The Effects of Cold Stress on Photosynthesis in Hibiscus Plants
Miriam Paredes, María José Quiles

TL;DR
This study explores how cold stress affects photosynthesis in Hibiscus plants and how different parts of the plant respond to chilling.
Contribution
The study reveals how root temperature modifies cold stress effects and the role of cyclic electron flow and chlororespiration in protecting photosystems.
Findings
Cold stress reduces PSII efficiency and electron transport capacity in Hibiscus plants.
Cyclic electron flow helps protect PSII when the whole plant is chilled, but not when only the stem is chilled.
Chlororespiratory enzymes increase when only the stem is chilled, indicating their role in stress response.
Abstract
The present work studies the effects of cold on photosynthesis, as well as the involvement in the chilling stress of chlororespiratory enzymes and ferredoxin-mediated cyclic electron flow, in illuminated plants of Hibiscus rosa-sinensis. Plants were sensitive to cold stress, as indicated by a reduction in the photochemistry efficiency of PSII and in the capacity for electron transport. However, the susceptibility of leaves to cold may be modified by root temperature. When the stem, but not roots, was chilled, the quantum yield of PSII and the relative electron transport rates were much lower than when the whole plant, root and stem, was chilled at 10°C. Additionally, when the whole plant was cooled, both the activity of electron donation by NADPH and ferredoxin to plastoquinone and the amount of PGR5 polypeptide, an essential component of the cyclic electron flow around PSI, increased,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
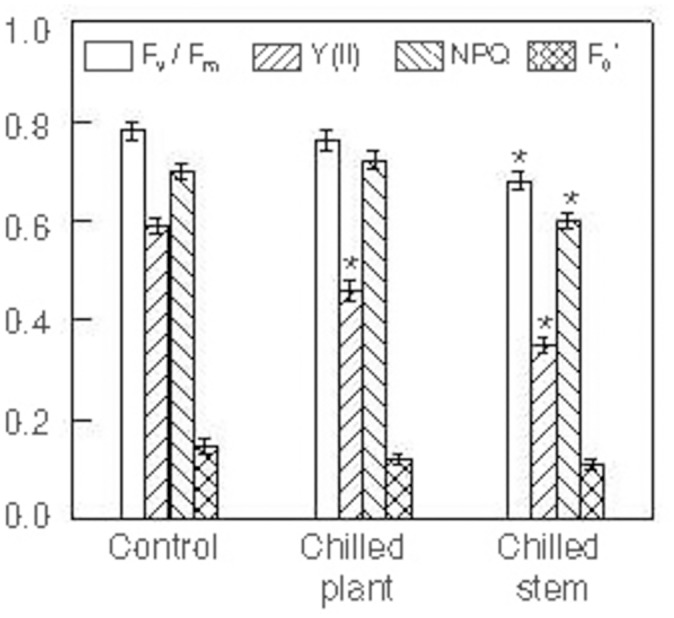 Fig 1
Fig 1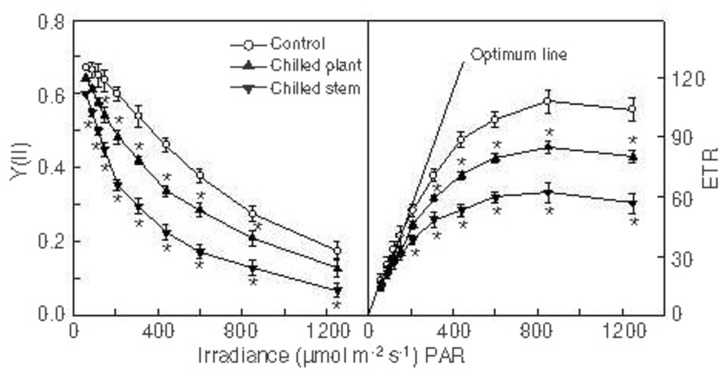 Fig 2
Fig 2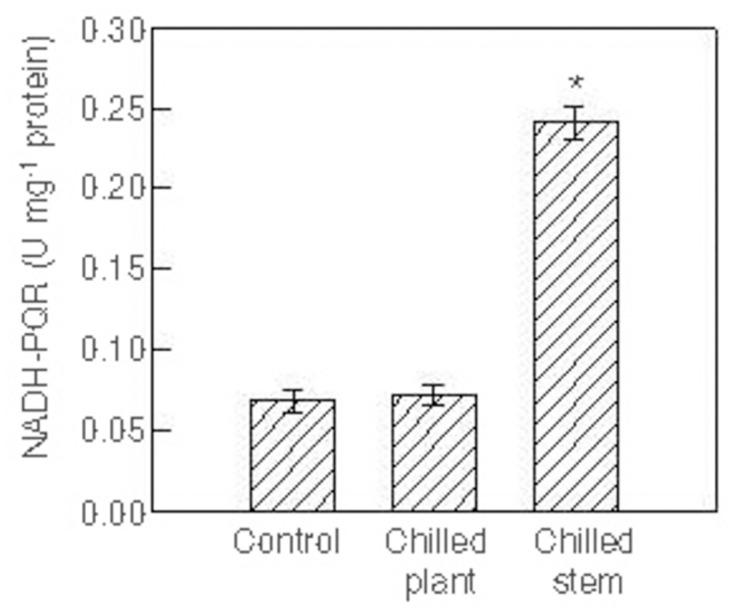 Fig 3
Fig 3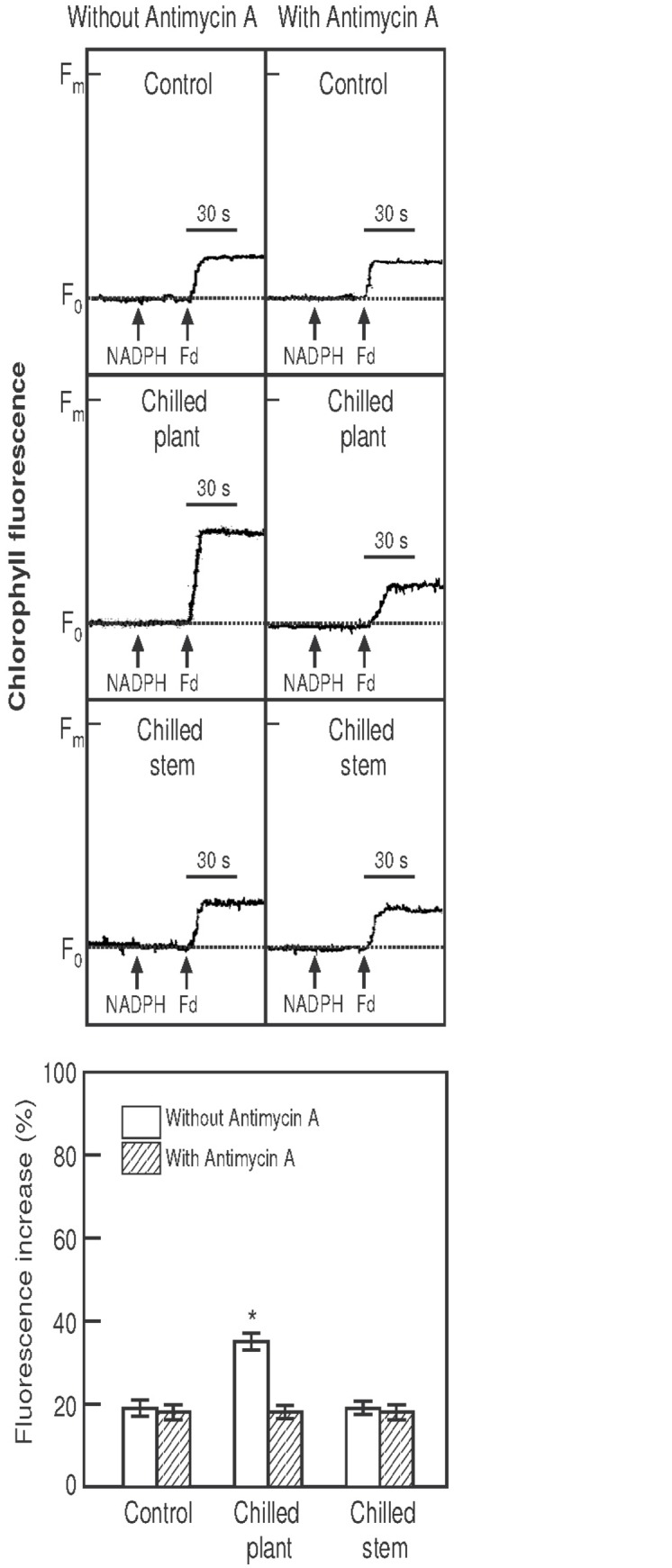 Fig 4
Fig 4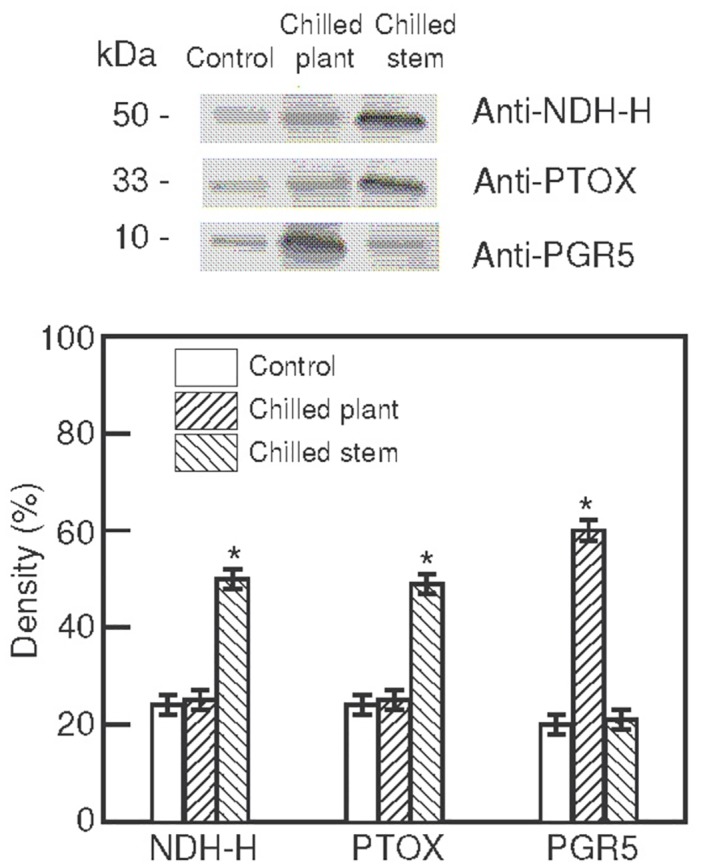 Fig 5
Fig 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGlobal Energy and Sustainability Research · Natural Resources and Economic Development
Introduction
Plants are frequently exposed to environmental stress, both in natural and horticultural conditions, and as a result growth may be affected. Understanding the physiological processes that underlie stress injury and the tolerance mechanisms of plants to environmental stress is of immense importance for both horticulture and the environment. Tolerance to stress results from integrated events occurring at all organization levels, from the anatomical and morphological to the cellular, biochemical and molecular levels. At the biochemical level, plants alter their metabolism in various ways to accommodate environmental stress, photosynthesis being one of these ways.
Hibiscus rosa-sinensis is a flowering plant species widely grown as an ornamental throughout the tropics, subtropics and also in temperate regions. Many plant species of tropical and subtropical origin manifest physiological dysfunctions when they are exposed to low, but non-freezing, temperatures of about 10–12°C [1,2]. Chilling injury under light has been attributed to the selective inhibition of the photosystems [3], PSII being the more sensitive to stress under light conditions, while PSI is more stable [3]. Until recently, relatively little attention has been paid to the effect of root temperature on the chilling response of plant leaves. Suzuki et al. [4,5] stated that root temperature is a very important factor for the response and susceptibility of leaves to chilling stress. In Oryza sativa visible damage to leaves exposed to light was observed when only leaves, but not roots, were chilled, whereas no visible damage was observed when both leaves and roots were chilled simultaneously [4]. Additionally, when only leaves, but not roots, were chilled PSII was photoinhibited, but not PSI [4]. These authors suggested that leaf chilling while root temperature is high blocks the electron transport between Q_A_ and Q_B_ in PSII, which causes the over-reduction of PSII in the light, although the biochemical basis of chilling stress has not been established [5].
Photosynthesis in chloroplasts involves a vectorial electron transfer from water in the lumen to NADP^+^ in the stroma, which is achieved by means of redox carriers. Besides this major pathway, based on functional measurements, alternative electron transfer pathways, involving non-photochemical reduction or the oxidation of plastoquinones at the expense of stromal electron donors or acceptors, have been proposed. These additional reactions cover two main concepts, one based on the cycling of electrons around PSI [6–8] and the other on chlororespiration [9,10], which consists of electron transfer reactions from stromal reductants to O_2_ through the plastoquinone pool [7,11,12]. Two thylakoidal enzymes, both of which are important in chlororespiration, have been molecularly characterised: the plastid-encoded NADH dehydrogenase (NDH) complex [13–18] and the nucleus-encoded plastid-localised terminal oxidase (PTOX) [19–21]. The NDH complex is an entry point for electrons into the photosynthetic electron-transport chain, involving the non-photochemical reduction of plastoquinones, and PTOX is a point of electron transfer from plastoquinol to molecular oxygen, resulting in the formation of water in the stroma and reducing the formation of reactive oxygen species (ROS) [11]. In addition to chlororespiration, the NDH complex is involved in the cyclic electron flow around PS I [22]. Two parallel cyclic pathways exist around PSI [22], one involving the NDH complex and the other sensitive to antimycin A, in which two proteins are essential components: the thylakoid membrane protein encoded by pgr5 gene (PGR5) and the thylakoid transmembrane protein (PGRL1), which interacts functionally and physically with PGR5 [23–25]. The physiological roles of the chloroplast electron pathways operating around PSI have been difficult to establish. Although these reactions probably do not play a major role during photosynthesis under optimal conditions [26–29], they probably contribute to the flexibility of the electron transfer reactions required to balance ATP/NADPH requirements when photosynthesis operates under a changing environment [12,30–35].
Several studies have proposed that chlororespiratory components may be involved in the response mechanisms of plants to environmental stress, such as drought, heat and high light [19,36–50]. However, the possible role of chlororespiratory components in conditions of cold stress remain unclear [12]. It was reported that PTOX levels increased in alpine plant species acclimated to high light and low temperature [44,51] and in cold-acclimated Arabidopsis thaliana plants [52]. In contrast, no increase in the NDH complex under cold stress was observed [52]. Recently, we described the involvement of the chlororespiratory pathway, together with another pathway involving PGR5 polypeptide, in the tolerance to high light intensity under chilling stress of the cold-acclimated Spathiphyllum wallisii, a shade plant [53]. However, the extent of any cooperation between these pathways remains unclear and needs further investigation.
The present work studies the effects of cold on photosynthesis, as well as the involvement in the chilling stress of chlororespiratory enzymes and ferredoxin-mediated cyclic electron flow, in illuminated plants of Hibiscus rosa-sinensis, an ornamental of tropical origin with growth temperatures of 15–25°C.
Materials and Methods
Plant material and treatments
Hibiscus rosa-sinensis plants were grown in soil in 500 mL pots at 22–25°C in a greenhouse under natural light conditions and controlled watering to avoid drought stress until flowering. For the cold treatments, adult plants were transferred to cultivation chambers, where they were exposed to two 18 h photoperiods (500 μmol·m^-2^·s^-1^ PPFD supplied by white light lamps of 100 W Flood OSRAM, Augsburg, Germany) and different temperature treatments. Each 18 h photoperiod was followed by a 6 h night-period. The temperature treatments were as follows: 24°C/24°C (control conditions); 10°C/10°C and 10°C/24°C, representing stem/root temperatures, respectively. To change the temperature of the root with respect to the stem, a thermostatic circulator (LKB, GmbH, Germany) with a coil around the pot was used. The soil temperature was measured with a digital stem thermometer (Herter Instruments, Barcelona, Spain). In each treatment, the photoperiod temperature was the same as during the night-period. The measurements were made after the second night-period. The experiments were replicated in four separate plants for each treatment.
Isolation of thylakoid membranes
Chloroplasts were isolated from leaves as described by Quiles and Cuello [13] using an extraction buffer (pH 7.6) containing 0.35 M sucrose, 25 mM Na-Hepes, 2 mM Na_2_-EDTA, 2 mM ascorbic acid, 4 mM dithiothreitol, 10 mM MgCl_2_ and 1 mM phenylmethylsulfonyl fluoride. As reported previously, a comparison of cytochrome c oxidase specific activity and the polypeptide profiles in mitochondrial and chloroplast fractions indicated that the chloroplast preparation was essentially mitochondrion-free [54]. The chloroplasts were washed twice and osmotically broken with 10 mM Tricine, 10 mM NaCl and 10 mM MgCl_2_ (pH 7.8) buffer as described previously [45]. The thylakoid membrane pellet was resuspended in a buffer (pH 7.5) containing 200 mM sorbitol, 130 mM KCl and 5 mM potassium phosphate at a chlorophyll concentration of 0.4 mg·mL^-1^, thus providing the suspension of the thylakoid membranes.
Chlorophyll fluorescence measurements
Chlorophyll fluorescence was measured in the thylakoid membrane suspension (50 μg Chl mL^-1^) using a PAM-210 chlorophyll fluorometer (Heinz Walz GmbH, Effeltrich, Germany) and chlorophyll fluorescence was imaged, using the MINI-version of the Imaging-PAM (Heinz Walz GmbH, Effeltrich, Germany) in entire leaves before dawn, at the same temperature in the cultivation chambers as applied in the treatments. Prior to the fluorescence measurements, the samples were dark-adapted, the thylakoid membrane suspensions for 30 min and the entire leaves for 6h (night-period). First F_0_, the minimal fluorescence yield, and Fm, maximal fluorescence yield, were measured in dark-adapted samples. F_0_ was measured at a low frequency of pulse modulated measuring light, while Fm was measured with the help of a saturation pulse. This was followed by exposure of the entire leaves to actinic light (200 μmol·m^-2^·s^-1^ PPFD), measuring the fluorescence yield (F) and the maximal fluorescence yield in illuminated samples (F’m). The maximal quantum yield of PS II (F_v_/F_m_ = (F_m_-F_0_)/F_m_), the effective PS II quantum yield of illuminated samples (Y(II) = (Fm’-F)/Fm’), non-photochemical quenching (NPQ = (F_m_- F_m_’)/F_m_’)) and minimal fluorescence yield of illuminated sample (F_0_’ = F_0_/ (F_v_/F_m_ + F_0_/F_m_’)) were automatically calculated by the ImagingWin software (Heinz Walz GmbH, Effeltrich, Germany). Then light response curves were made in the entire leaves using pre-programmed rapid light curves of the PAM fluorometer software (Heinz Walz GmbH, Effeltrich, Germany) by illuminating the entire leaves with actinic light of different intensities (60, 90, 120, 150, 210, 310, 440, 600, 850 and 1250 μmol m^-2^ s^-1^ PAR), with 2 min illumination periods at each intensity. After each illumination period a saturation pulse was applied to determine the quantum yield of PS II and the relative electron transport rate, both of which were calculated using the PAM fluorometer software (Heinz Walz GmbH, Effeltrich, Germany).
NADH dehydrogenase activity measurements
The NADH dehydrogenase activity of thylakoid membranes was determined by measuring NADH oxidation at 340 nm in a Perkin Elmer (Germany) spectrophotometer at 25°C, using decylplastoquinone (Sigma, USA) as electron acceptor. the NADH-plastoquinone oxidoreductase (NADH-PQR) activity was determined as described by Gamboa et al. [55]. One unit (U) of enzymatic activity is defined as the amount of enzyme preparation, which oxidized 1 μmol of substrate (NADH) per minute in the reaction conditions. The extinction coefficient of 6.22 mM^-1^·cm^-1^ at 340 nm was used to calculate the NADH oxidation rate.
Gel electrophoresis and immunoblot analysis
SDS-PAGE was carried out in a linear gradient of 10–20% polyacrylamide gel (2.5% bis-acrylamide), as previously described [13]. Protein samples were denatured by the addition of 3.5% (w/v) SDS and 5% (v/v) mercaptoethanol and heating for 10 min at 70–80°C before being subjected to SDS-PAGE. Prestained SDS-PAGE standards (Bio-Rad Laboratories, USA) were used for immunoblot analyses. After SDS-PAGE, the polypeptides were electroblotted from gels onto polyvinylidene difluoride membranes (Immobilon-P, Millipore, USA). Transfer and immuno-detection of the blotted protein was carried out as described previously [13]. The secondary antibody was conjugated to alkaline phosphatase (Sigma, USA). Controls in which the primary and/or secondary antibodies were omitted during incubation did not reveal any bands.
Other determinations
Protein was quantified using the method of Lowry et al. [56] after precipitation with 10% (w/v) trichloroacetic acid. Chlorophyll was determined by Lichtenthaler and Wellburn´s [57] method using 80% (v/v) acetone as solvent. Densitometric analysis and estimation of the polypeptide molecular masses were performed by ACTIB 1D digital image analyzer (Microptic, Barcelona, Spain). Analysis of variance (ANOVA) was used to compare the control and cold treatments.
Results
Photosynthetic parameters
The fluorescence imaging technique was used to test photosynthesis in intact leaves from plants exposed to two photoperiods with different temperatures in the stem and root (°C stem/°C root): 24°C/24°C (control), 10°C/10°C (chilled plant) and 10°C/24°C (chilled stem). Fig 1 shows the values of the maximal quantum yield of PS II (F_v_/F_m_), the effective PS II quantum yield (Y(II)), the non-photochemical quenching (NPQ) and minimal fluorescence yield of illuminated sample (F_0_’). The F_v_/F_m_ values in plants exposed to cold were lower than the values of control plants, although a significant difference was only observed in plants exposed to 10°C/24°C. The effective PSII quantum yield decreased significantly in the plants exposed to both cold treatments, the lowest values being seen in 10°C/24°C plants. The NPQ, which is an indicator of thermal dissipation [58], did not increase significantly in plants exposed to 10°C/10°C, but decreased significantly in plants exposed to 10°C/24°C.
Photosynthetic parameters.Maximal quantum yield of PS II (Fv/Fm), effective PS II quantum yield (Y(II)), non-photochemical quenching (NPQ) and minimal fluorescence yield of illuminated sample (F0’) in intact leaves from Hibiscus rosa-sinensis plants after exposure to two photoperiods (18 h, 500 μmol·m-2·s-1 PPFD), with different temperatures maintained in the stem and in the root (°C stem/°C root): 24°C/24°C (control); 10°C/10°C (chilled plant) and 10°C/24°C (chilled stem). Each photoperiod was followed by a 6 h night-period, maintaining the same temperatures as in the photoperiod. The values are means ± SE from four separate plants for each treatment, with duplicate measurements in each plant. Significant differences are indicated by an asterisk (ANOVA, p<0.05).
The light response curves for the quantum yield of PSII and the relative electron transport rate (ETR) in intact leaves from plants in control conditions and those exposed to the cold treatments are shown in Fig 2. The quantum yield of PSII decreased with increasing photonic flux density in all cases, because electrons accumulated on the PSII acceptor side and there was a relative increase in non-radiative energy dissipation. When light was not excessive, the PSII quantum yield maintained its maximum value and the relationship between the relative electron transport rate and light intensity was linear (optimum line). The relative electron transport rate fell below the values predicted by the optimum line when the PSII quantum yield decreased because the light became excessive. Eventually a saturated rate was reached, which represents the photosynthetic electron transport capacity. This capacity, as well as the PSII quantum yields and the relative electron transport rates, were lower in plants exposed to cold temperatures than in control plants, although the differences with respect to the control values were greater in plants exposed to 10°C/24°C (chilled stem) than in those exposed to 10°C/10°C (chilled plant).
Light response curves.Light response curves for the quantum yield of PS II (Y(II)) and the relative electron transport rate (ETR) in intact leaves from Hibiscus rosa-sinensis plants after exposure to two photoperiods (18 h, 500 μmol·m-2·s-1 PPFD), with different temperatures maintained in the stem and in the root (°C stem/°C root): 24°C/24°C (control); 10°C/10°C (chilled plant) and 10°C/24°C (chilled stem). Each photoperiod was followed by a 6 h night-period, maintaining the same temperatures as in the photoperiod. The values are means ± SE from four separate plants for each treatment, with duplicate measurements in each plant. Significant differences are indicated by an asterisk (ANOVA, p<0.05).
NADH-PQR activity and electron donation to plastoquinone in the thylakoid membranes
The NDH activity was assayed as NADH-plastoquinone oxidoreductase (NADH-PQR) activity in thylakoid membranes isolated from leaves of plants exposed to two photoperiods with different temperatures (Fig 3). The activity was similar in plants exposed to 24°C/24°C (control) and 10°C/10°C (chilled plant). However, in plants exposed to 10°C/24°C (chilled stem) the activity was approximately 3.5 times higher than in the control.
NADH-plastoquinone oxidoreductase activity.NADH-plastoquinone oxidoreductase (NADH-PQR) activity in the thylakoid membranes isolated from leaves of Hibiscus rosa-sinensis plants after exposure to two photoperiods (18 h, 500 μmol·m-2·s-1 PPFD), with different temperatures maintained in the stem and in the root (°C stem/°C root): 24°C/24°C (control); 10°C/10°C (chilled plant) and 10°C/24°C (chilled stem). Each photoperiod was followed by a 6 h night-period, maintaining the same temperatures as in the photoperiod. The values are means ± SE from four separate plants for each treatment, with duplicate measurements in each plant. Significant differences are indicated by an asterisk (ANOVA, p<0.05).
The activity of electron donation by NADPH and ferredoxin to plastoquinone was assayed as an increase in the chlorophyll fluorescence emitted during exposure to light of a very low intensity (1.0 μmol m^-2^ s^-1^) [45], at which intensity, the fluorescence level predominantly reflects the reduction of plastoquinone by electron transport from ferredoxin [23]. Chlorophyll fluorescence increased after the addition of NADPH (0.25 mM) and ferredoxin (5 μM) under weak measuring light (Fig 4) in thylakoid membrane suspensions (50 μg Chl mL-1) isolated from the chloroplasts of leaves recently detached from plants exposed to two photoperiods with different temperatures. Additionally, thylakoid suspensions were incubated with antimycin A (2 μM) for 2 min prior to the measurements. Control plants and plants exposed to 10°C/24°C (chilled stem) showed a similar increase in chlorophyll fluorescence. However, plants exposed to 10°C/10°C (chilled plant) showed a more pronounced increase in chlorophyll fluorescence (approximately 2 times the control level). Antimycin A inhibited the increase in chlorophyll fluorescence compared with measurements made without inhibitor, although its effect was slight in the control and the 10°C/24°C plants.
Fluorescence increase.Increases in chlorophyll fluorescence after the addition of NADPH (0.25 mM) 30 s into the run and ferredoxin (5 μM) 60 s into the run, under weak measuring light (1.0 μmol·m-2·s-1) in thylakoid membranes (50 μg Chl·mL-1) isolated from recently detached leaves of Hibiscus rosa-sinensis plants after exposure to two photoperiods (18 h, 500 μmol·m-2·s-1 PPFD), with different temperatures maintained in the stem and in the root (°C stem/°C root): 24°C/24°C (control); 10°C/10°C (chilled plant) and 10°C/24°C (chilled stem). Each photoperiod was followed by a 6 h night-period, maintaining the same temperatures as in the photoperiod. Additionally, thylakoid suspensions were incubated with antimycin A (2 μM) for 2 min prior to the measurements. The figure shows typical curves and the histograms of the means ± SE from four separate plants for each treatment, with duplicate measurements in each plant. Significant differences are indicated by an asterisk (ANOVA, p<0.05).
The results suggest that cyclic electron transport was stimulated in leaves only when the whole plant was chilled, while chlororespiration was stimulated in leaves when the roots were not chilled and kept at 24°C and the stem was cooled to 10°C.
Immunoblot analysis
Thylakoid membranes isolated from leaves of plants exposed to two photoperiods with different temperatures were used for immunoblot analyses, using specific antibodies against the NDH-H subunit of the thylakoidal NDH complex (anti-NDH-H), PTOX and PGR5. Fig 5 shows that the antibody anti-NDH-H clearly recognised only one polypeptide of 50 kDa, while the antibodies anti-PTOX and anti-PGR5 recognised others of 33 kDa and 10 kDa, respectively, in each thylakoid membrane sample. No additional polypeptide was recognised by the antibodies assayed (not shown). Immunoblots were analysed densitometrically and the results are also shown in Fig 5. The levels of PTOX and the NDH-H subunit of the NDH complex were higher in the thylakoids of plants exposed to 10°C/24°C (chilled stem) than in leaves from control plants. However, the levels in leaves from plants exposed to 10°C/10°C (chilled plant) remained similar to the control. In contrast, the PGR5 polypeptide levels only increased in plants exposed to 10°C/10°C and remained similar to the control in leaves from plants exposed to 10°C/24°C.
Immunoblot analysis.Immunoblot analysis of the thylakoid polypeptides isolated from leaves of Hibiscus rosa-sinensis plants after exposure to two photoperiods (18 h, 500 μmol·m-2·s-1 PPFD), with different temperatures maintained in the stem and in the root (°C stem/°C root): 24°C/24°C (control); 10°C/10°C (chilled plant) and 10°C/24°C (chilled stem). Each photoperiod was followed by a 6 h night-period, maintaining the same temperatures as in the photoperiod. Twenty micrograms per well of the thylakoid proteins were separated by SDS-PAGE prior to immunoblot assays and tested with anti-NDH-H, anti-PTOX, and anti-PGR5 antibodies. The molecular masses of the polypeptides are indicated in the margin. The immunoblots from three independent experiments were analysed by video densitometric analysis. In each case, the total density of the samples (control + chilled plant + chilled stem) was taken as 100%. The values are means ± SE. Significant differences are indicated by an asterisk (ANOVA, p<0.05).
Discussion
The tropical species Hibiscus rosa-sinensis is sensitive to chilling stress. Under cold conditions PSII was inhibited and photosynthetic linear electron transport was down-regulated, as indicated by a reduction in the photochemistry efficiency of PSII and in the electron transport capacity. However, the susceptibility of leaves to cold may be modified by the root temperature, we have seen in this work that when the roots were kept at 24°C while the stem was cooled to 10°C, the quantum yield of PSII and the relative electron transport rates were much lower than when the whole plant (root and stem) was exposed to 10°C. In rice seedlings Suzuki et al. [4,5] also observed that PSII activity was severely inhibited and that NPQ and photosynthetic electron transport decreased when shoots, but not roots, were cooled. These authors suggested [5] that the chilling of rice leaves accompanied by high root temperature blocked the electron transport between Q_A_ and Q_B_ in PSII, leading to the over-reduction of PSII in the light, the blockage being attributed to an interaction of a certain molecule with a component or components of PSII. Suzuki et al. [5] also suggested that this molecule was probably nitric oxide, which to have the potential to bind to the Q_A_-Fe^2+^-Q_B_ complex, thereby inhibiting electron transfer from Q_A_ ^-^ to Q_B_. Nitric oxide may be produced when nitrate is accumulated and, more recently, Suzuki et al. [59] described that the malfunction of photosynthesis induced by high root temperature is associated with the accumulation of nitrate and nitrite in leaves.
The inhibition of photosynthetic linear electron transport, the main electron transfer pathway, could trigger alternative pathways, such as cyclic electron flow around PSI and chlororespiration [12], since regulation of the flow of photosynthetic electrons is essential for the health and survival of plants. These alternative ways of photosynthetic linear electron transport may contribute to protecting the photosynthetic apparatus under stress conditions [12,24,35,43,45,46,48,52,55,60–62]. Recently, we described the involvement of the chlororespiratory pathway in the tolerance to high light intensity under chilling stress [53]. However, the extent of any cooperation between chlororespiration and alternative photosynthetic electron transport pathways, such as cyclic electron flow around PSI involving PGR5 polypeptide, was unclear [53]. In the present work, it was observed that the activity of electron donation by NADPH and ferredoxin to plastoquinone, which was inhibited by antimycin A, increased in relation to the control, only when the whole plant, root and stem, was exposed to 10°C. Additionally, the levels of PGR5 polypeptide, an essential component of cyclic electron flow sensitive to antimycin A [23], also increased in plants where both roots and stems were exposed to 10°C, suggesting that cyclic electron transport is stimulated only when the whole plant is chilled. However, temperatures of 10°C/24°C (stem/root) did not increase the activity of electron donation by NADPH and ferredoxin to plastoquinone, probably because, at these temperatures, a malfunction in electron transport somewhere between PSII and PSI there was, as suggested by Suzuki et al. [5]. This malfunction blocks both linear and cyclic electron flows, leading to the over-reduction of PSII in the light [5]. Interestingly, both NDH complex and PTOX increased only in plants exposed to 10°C/24°C (stem/root), suggesting that chororespiration was stimulated only when the roots were kept at 24°C and the stem was cooled to 10°C.
The concerted action of NDH complex and PTOX would optimize the efficiency of the cyclic pathways, preventing over-reduction of the electron transfer chain [12,63] and reducing the accumulation of reactive oxygen species by recycling electrons to the plastoquinone pool and, ultimately, to oxygen through PTOX, forming water in the stroma. Additionally, these pathways contribute to balancing ATP/NADPH requirements and to generating a large proton gradient and acidification of the lumen, which play a crucial role in the regulation, preventing the light-induced inactivation of both PSI and PSII through the formation of non-photochemical quenching [12,35].
In conclusion, this work has shown that illuminated plants were more tolerant to chilling when both stem and roots were chilled. In such conditions, neither NDH complex nor PTOX increased. However, both the activity of electron donation by NADPH and ferredoxin to plastoquinone and the amount of PGR5 polypeptide, an essential component of cyclic electron flow around PSI, increased, suggesting that in these conditions cyclic electron flow helps protect photosystems. However, when the stems but not the roots were cooled, cyclic electron flow did not increase and the efficiency of PSII photochemistry decreased considerably as a result of insufficient dissipation of the excess light energy. In such conditions, both chlororespiratory enzymes, NDH complex and PTOX, increased. The apparent correlation between the low functioning of PSII and the up-regulation of PTOX and the thylakoidal NDH complex supports a role for chlororespiration in the response of plants to chilling stress when other pathways, such as cyclic electron flow around PSI, are insufficient to protect PSII.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lyons JM (1973) Chilling injury in plants. Annu. Rev. Plant Physiol. 24: 445–466.
- 2Allen DJ, Ort DR (2001) Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends Plant Sci. 9: 36–42.10.1016/s 1360-1385(00)01808-211164376 · doi ↗ · pubmed ↗
- 3Sonoike K (1998) Photoinhibition of photosystem I in chilling sensitive plants determined in vivo and in vitro, in: Garab G. (Ed.), Photosynthesis: Mechanisms and Effects. Kluwer Academic Publishers, Dordrecht, The Netherlands, pp. 2217–2220.
- 4Suzuki K, Nagasuga K, Okada M (2008) The chilling injury induced by high root temperature in the leaves of rice seedlings. Plant Cell Physiol. 49: 433–442. 10.1093/pcp/pcn 020 18252732 · doi ↗ · pubmed ↗
- 5Suzuki K, Ohmori Y, Ratel E (2011) High root temperature blocks both linear and cyclic electron transport in the dark during chilling of the leaves of rice seedlings. Plant Cell Physiol. 52: 1697–1707. 10.1093/pcp/pcr 104 21803813 · doi ↗ · pubmed ↗
- 6Fork DC, Herbert SK (1993) Electron transport and photophosphorylation by Photosystem I in vivo in plants and cyanobacteria. Photosynth. Res. 36: 149–168. 10.1007/BF 00033035 24318920 · doi ↗ · pubmed ↗
- 7Bukhov N, Carpentier R (2004) Alternative photosystem I-driven electron transport routes: mechanisms and functions. Photosynth. Res. 82: 17–33. 1622861010.1023/B:PRES.0000040442.59311.72 · doi ↗ · pubmed ↗
- 8Johnson GN (2005) Cyclic electron transport in C 3 plants: fact or artefact? J. Exp. Bot. 56: 407–416. 1564731410.1093/jxb/eri 106 · doi ↗ · pubmed ↗