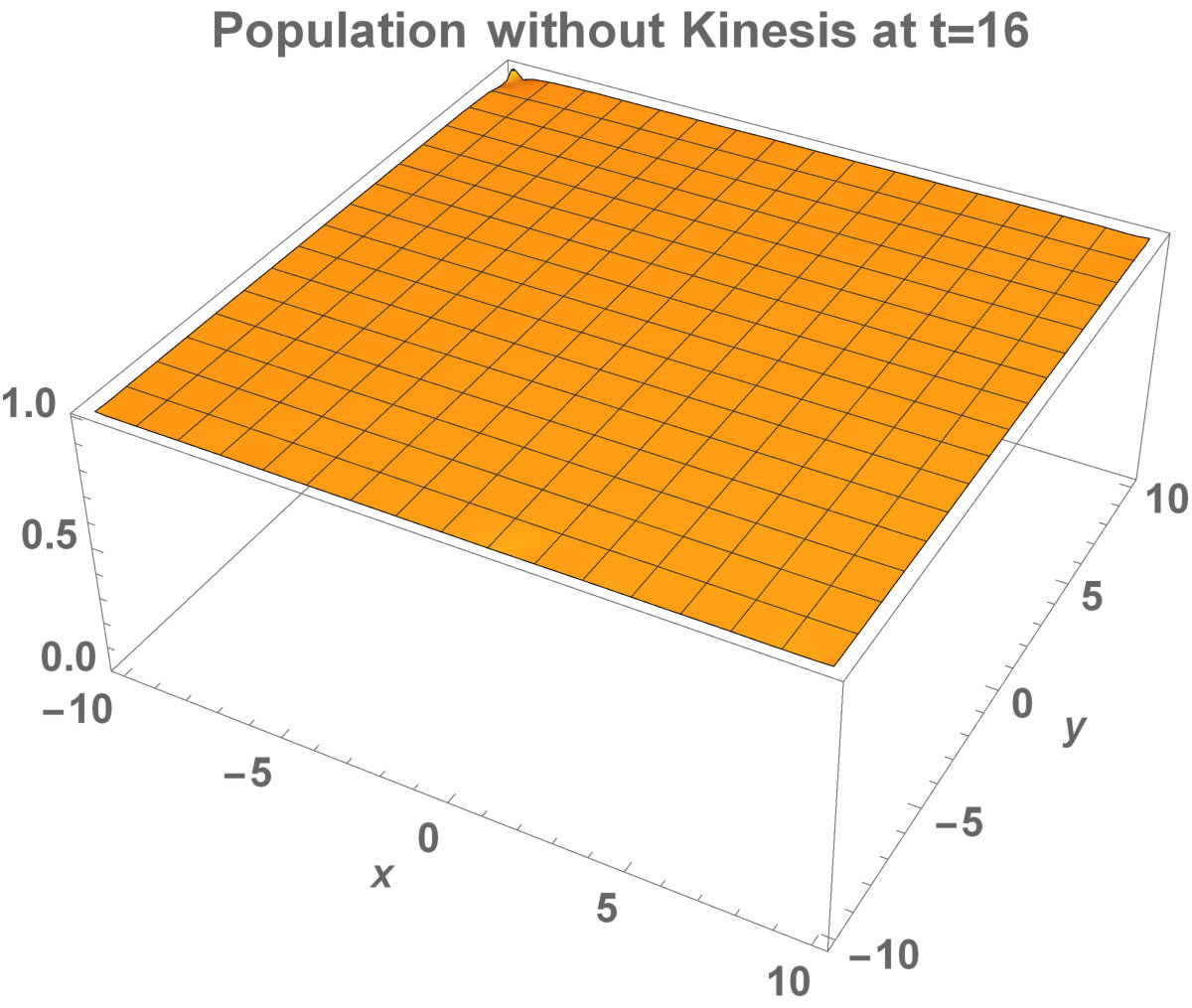 Figure 18
Figure 18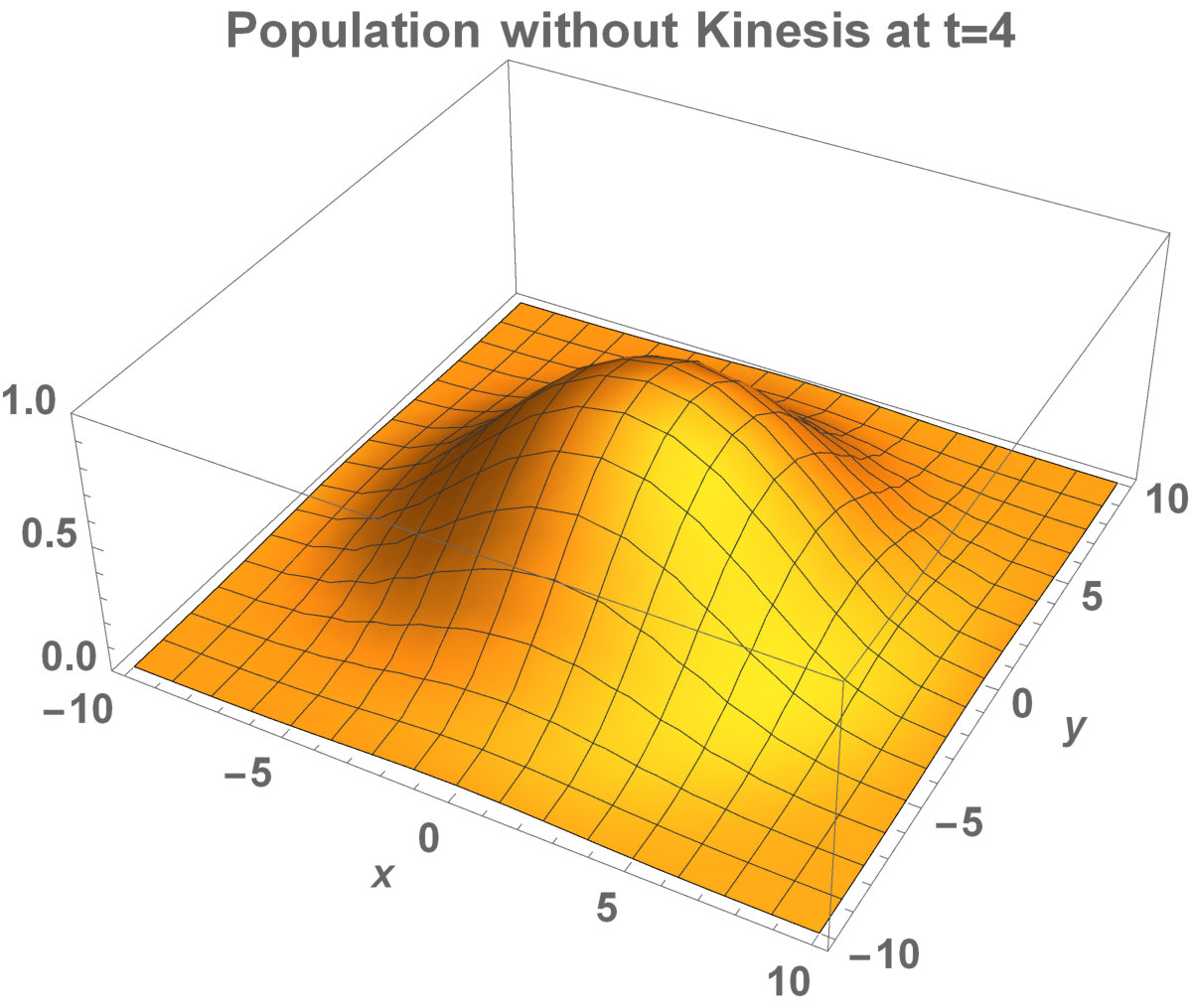 Figure 19
Figure 19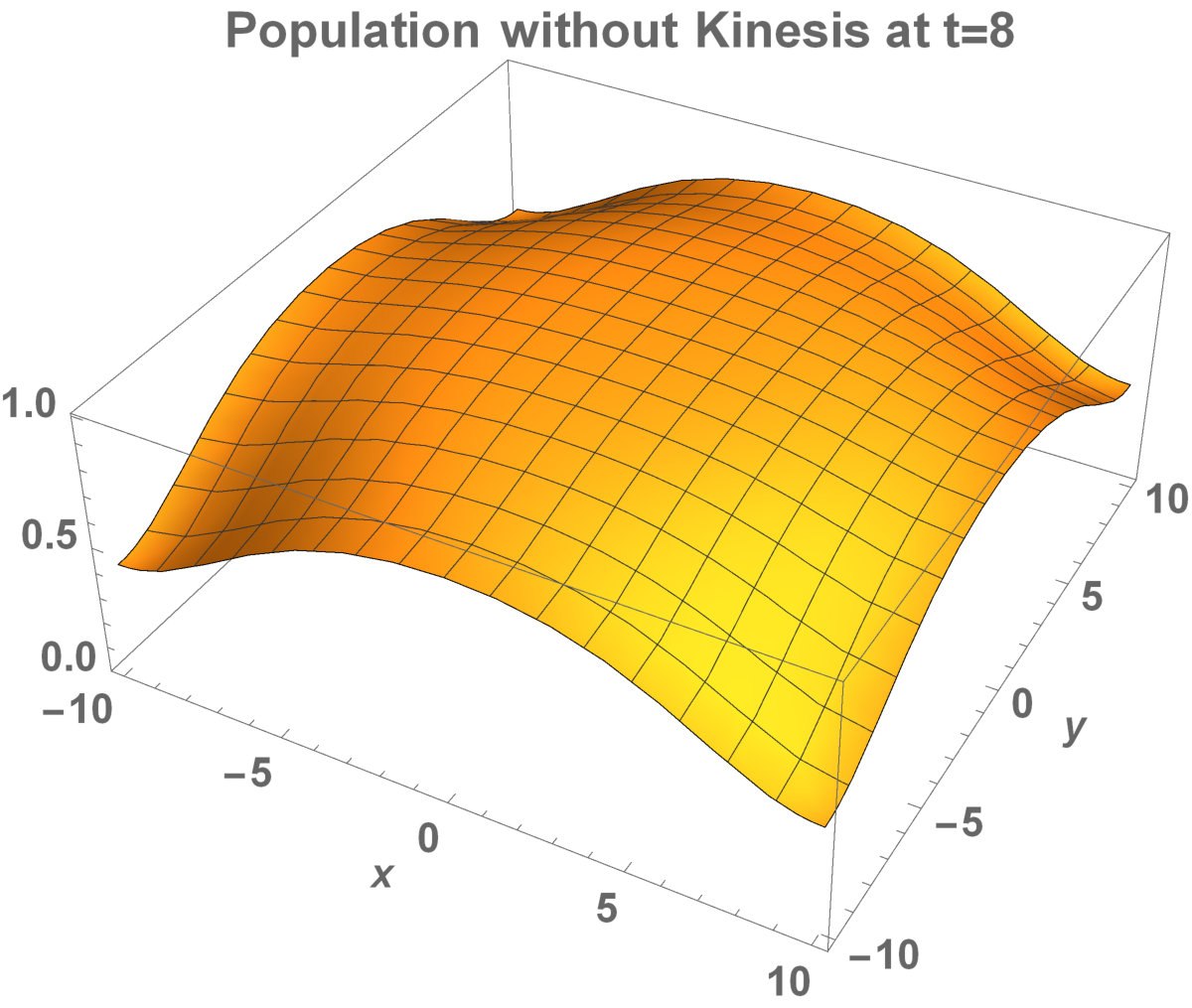 Figure 20
Figure 20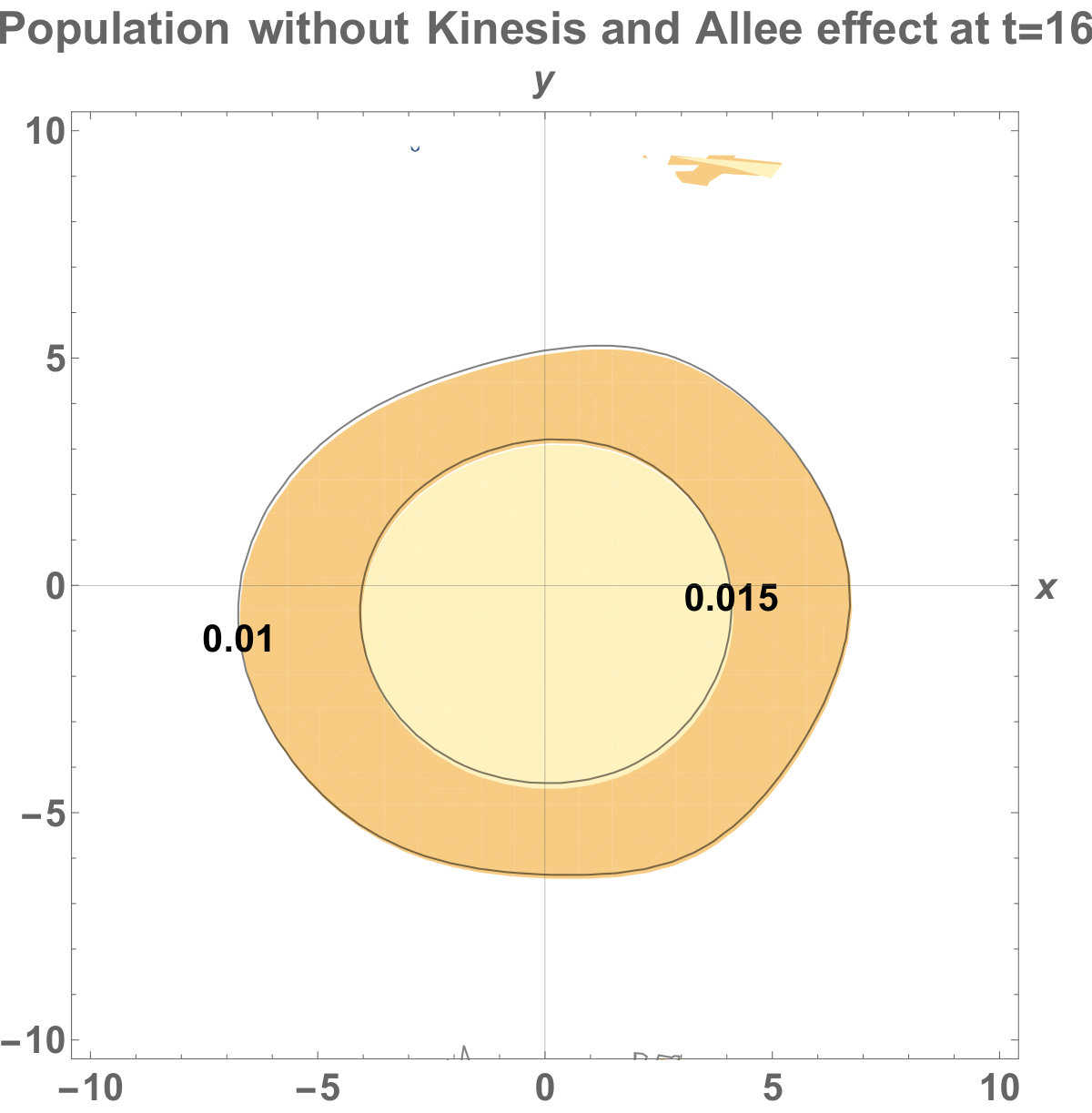 Figure 21
Figure 21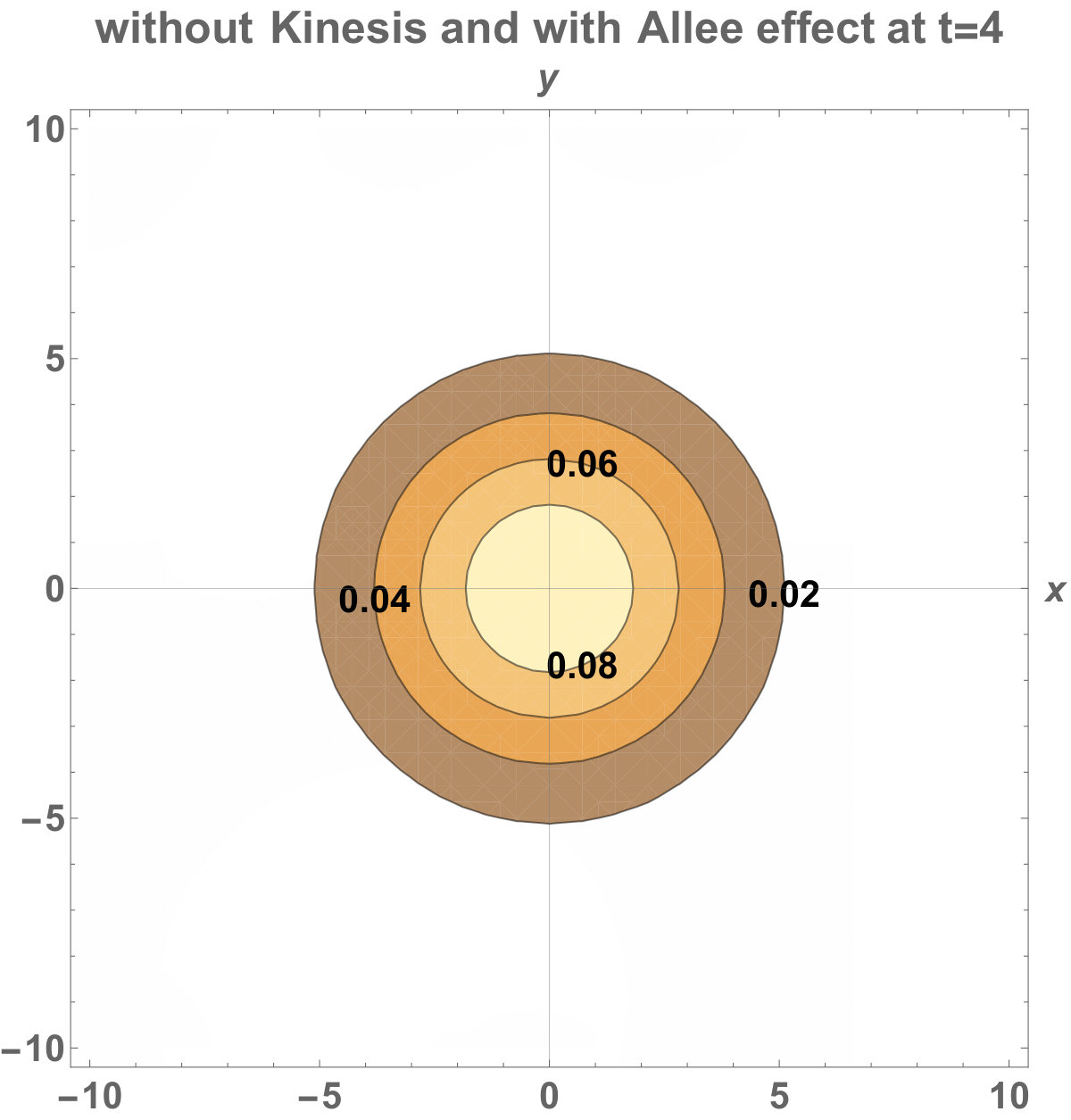 Figure 22
Figure 22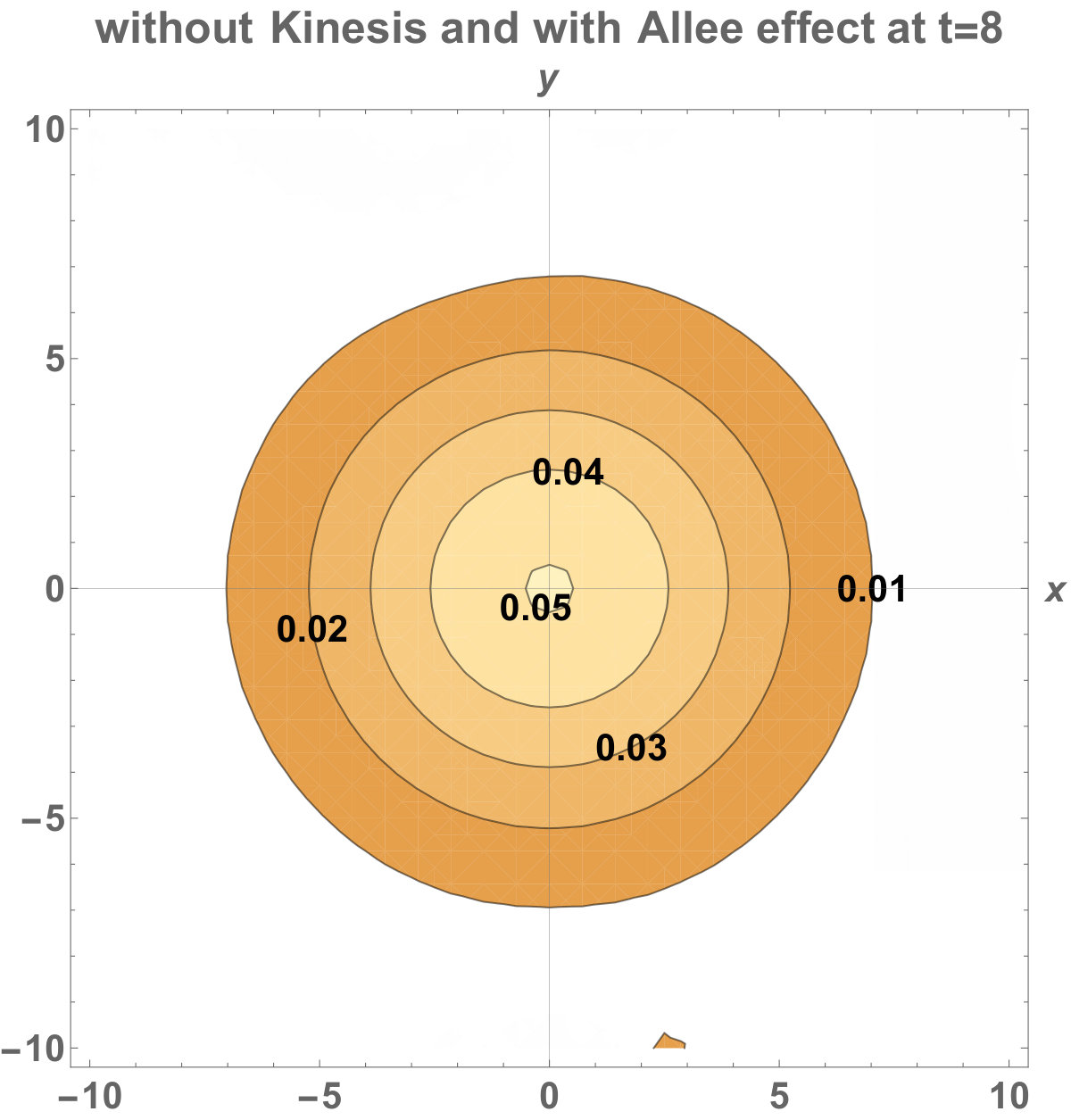 Figure 23
Figure 23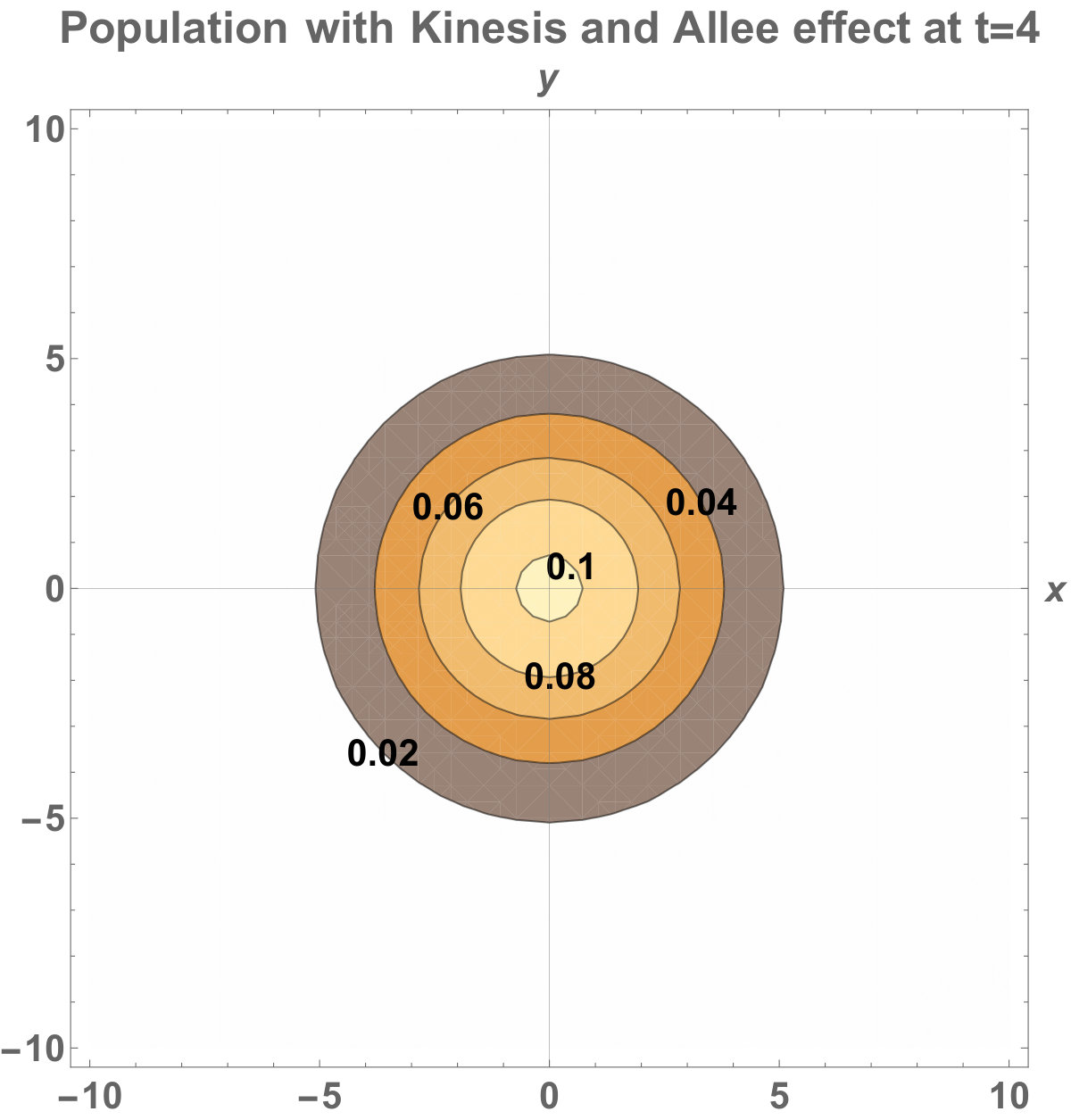 Figure 24
Figure 24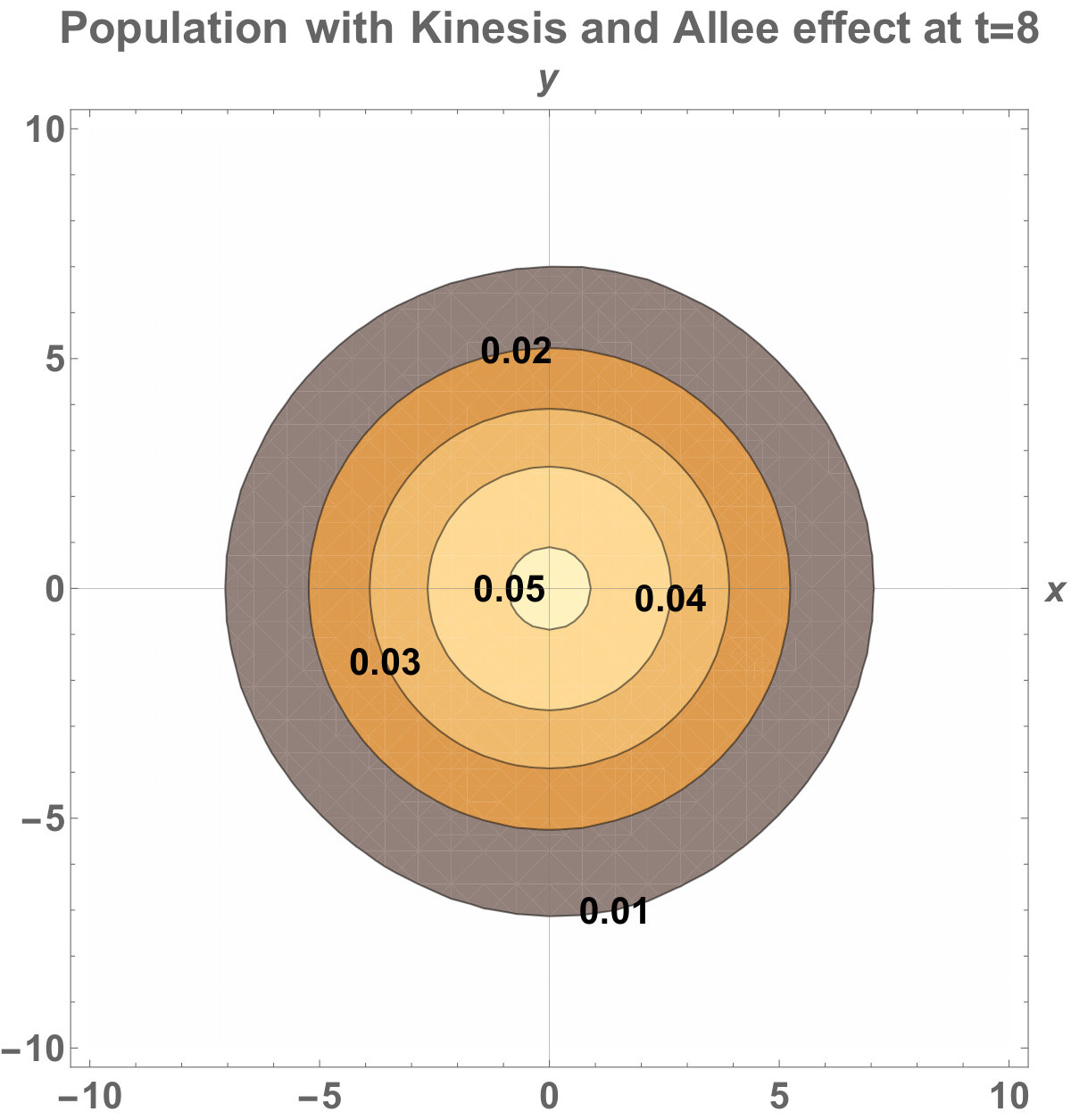 Figure 25
Figure 25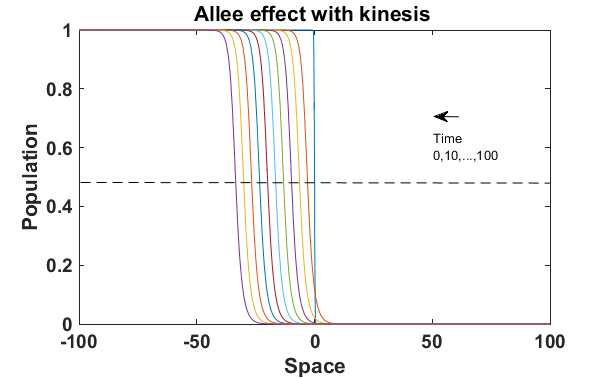 Figure 2
Figure 2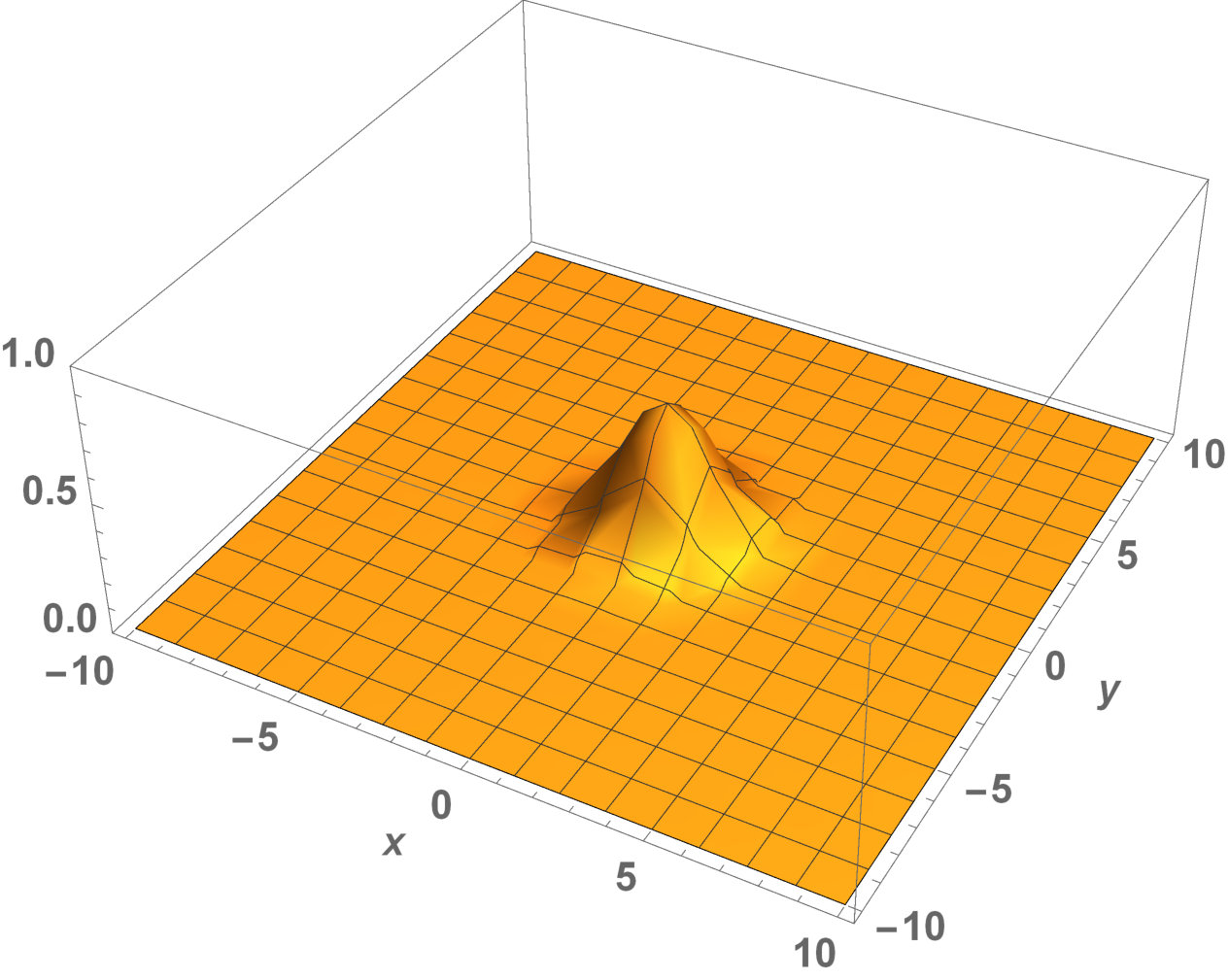 Figure 27
Figure 27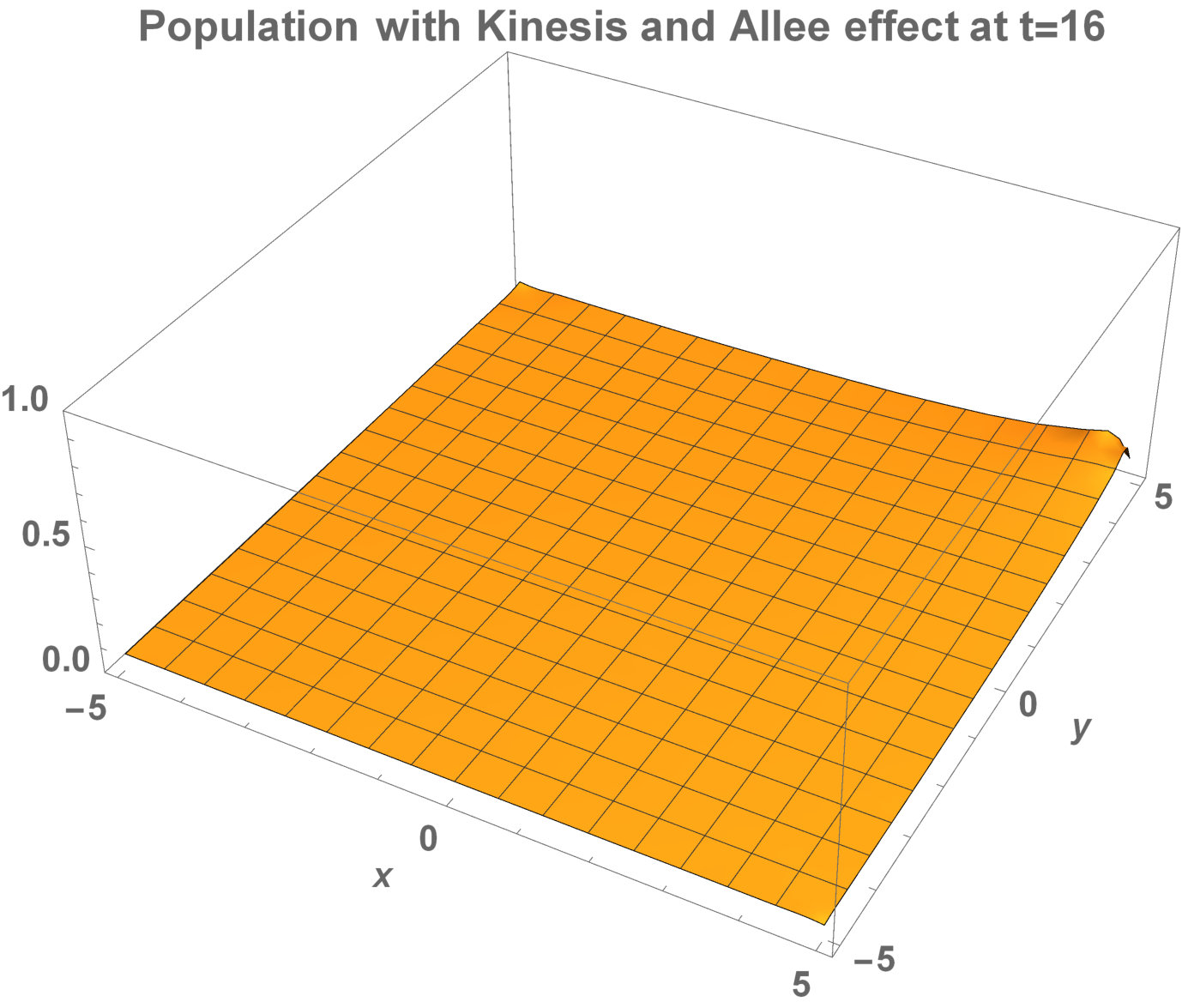 Figure 28
Figure 28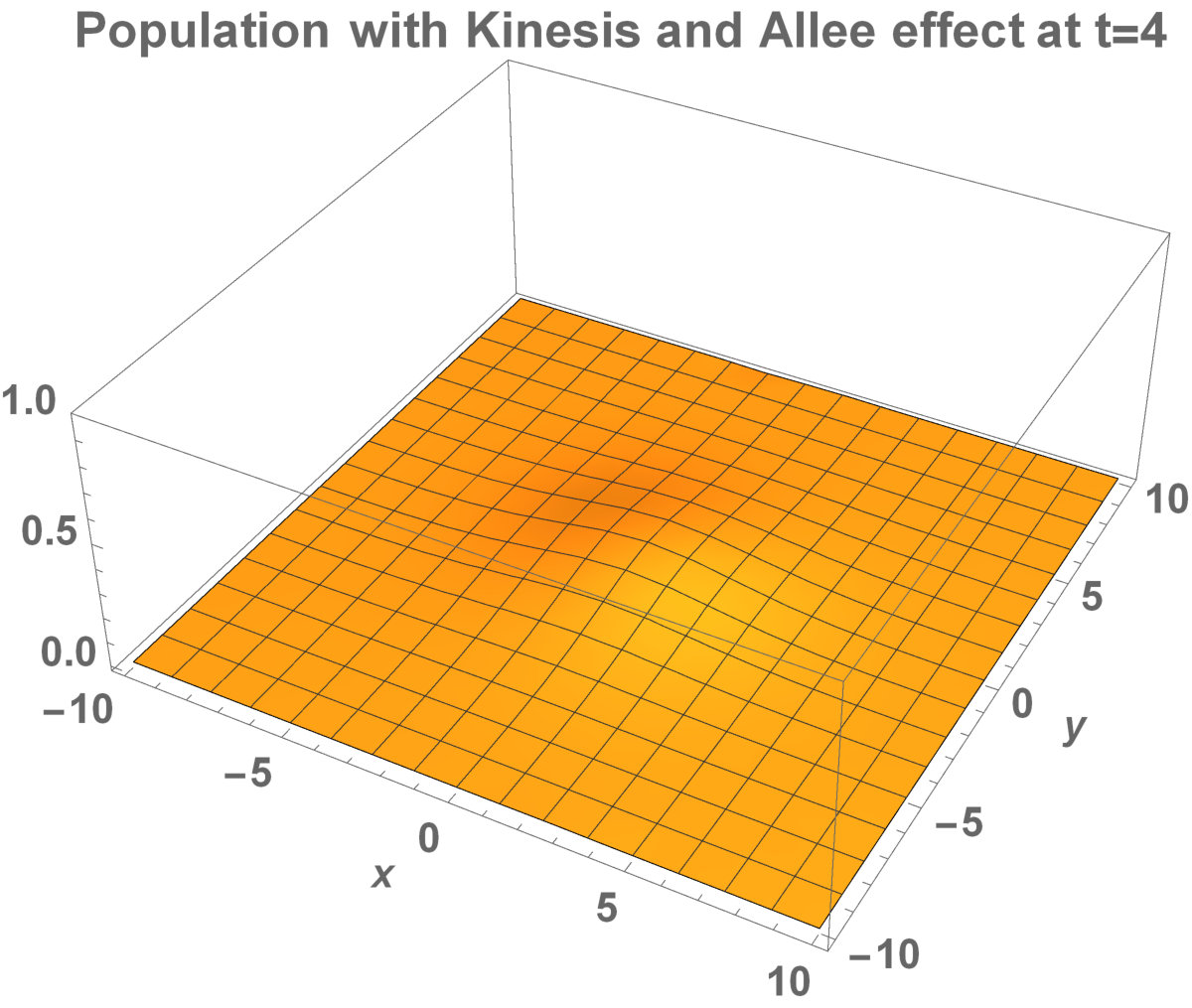 Figure 29
Figure 29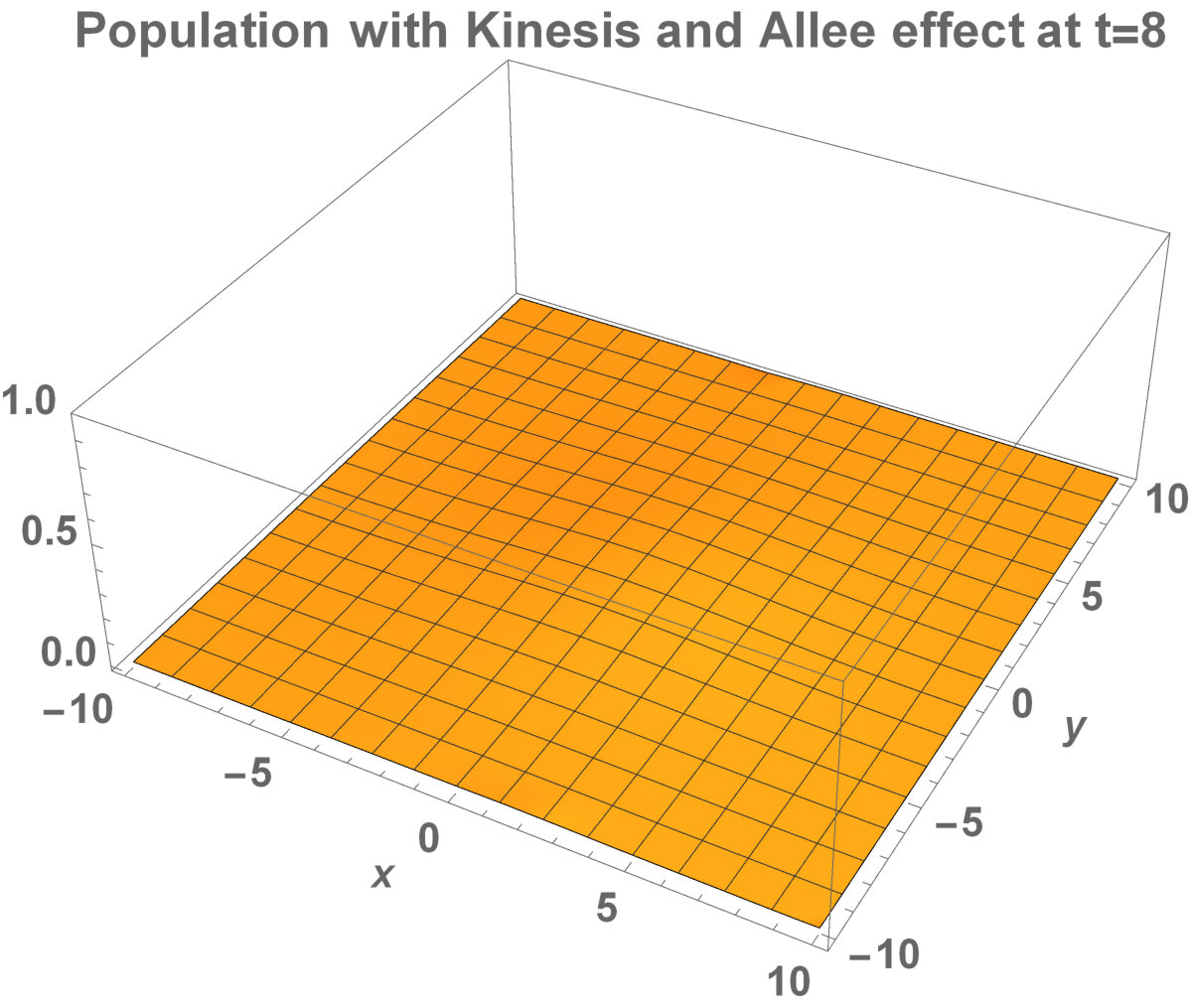 Figure 30
Figure 30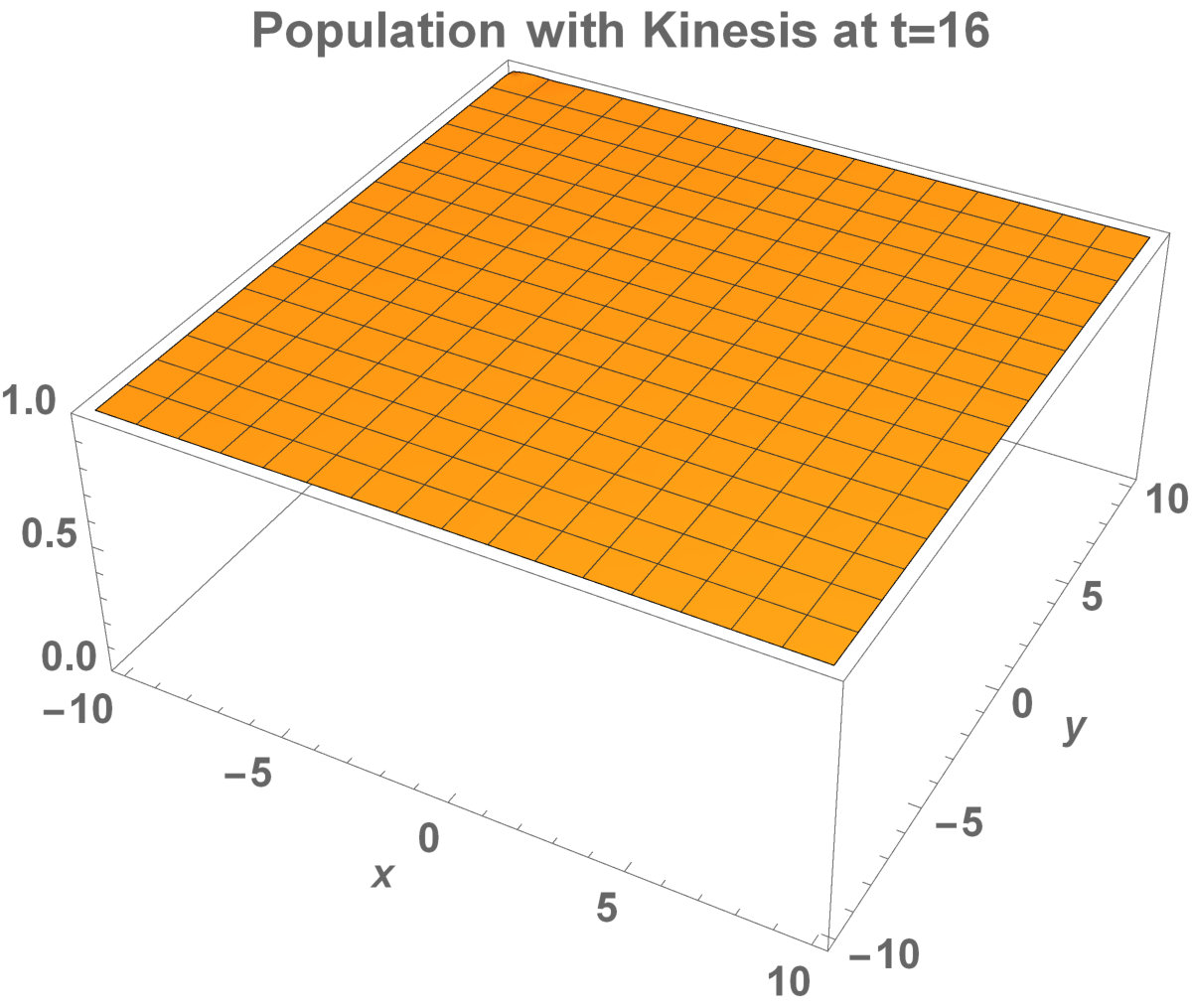 Figure 31
Figure 31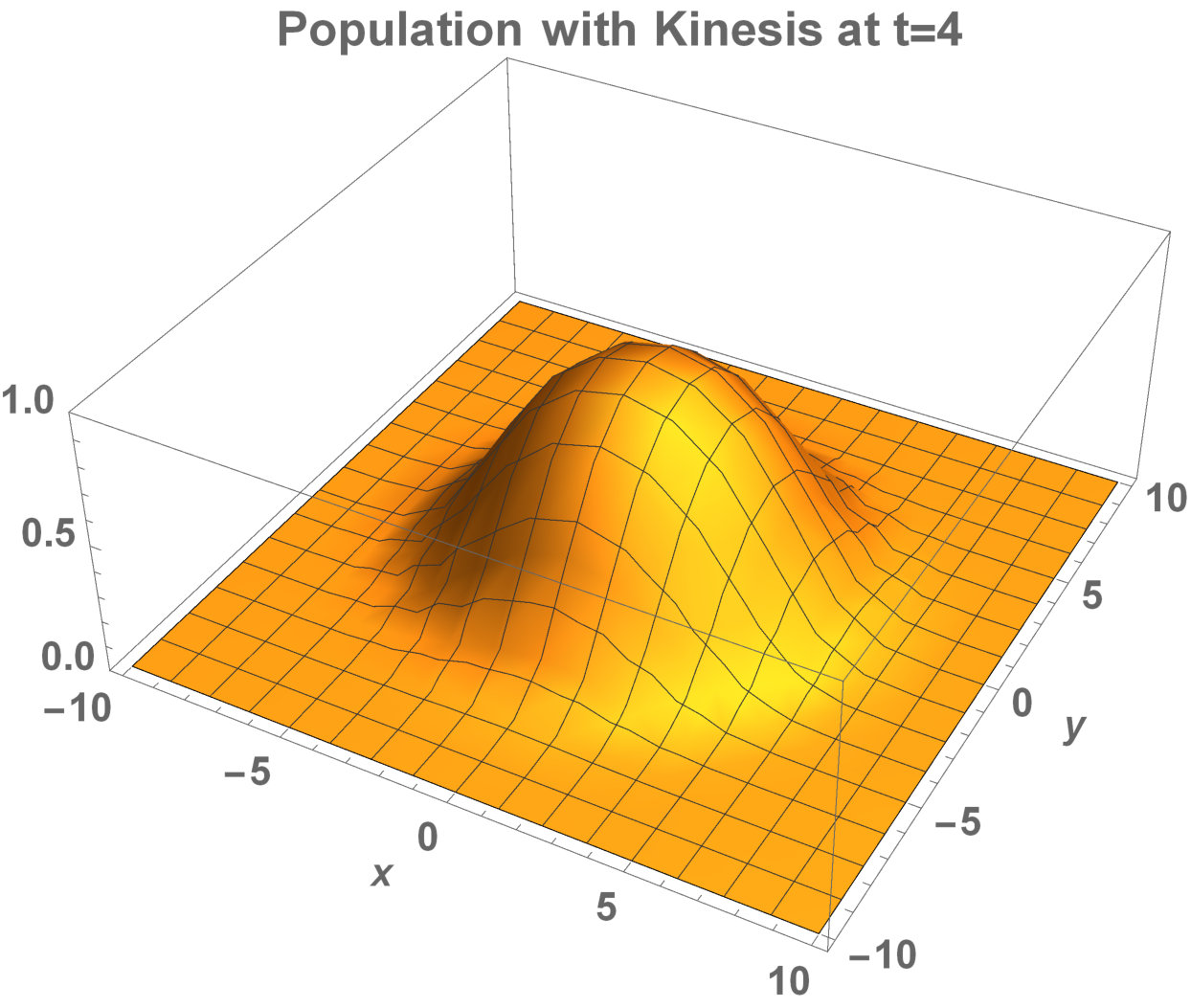 Figure 32
Figure 32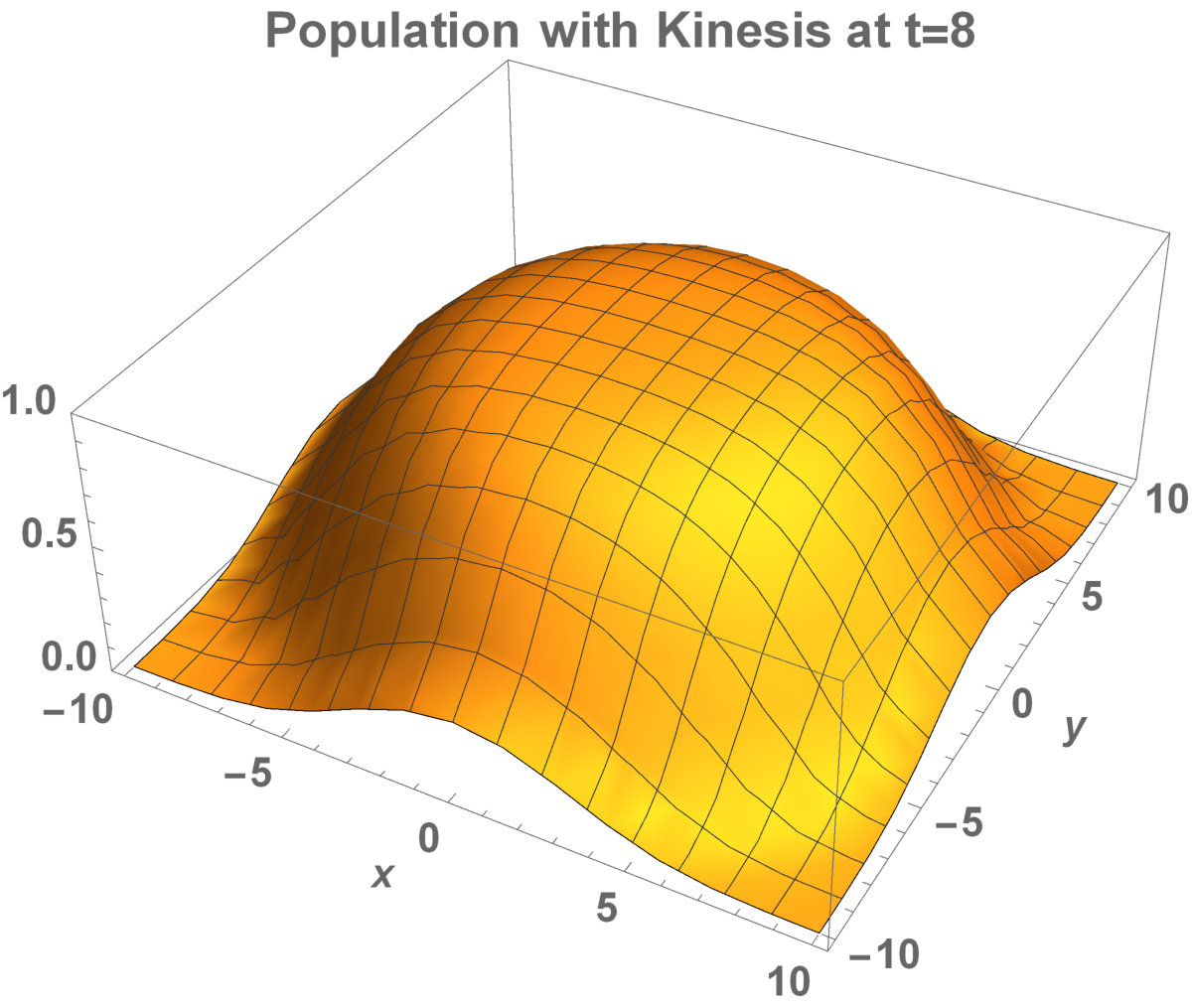 Figure 33
Figure 33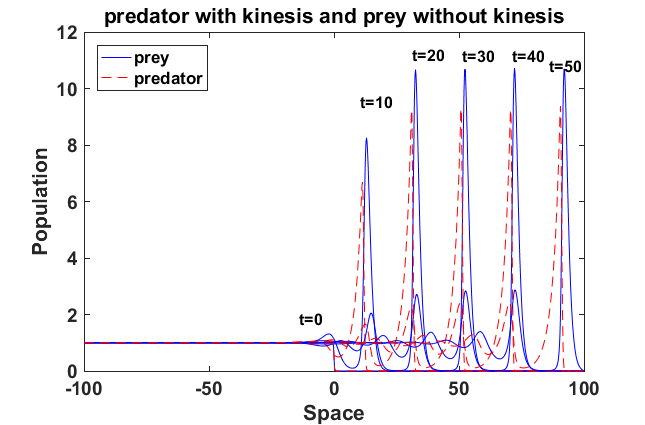 Figure 34
Figure 34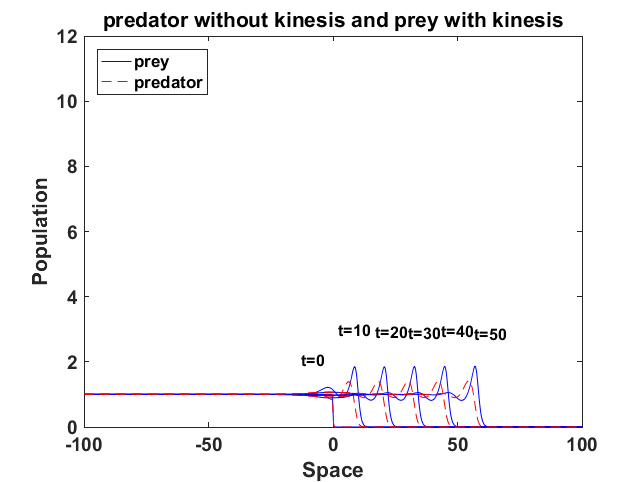 Figure 35
Figure 35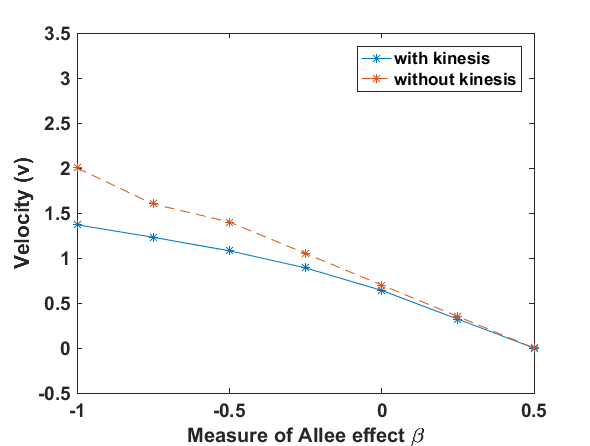 Figure 36
Figure 36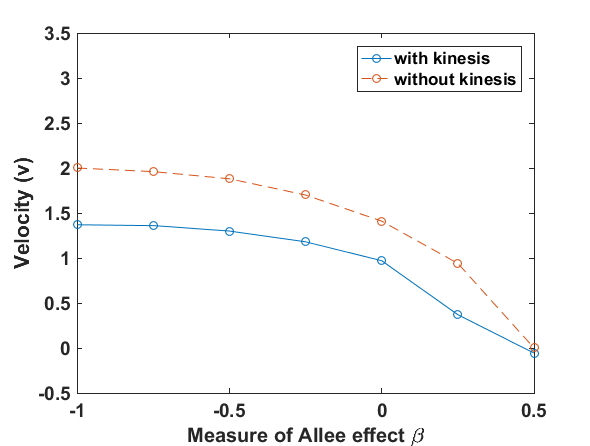 Figure 37
Figure 37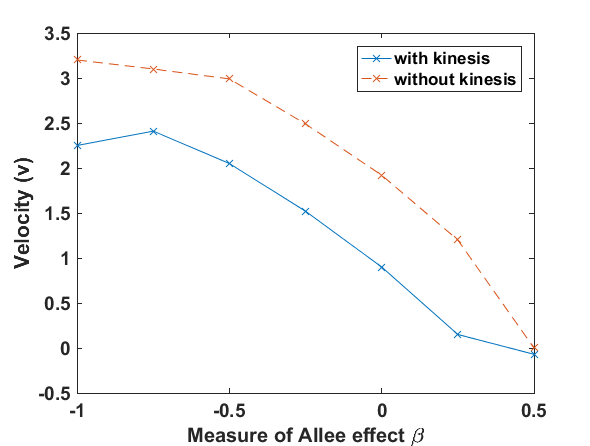 Figure 38
Figure 38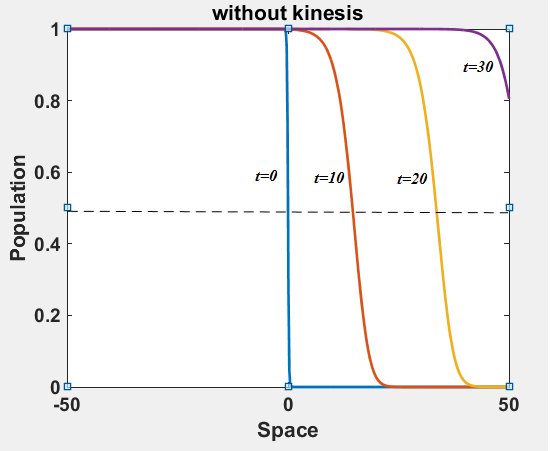 Figure 39
Figure 39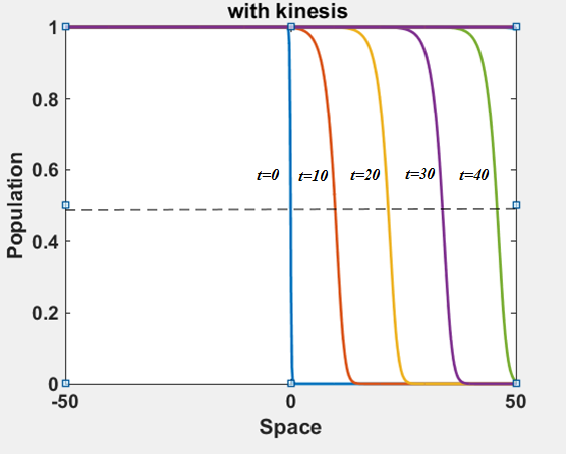 Figure 40
Figure 40Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMathematical Biology Tumor Growth · Gene Regulatory Network Analysis · Evolution and Genetic Dynamics
Impact of the purposeful kinesis on running waves
N. Çabukoǧlu
Department of Mathematics, University of Leicester, Leicester, LE1 7RH, UK
Abstract
The basic model of purposeful kinesis was developed recently (Gorban & Çabukoǧlu, Ecol. Complex. 2018, 33, 75–83) on the basis of the “Let well enough alone” idea. According to this model the diffusion drops while the reproduction coefficient is increasing. That is, species prefer to stay in a good condition and the population gives birth; otherwise, in the bad situation individuals want to run away because of the fatal conditions. In this study, we analyse the impact of the purposeful kinesis model on running waves. The running waves in the population with kinesis are studied using numerical experiments. Both monotonic and non-monotonic (Allee effect) dependence of the reproduction coefficient on the population density are studied. The possible benefits of the purposeful kinesis are demonstrated: with the higher diffusion, while the population without kinesis ends up with extinction, the population with kinesis stays alive and has the running wave behaviour. While the kinesis of the prey population is decreasing, the wave amplitude gets smaller. On the other hand, for the lower kinesis of predators they have a sharp increase.
keywords:
kinesis , diffusion , population , extinction , Allee effect , travelling waves
1 Introduction
In this study, we have analysed the diffusion model with kinesis. Kinesis is the non-directional movement according to the change of the local environment. That is, the organism gets the information from the living area, and therefore prefer to stay or move to the other beneficial place. Taxis is also related to mobility. However, taxis requires the non-local information. The organism with taxis has the directed movement towards to the condition or opposite side of the stimulus.
We aim to explore the impact of purposeful kinesis on running waves using PDE models. The classical PDE model of population dispersal was proposed by Patlak [1953] and Keller and Segel [1971]. It was used to model taxis behaviour [Hillen and Painter, 2009]. The reaction-diffusion model can be used for kinesis modelling. The diffusion coefficient can depend on the local situation. Cosner [2014] studied the reaction-diffusion models on animal dispersal. He presented the study on animal movement: which is the better condition to stay or leave, being slow or random movement, increasing the population size is harmful or not.
In this study, we employ the purposeful kinesis model [Gorban and Cabukoglu, 2018a]. Often ‘Purposeful’ means intentionality that individuals are unable. Rosenblueth and Wiener [1950] developed the general concept of purposeful behaviour. ‘Purpose’ brings the optimization idea and this concept requires the evolutionary optimality [Parker and Smith, 1990].
There is a connection between the reproduction rate and diffusion coefficient. Average reproduction coefficient was defined as Darwinian fitness [Haldane, 1932, Metz et al., 1992, Gorban, 2007]. Migration should increase Darwinian fitness.
In this paper, we analysed impact of the purposeful kinesis on running waves. In particular, it was presented that the population with Allee effect can spread late but with a higher diffusion the population will stay alive, and in time will continue the running wave behaviour. We did numerical experiments that kinesis model holds spreading invaded area, while the population without kinesis dies.
2 Main Results
In this section, we will give the main results.
2.1 Running Waves
The kinesis model was selected in the following form [Gorban and Cabukoglu, 2018a]:
[TABLE]
where:
- is the th species-population density,
- is the abiotic characteristics of the living conditions,
- is the reproduction coefficient,
- is the equilibrium diffusion coefficient which is defined when the reproduction coefficient is 0,
- defines the relation between the diffusion coefficient on the reproduction coefficient.
characterizes the diffusion depending on reproduction coefficient. According to that model, the diffusion depends on well-being which is measured by reproduction coefficient.
Gorban and Cabukoglu [2018b] later studied the cost of mobility. The changes of reproduction coefficient give the mobility cost value. If we take into account the cost of mobility then the bifurcation is predicted: when the conditions are getting worse, the mobility increases to a threshold level, and then it vanishes. Therefore, in the worse conditions, there is no solution for mobility. This mobility cost equation can be solved with Lambert -function.
We will present below how the dependence of diffusion on well-being effects the running waves of the population density on space.
Let us compare two models:
The PDE model for population with the constant diffusion coefficient: without kinesis, KPP (Kolmogorov, Petrovsky and Piskunov, 1937):
[TABLE]
- 2.
The PDE model for population with kinesis (KPP with modified diffusion coefficient [KPP, 1937]):
[TABLE]
We have first used MATLAB pdepe [2017] function to solve one dimensional system of PDE. Then, we used the MATHEMATICA NDSolve [2014] solver which is applied the Hermite method and Newton’s divided difference formula to solve two dimensional system.
We selected the space on the interval with Dirichlet boundary conditions and with the initial conditions:
[TABLE]
The values of the constants are: , , .
In the large time the running waves converge to a unique front with the velocity . For example, the logistic growth model speed will be .
Fig. 1 presents that the proposed ‘minimal purposeful kinesis model’ has the running wave behaviour on space. In these given conditions, the waves in the population model with kinesis is slower which is expalined in Fig. 2. When we increase the kinesis parameter , the spreading velocity will decrease monotonically.
We have done numerical experiments on MATLAB to see the relations between the wave velocity and the kinesis parameter (see Fig. 2). While is increasing, the velocity starts to decrease monotonically. After , the velocity is almost stabilizing. That is, the population spreading on space can be almost the same in the different time lines.
2.2 Running Waves with the Allee effect
Allee effect was introduced as nonmonotonic behaviour of the reproduction coefficient as a function of population density. At the beginning, the reproduction rate is minimum and then reaches to the peak. After the maximum reproduction rate, it starts to decrease monotonically while the population size is increasing [Allee et al., 1949]. There are a lot of reasons why populations show Allee effect such as low rate feeding [Way and Banks, 1967, Way and Cammell, 1970], and for some reasons the population reduces the predator defence [Kenward, 1978, Kruuk, 1964], give up the breeding for psychological reasons [Ralls et al., 1986, Soule et al., 1986], and a wide variety of reasons [Caraco and Pulliam, 1984, Folt, 1987, Foster and Treherne, 1981, Turchin and Kareiva, 1989].
Lewis and Kareiva [1993] explored the population spread of asymptotic rates with the Allee effect. In two dimensional analysis, they investigated that the patterns of spread have been influenced by this population model. We have used the coefficients in that study to analyse how the population with kinesis affects the two-dimensional patterns of spread.
The basic form of the reproduction coefficient with Allee effect is
[TABLE]
The models with purposeful kinesis on Allee effect were studied by Gorban and Cabukoglu [2018a] previously.
Gorban and Sadovskiy [1989] introduced the study that when the average population size is less than the optimum density, the evolutionary optimal strategy for populations with the Allee effect collapses with optimal density. There are some other consequences of Allee effect with diffusion: the population spreads in a massive way through the formation, the interaction between species and the movement of separate patches even if the population is inhomogeneous external condition [Morozov et al., 2006, Petrovskii et al., 2002].
We may present the reaction-diffusion equations for a single population with the Allee effect for a system without kinesis (4) and for a system with kinesis (5).
[TABLE]
[TABLE]
The values of constants are: , , .
Eqs. (4) and (5) are solved for one space variable with Dirichlet boundary conditions and with the initial conditions:
[TABLE]
We choose as the normalization constant which was given by Lewis and Kareiva [1993] as determined by the maximum growth rate. has been determined as
[TABLE]
[TABLE]
[TABLE]
It is considered that travelling wave solutions to Eqs. (4) and (5) are in the form of with , where is the velocity.
It was shown that there is a unique solution to these Eqs. (4) and (5) with a unique velocity with the condition by Fife [1979]. When the Allee effect parameter is negative, there is a minimum value of velocity such that there exists a corresponding running wave solution [Aronson and Weinberg, 1978, Fife, 1979, Hadeler and Rothe, 1975]. After a long time period , the solution of the system 4 may converge to a travelling wave solution.
The numerical results (Fig. 3) show that the population with Allee effect has the running wave behaviour both with kinesis and without kinesis. Population with kinesis demonstrates faster running waves on space. Fast-spreading effect of the population could also end up to extinction for the population with Allee effect. When the Allee effect is higher than the population density, the reproduction coefficient becomes negative. Therefore, this leads the population to extinction.
Wave velocity for different models has been displayed as the Fig. 4 with the Allee effect parameter . The Allee effect decreases the rate of spread of an invading population. We can see from the Fig. 4 that with these conditions the spreading of the population with the Allee effect is decreasing when is getting higher value. Moreover, the spreading velocity of the population without kinesis is faster than the population with kinesis.
2.3 Two-Dimensional Spread
Analytical representation of two-dimensional system may be too complex to solve. Aronson and Weinberg [1978] presented that the planar travelling waves exist and this form can be used by an invading population. They studied the two-dimensional population model with Allee dynamics to see the invaded area of the population. The population, with Allee effect, may end up with extinction. This can happen even if the population density exceeds the threshold. The reason is that reproduction, the population growth, cannot be sufficient to prevent the population dispersal. There are some critical factors to determine the invading success: shape and size of the environment. It has been already shown that how the population stay alive between invaded areas and unoccupied places [Lewis and Kareiva, 1993].
Now, in this section, we present the two-dimensional model that we want to see the invaded and unoccupied areas of the population with kinesis and without kinesis. In addition to that model, we discuss the population with Allee effect.
Fig. 5 shows the initial distribution that the invaded area is the same for the population with kinesis and without kinesis.
3 Predator-Prey Running Waves
In this section, we consider competing species in the same environment. These two species interact with the model without kinesis and the new model with kinesis. Volpert and Petrovskii analysed the running waves of predator-prey interaction model in their study [Volpert and Petrovskii, 2009]. Now, we compare the population with kinesis and without kinesis as running wave behaviour.
The PDE model for population with the constant diffusion coefficient (i.e. without kinesis):
[TABLE]
- 2.
The PDE model for population with kinesis:
[TABLE]
We consider the simplest predator-prey model in the classical Lotka-Volterra form in dimensionless variables and ; and are the population densities of prey and predator.
Eqs. (10), (11), (12) and (13) are solved for one space variable with Dirichlet boundary conditions and with the initial conditions:
[TABLE]
The numerical simulations with the 1D case show that depending on the values the systems (10), (11), (12) and (13) show a variety of travelling fronts.
We compare four cases with Figs. 16 and 17:
both predator and prey without kinesis,
- 2.
both predator and prey with kinesis,
- 3.
predator without kinesis and prey with kinesis,
- 4.
predator with kinesis and prey without kinesis.
All of them have the running wave behaviour. We demonstrate with Fig. 16b that the predator and prey populations with kinesis have the running wave behaviour and the peaks between predator and prey increase. For the predator without kinesis and prey with kinesis (Fig. 17a), the amplitude decreases. On the other hand, the predator with kinesis and prey without kinesis (Fig. 17b) running wave peaks of the population density become sharper.
4 Discussion
It has been suggested a purposeful kinesis model that the diffusion coefficient depends on well-being which is measured by the reproduction coefficient by Gorban and Cabukoglu [2018a]. The system formalizes the simple rule: “Let well enough alone”. Volpert and Petrovskii presented the study on travelling waves of the reaction-diffusion model Volpert and Petrovskii [2009].
In this paper, we analysed the running wave behaviour of the reaction-diffusion model with kinesis. The running wave theory begins with Fisher [1930] and Kolmogorov, Petrovskii and Piskunov [KPP, 1937]. They defined the travelling wave solutions of the scalar reaction-diffusion equation and analysed their existence, stability and the speed of these waves. Lewis and Kareiva explored the relevance between the Allee effect parameter and the relative wave speed. They also studied on the two-dimensional spread of invading populations with Allee effect.
In this study, we have demonstrated the speed of the running waves of the population with kinesis. These following have been observed:
While the relation between the diffusion coefficient and reproduction coefficient is increasing the running wave speed monotonically decreases, and at some point stabilizes (see Fig. 2).
- 2.
The Allee effect parameter and velocity have been displayed by Fig. 4 for the values of normalization coefficient on the running wave front . While the Allee effect parameter is increasing, the populations’ running speed decreases. When , the velocity is 0. After that value of , the waves of the population model show the running wave behaviour through the left hand side. The individuals with kinesis run slower than the population without kinesis. This may lead up an extinction in the invaded area.
- 3.
The initial distributions of the invaded area are the same for both populations (see Fig. 5). System stabilization is observed by . Therefore, Kinesis does not change the stability of homogeneous positive steady-states.
- 4.
The invasion of the population with kinesis occupies the larger area than without kinesis. That is, they stay alive in the habitat.
- 5.
Allee dynamics can affect the population extinction (see Fig. 14). On the other hand, with the higher diffusion, the population with kinesis do not extinct and can invade a large area (see Fig. 15).
- 6.
We have also presented the running wave behaviour of the predator-prey model with kinesis. The constant diffusion model waves were recently analysed by Volpert and Petrovskii [2009] for the predator-prey model. We have observed also that this model can have running wave behaviour. Kinesis of prey decreases amplitude of the wave smaller, whereas kinesis of the predators can significantly increase the peak value (Fig. 17b).
These population models with kinesis can be adapted to several models. It can be seen that individuals may have the running wave behaviour in the invaded areas.
Acknowledgement
I am grateful to A.N. Gorban for his advises.
References
- Allee et al. [1949] Allee, W.C., Park, O., Emerson, A.E., Park, T., Schmidt, K.P., 1949. Principles of animal ecology. Saunders, Philadelphia.
- Aronson and Weinberg [1978] Aronson, D. G., and Weinberger, H. F. 1978. Multidimensional nonlinear diffusion arising in population genetics. Advances in Mathematics, 30(1), 33-76.
- Caraco and Pulliam [1984] Caraco, T. and Pulliam, H.R., 1984. Sociality and survivorship in animals exposed to predation.
- Cosner [2014] Cosner, C., 2014. Reaction-diffusion-advection models for the effects and evolution of dispersal. Discrete Contin. Dyn. Syst. 34, 1701–1745.
- Fife [1979] Fife, P.C., 1979. Lecture notes in biomathematics. In Mathematical Aspects of Reacting and Diffusing Systems (Vol. 28). Springer-Verlag Berlin.
- Fisher [1930] Fisher, R.A., 1999. The genetical theory of natural selection: a complete variorum edition. Oxford University Press.
- Folt [1987] Folt, C.L., 1987. An experimental analysis of costs and benefits of zooplankton aggregation. Predation, Direct and Indirecct Impacts on Aquatic Communities, pp.300-314.
- Foster and Treherne [1981] Foster, W.A. and Treherne, J.E., 1981. Evidence for the dilution effect in the selfish herd from fish predation on a marine insect. Nature, 293(5832), p.466.
- Gorban [2007] Gorban, A.N., 2007. Selection theorem for systems with inheritance. Math. Model. Nat. Phenom. 2(4), 1–45.
- Gorban and Cabukoglu [2018a] Gorban, A.N. and Çabukoǧlu, N., 2018. Basic model of purposeful kinesis. Ecological Complexity, 33, pp.75-83.
- Gorban and Cabukoglu [2018b] Gorban, A.N. and Çabukoǧlu, N., 2018. Mobility cost and degenerated diffusion in kinesis models. Ecological Complexity, 36, pp.16-21.
- Gorban and Sadovskiy [1989] Gorban, A.N., Sadovskiy, M.G., 1989. Optimal strategies of spatial distribution: the Allee effect. Zh. Obshch. Biol. 50(1), 16–21.
- Hillen and Painter [2009] Hillen, T., Painter, K.J., 2009. A user’s guide to PDE models for chemotaxis. J. Math. Biol. 58, 183–217.
- Hadeler and Rothe [1975] Hadeler, K.P. and Rothe, F., 1975. Travelling fronts in nonlinear diffusion equations. Journal of Mathematical Biology, 2(3), pp.251-263.
- Haldane [1932] Haldane, J.B.S., 1932. The Causes of Evolution, Longmans Green, London.
- Keller and Segel [1971] Keller, E.F., Segel, L.A., 1971. Model for chemotaxis. J. Theor. Biol. 30(2), 225–234.
- Kenward [1978] Kenward, R.E., 1978. Hawks and doves: factors affecting success and selection in goshawk attacks on woodpigeons. The Journal of Animal Ecology, pp.449-460.
- KPP [1937] Kolmogorov, A.N., Petrovsky,I.G. and Piskunov, N.S., 1937. Investigation of the equation of diffusion combined with increasing of the substance and its application to a biology problem Bull Moscow State Univ Ser A: Math and Mech, 1 (6), pp. 1-25
- Kruuk [1964] Kruuk, H., 1964. Predators and Anti-predator Behaviour of the Black-headed Gull:(Laurus Ridibundus L.) (Vol. 11). Brill Archive.
- Lewis and Kareiva [1993] Lewis, M.A. and Kareiva, P., 1993. Allee dynamics and the spread of invading organisms. Theoretical Population Biology, 43(2), pp.141-158.
- Metz et al. [1992] Metz, J.A., Nisbet, R.M., Geritz, S.A., 1992. How should we define ‘fitness’ for general ecological scenarios?. Trends. Ecol. Evol. 7(6), 198–202.
- Morozov et al. [2006] Morozov, A., Petrovskii, S., Li, B.L., 2006. Spatiotemporal complexity of patchy invasion in a predator-prey system with the Allee effect. J. Theor. Biol. 238(1), 18–35.
- NDSolve [2014] NDSolve. 2014. Wolfram Language & System Documentation Center. urlhttp://reference.wolfram.com/language/ref/NDSolve.html.
- Parker and Smith [1990] Parker, G.A., Smith, J.M., 1990. Optimality theory in evolutionary biology. Nature 348(6296), 27–33.
- Patlak [1953] Patlak, C. S. 1953. Random walk with persistence and external bias. B. Math. Biophys. 15, 311–338.
- pdepe [2017] pdepe. 2017. Solve initial-boundary value problems for parabolic-elliptic PDEs in 1-D. MatWorks Documentation urlhttps://uk.mathworks.com/help/matlab/ref/pdepe.html.
- Petrovskii et al. [2002] Petrovskii, S.V., Morozov, A.Y., Venturino, E., 2002. Allee effect makes possible patchy invasion in a predator–prey system. Ecol. Lett. 5(3), 345–352.
- Ralls et al. [1986] Ralls, M., Harvey P., and Lyles, A. 1986. inbreeding in natural populations of birds and mammals, in ”Conservation Biology” (M. Soule Ed.), pp. 35-56, Sinauer Associates, Massachusetts.
- Rosenblueth and Wiener [1950] Rosenblueth, A., Wiener, N., 1950. Purposeful and non-purposeful behavior. Philos. Sci. 17(4), 318–326.
- Soule et al. [1986] Soule, M., Gilpin, M., Conway, W. and Foose, T., 1986. The millenium ark: how long a voyage, how many staterooms, how many passengers?. Zoo biology, 5(2), pp.101-113.
- Turchin and Kareiva [1989] Turchin, P. and Kareiva, P., 1989. Aggregation in Aphis varians: an effective strategy for reducing predation risk. Ecology, 70(4), pp.1008-1016.
- Volpert and Petrovskii [2009] Volpert, V., and Petrovskii, S. (2009). Reaction–diffusion waves in biology. Physics of life reviews, 6(4), 267-310.
- Way and Banks [1967] Way, M. J., and Banks, C. J., 1967. Intra‐specific mechanisms in relation to the natural regulation of numbers of Aphis fabae Scop. Annals of Applied Biology, 59(2), 189-205.
- Way and Cammell [1970] Way, M. J., and Cammell, M., 1970. Aggregation behaviour in relation to food utilization by aphids. In Animal populations in relation to their food resources. A symposium of the British Ecological Society, Aberdeen 24-28 March 1969. (pp. 229-247). Oxford and Edinburgh, Blackwell.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Allee et al. [1949] Allee, W.C., Park, O., Emerson, A.E., Park, T., Schmidt, K.P., 1949. Principles of animal ecology. Saunders, Philadelphia.
- 2Aronson and Weinberg [1978] Aronson, D. G., and Weinberger, H. F. 1978. Multidimensional nonlinear diffusion arising in population genetics. Advances in Mathematics, 30(1), 33-76.
- 3Caraco and Pulliam [1984] Caraco, T. and Pulliam, H.R., 1984. Sociality and survivorship in animals exposed to predation.
- 4Cosner [2014] Cosner, C., 2014. Reaction-diffusion-advection models for the effects and evolution of dispersal. Discrete Contin. Dyn. Syst. 34, 1701–1745.
- 5Fife [1979] Fife, P.C., 1979. Lecture notes in biomathematics. In Mathematical Aspects of Reacting and Diffusing Systems (Vol. 28). Springer-Verlag Berlin.
- 6Fisher [1930] Fisher, R.A., 1999. The genetical theory of natural selection: a complete variorum edition. Oxford University Press.
- 7Folt [1987] Folt, C.L., 1987. An experimental analysis of costs and benefits of zooplankton aggregation. Predation, Direct and Indirecct Impacts on Aquatic Communities, pp.300-314.
- 8Foster and Treherne [1981] Foster, W.A. and Treherne, J.E., 1981. Evidence for the dilution effect in the selfish herd from fish predation on a marine insect. Nature, 293(5832), p.466.