Metabolic state modulates risky foraging behavior via multiple branches of the insulin/IGF-1-like pathway in C. elegans
Kaiden H Price, Jason T Braco, Sofia Franco, Ansley N Varisco, Preeti F Sareen, Peter J Niesman, Hanna N Wetzel, Kelly E Crowe, Michael N Nitabach

TL;DR
This study shows how the metabolic state of C. elegans influences risky foraging behavior through the insulin/IGF-1 signaling pathway.
Contribution
The study identifies multiple branches of the insulin/IGF-1-like pathway involved in modulating foraging behavior in C. elegans.
Findings
DAF-2, the insulin/IGF-1 receptor, is necessary for modulating risky foraging behavior in response to food deprivation.
CASY-1, a calsyntenin ortholog, is required for food deprivation to influence foraging behavior.
DAF-16 is not essential for food-deprivation-driven changes in foraging behavior.
Abstract
Foraging to acquire nutrients is an essential and sometimes risky behavior displayed by nearly all animals. Appropriately balancing foraging risks with nutrient requirements is pivotal for peak survival and reproduction, and metabolic state (ie how urgently the animal requires nutrients) is a strong modulator of risky foraging behavior. In this study, we asked what molecular signal allows Caenorhabditis elegans to change its foraging behavior in response to changes in its metabolic state. We used an assay of risky foraging behavior, where wild type worms increase risky foraging behavior after food deprivation, to screen for candidate genes. We found that DAF-2, the singular receptor in the C. elegans insulin/IGF-1 signaling (IIS) pathway, is necessary for worms to modulate risky foraging behavior in response to short-term food deprivation. Worms with mutations in genes upstream and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
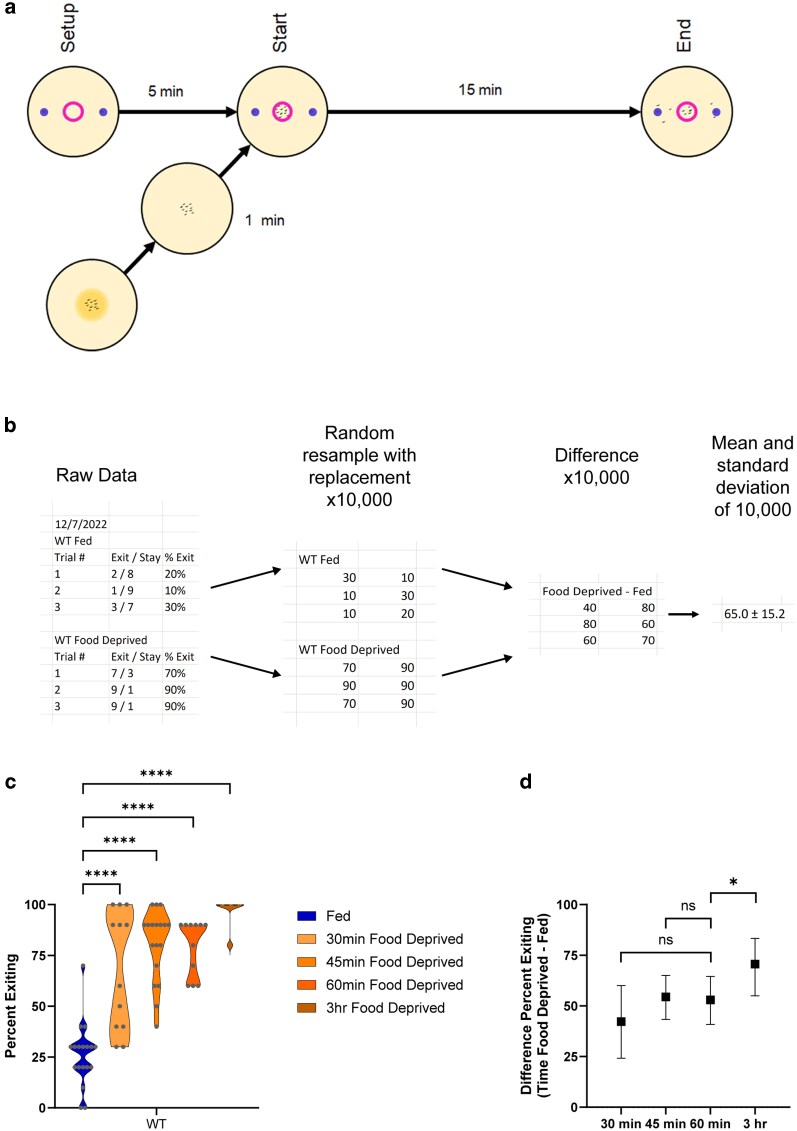 Fig. 1
Fig. 1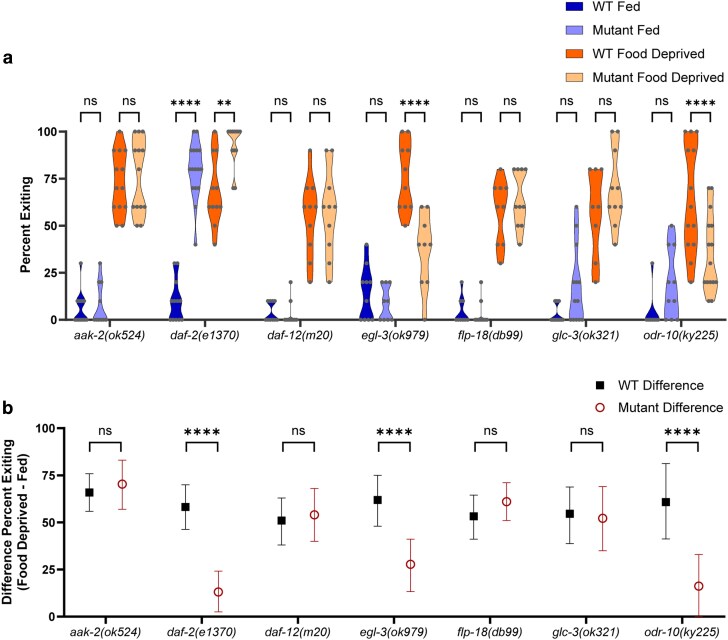 Fig. 2
Fig. 2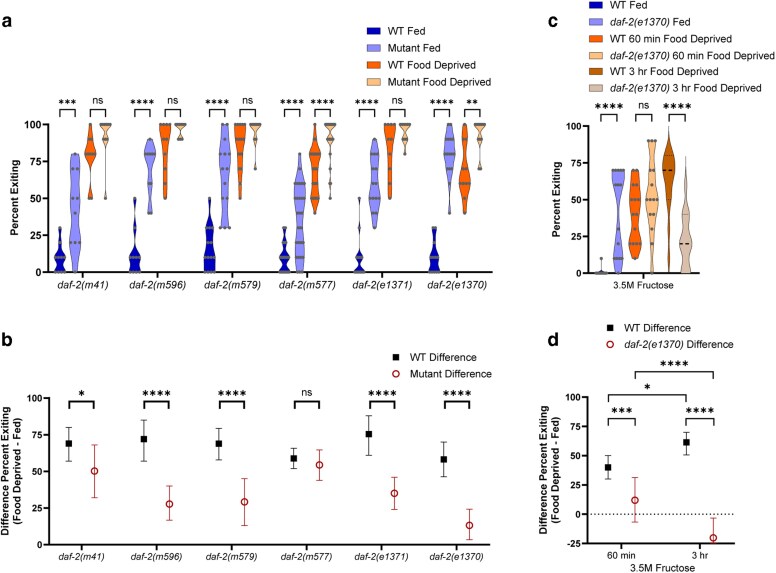 Fig. 3
Fig. 3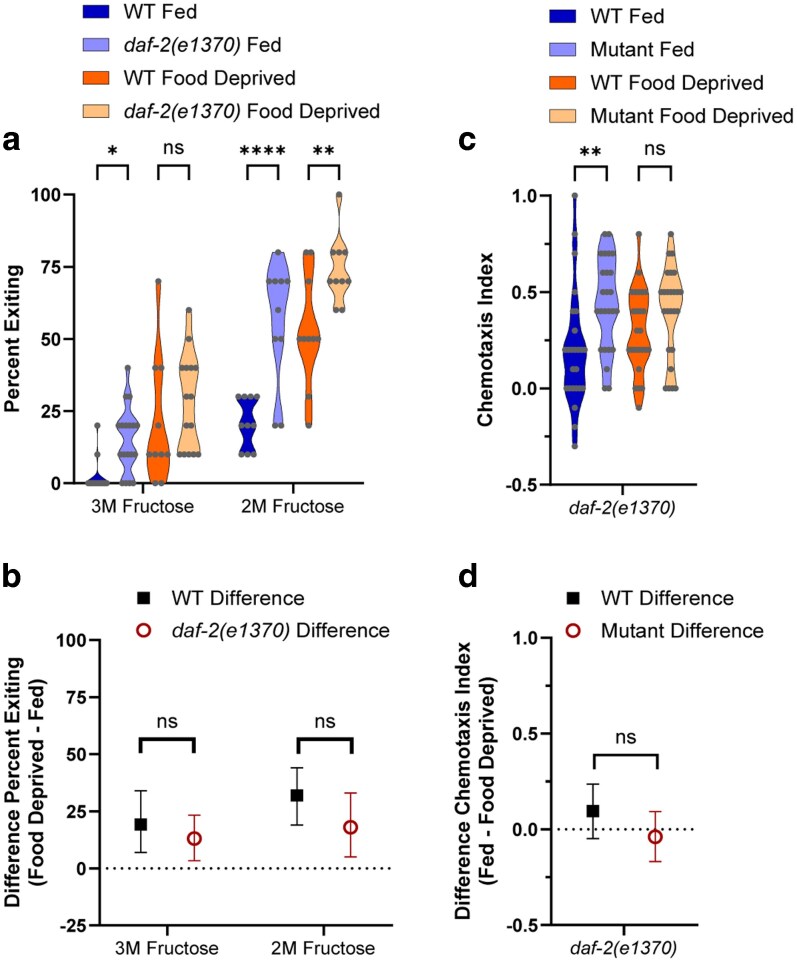 Fig. 4
Fig. 4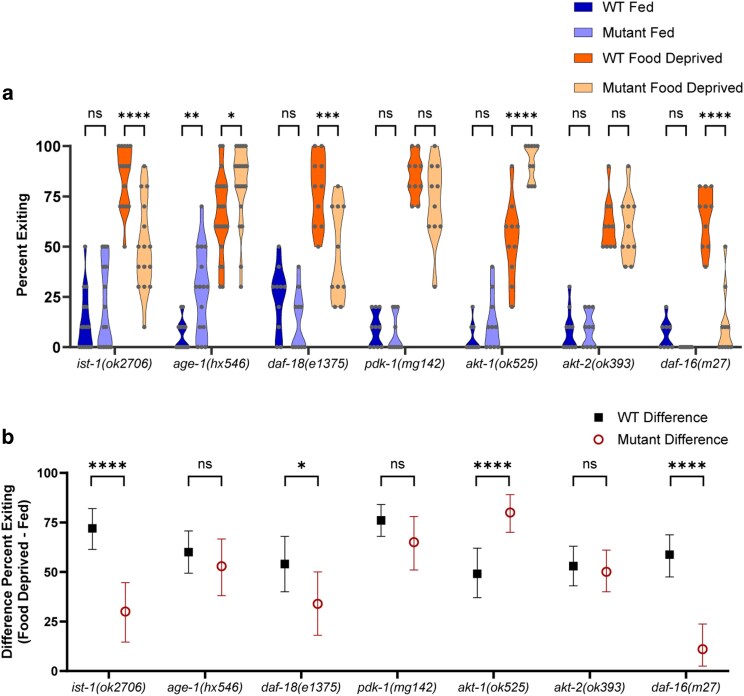 Fig. 5
Fig. 5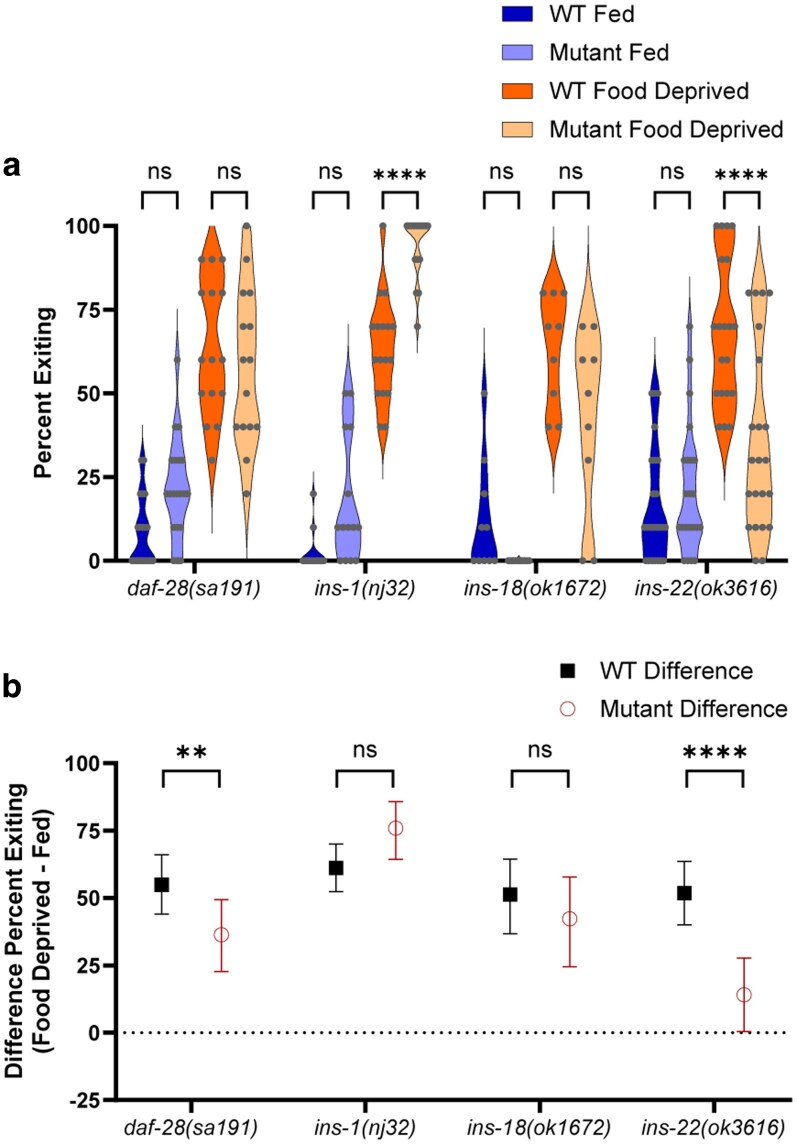 Fig. 6
Fig. 6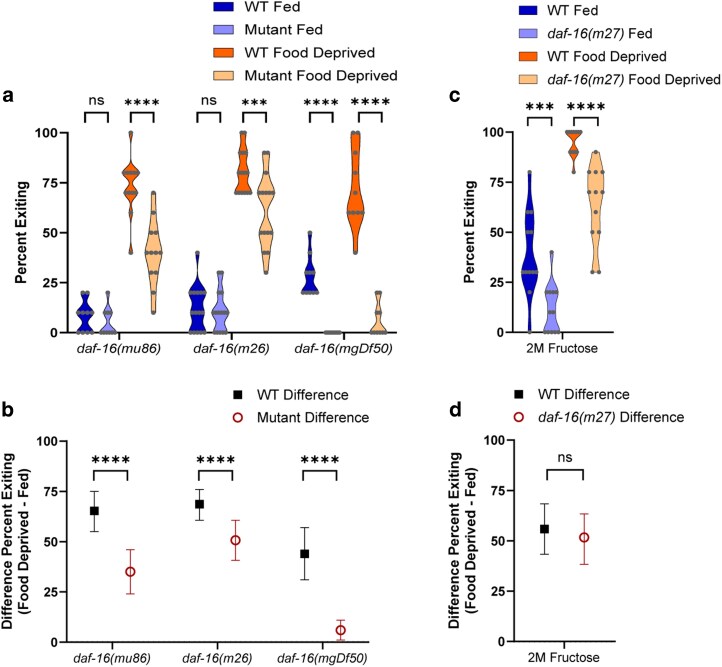 Fig. 7
Fig. 7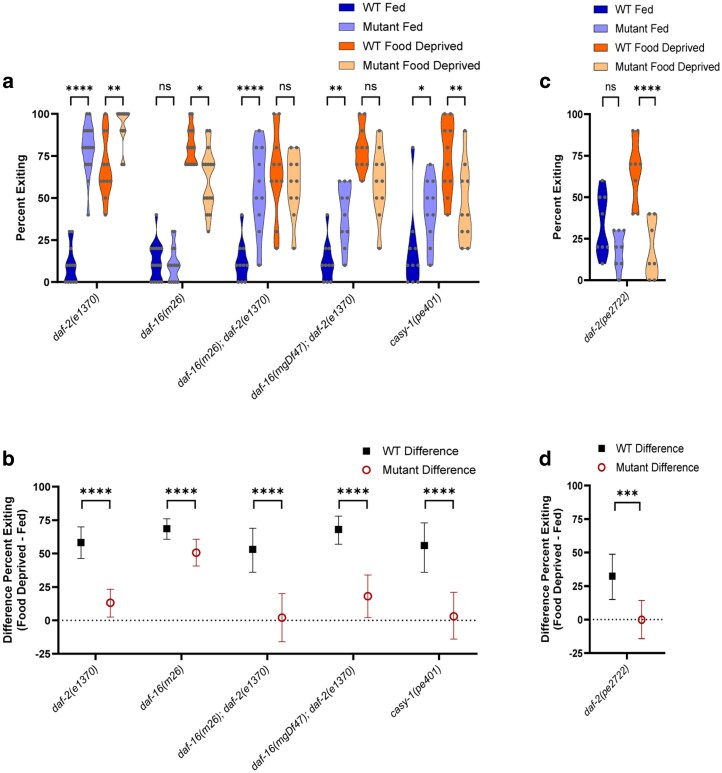 Fig. 8
Fig. 8| Figure | Type of ANOVA | Three-way interaction | Two-way interaction | Main effects |
|---|---|---|---|---|
| 2a | Three-way | Interaction of mutant genotype × fed condition × WT control, | ND | ND |
| 2b | Two-way | ND | Mutant genotype × WT control, | ND |
| 3a | Three-way | Mutant genotype × fed condition × WT control, | ND | ND |
| 3b | Two-way | ND | Mutant genotype × WT control, | ND |
| 3c | Two-way | ND | Genotype × fed condition, | ND |
| 3d | Two-way | ND | Genotype × duration of food deprivation, | genotype, |
| 4a | Three-way | Genotype × fed condition × fructose concentration, | Genotype × fed condition, | ND |
| 4b | Two-way | ND | Genotype × fructose concentration, | Fructose concentration, |
| 4c | Two-way | ND | Genotype × fed condition, | Genotype, |
| 5a | Three-way | Mutant genotype × fed condition × WT control, | ND | ND |
| 5b | Two-way | ND | Mutant genotype × WT control, | ND |
| 6a | Three-way | Mutant genotype × fed condition × WT control, | ND | ND |
| 6b | Two-way | ND | Mutant genotype × WT control, | ND |
| 7a | Three-way | Mutant genotype × fed condition × WT control, | Mutant genotype × fed condition, | ND |
| 7b | Two-way | ND | Mutant genotype × WT control, | ND |
| 7c | Two-way | ND | Genotype × fed condition, | genotype, |
| 8a | Three-way | Mutant genotype × fed condition × WT control, | Mutant genotype × fed condition, | ND |
| 8b | Two-way | ND | Mutant genotype × WT control, | ND |
| 8c | Two-way | ND | Mutant genotype × WT control, | ND |
- —National Institute of General Medical Sciences10.13039/100000057
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms · FOXO transcription factor regulation · Reproductive Biology and Fertility
Introduction
Almost all animals forage—they display distinct behavioral patterns to search for and acquire nutrients (Kuburich et al. 2016; Rudebeck and Izquierdo 2022; Zjacic and Scholz 2022). However, even for the simplest of organisms, foraging is a complex tradeoff between risk and reward. Typically, conservative foraging strategies (eg staying in one place) are less likely to result in nutrient acquisition; however, more risky foraging strategies cost energy and increase the likelihood that the animal will encounter environmental hazards, predation, and injury. Effectively balancing nutrient requirements with foraging risks requires ongoing assessment of the external environment (nutrient sources, predation risks, environmental hazards, etc) and the internal environment (metabolic state, nutrient deficiencies, developmental stage, etc). The animal must then integrate that information to drive appropriate behavioral choices (Lima and Dill 1990; Mukherjee and Heithaus 2013; Zjacic and Scholz 2022).
In humans, foraging behavior now occurs in the context of a broad array of human cultures and agricultural systems (Capocasa and Venier 2024), and human behaviors around seeking and obtaining food are commonly understood as dietary choice behaviors (Wadi et al. 2024). These dietary choices impact a wide variety of health outcomes, including body fat composition, cancer risk, heart health, and risk of dementia (Guasch-Ferré and Willett 2021; Clemente-Suárez et al. 2023). Body mass index (a flawed but frequently used measure that estimates body fat on the population level; Gonzalez et al. 2017; Bray 2023) has been found to be 30%–55% heritable (Bouchard 2021), suggesting that internal physiologic drivers may deeply influence human feeding behaviors. Additionally, surgical and pharmacological interventions have shown substantially greater effects on weight loss than behavioral interventions alone (Blüher et al. 2023), further suggesting that a significant part of human foraging behavior is driven by physiological signals (Rebello and Greenway 2016; Watts et al. 2022). Expanding our understanding of how foraging behavior is regulated at the molecular level may lead to better interventions that can alter human dietary choices in ways that improve human health.
We chose to study the regulation of foraging behavior in the microscopic nematode worm Caenorhabditis elegans. This worm is an excellent model system for investigating the neural, genetic, and molecular mechanisms of behavior. Substantial work has already been done to examine the regulation of feeding behavior in C. elegans, identifying relevant neurons (Gray et al. 2005; Ghosh et al. 2016; Rhoades et al. 2019), neurotransmitters (Flavell et al. 2013; Lee et al. 2017; Gadenne et al. 2022), and genes (Matty et al. 2022; McLachlan et al. 2022; Boor et al. 2024). However, the molecular mechanism by which C. elegans integrates metabolic state into foraging decisions is not yet fully understood.
We previously employed a multisensory decision assay that is well-suited for assessing foraging risk tolerance in C. elegans (Ghosh et al. 2016). This assay presents worms with a complex choice: cross a hyperosmolar barrier to approach a food odor, or avoid the hyperosmolar barrier and thus fail to approach the food odor (Ishihara et al. 2002; Shinkai et al. 2011). While well-fed worms overwhelmingly avoid both the hyperosmolar barrier and the food odor, worms that have been food deprived will generally cross the hyperosmolar barrier and move toward the food odor (Ishihara et al. 2002; Ghosh et al. 2016; Matty et al. 2022). Here, we used this foraging risk assay to examine which genes may be involved in allowing the worm to integrate its internal metabolic state (ie whether it is well-fed or food deprived) into its foraging behavior. We initially ran a mutant screen for candidate genes using established mutant worm lines. This candidate screen had 2 clear hits: egl-3 and daf-2. We chose to focus on daf-2, the insulin/IGF-1-like receptor in C. elegans.
The canonical insulin/IGF-1 signaling (IIS) pathway in C. elegans first activates or antagonizes the DAF-2 receptor with 1 or more of 40 ILPs (insulin-like peptides) (Zheng et al. 2018). The cell then transmits the signal from the DAF-2 receptor via a series of kinases, beginning with AGE-1, the ortholog of mammalian PI3K (Weinkove et al. 2006). AGE-1 phosphorylates PIP2 to increase concentrations of PIP3, which activates downstream kinases; DAF-18, a PTEN ortholog, opposes this activity, dephosphorylating PIP3 and decreasing activating of downstream kinases (Murphy 2013). The insulin receptor substrate protein, IST-1, facilitates AGE-1 activity, and may act to pair non-canonical effectors to DAF-2 receptor activity (Murphy 2013). Ultimately, the AKT kinases phosphorylate the transcription factor DAF-16 (orthologs to the mammalian FOXO) (Paradis and Ruvkun 1998). When phosphorylated, DAF-16 is sequestered in the cell soma, where it is unable to act as a transcription factor (Murphy 2013). We found that many of the genes in the IIS pathway in C. elegans are necessary for the worm to adjust its foraging behavior in a way that appropriately discriminates between a well-fed and a food-deprived metabolic state. Moreover, while previous research supports a role for daf-2 in modulating foraging behavior (Matty et al. 2022), our research further suggests that 2 downstream IIS pathway elements (the FOXO transcription factor DAF-16 and the calsyntenin ortholog CASY-1) are likely acting in 2 parallel pathways to modulate foraging risk tolerance.
Materials and methods
Strains and culturing
Worms were maintained at 20 °C on nematode growth media (NGM) media cultured with Escherichia coli OP50 as a food source. All strains were obtained from the Caenorhabditis Genetics Center. The Bristol N2 strain was used as wildtype (WT) in all assays. In all assays, worms were age-matched by picking L4 worms 18–24 h before assays. As many daf-2 strains form dauers at 20 °C (Gems et al. 1998), all daf-2 strains were maintained at 15 °C alongside paired WT control plates. For these strains, L4 worms were picked and moved to 20 °C 18–24 h before being used in assays. (In experiments where the mutant worms were maintained at 15 °C, the respective WT control worms were also maintained at 15 °C).
Foraging risk assay and osmotic avoidance assays
Multisensory foraging risk assays were conducted as in Ghosh et al. (2016). Plates were first prepared by taking fresh (no more than 5 d old) 60 mm NGM plates and drying the open plates in a closed chemical hood for 1 h. A 1 cm diameter ring was drawn in the middle of the dried NGM plate using 10 µl 3 M fructose (shown in pink in Fig. 1a). (1 experiment—the daf-2c(pe2722) mutant—used 3.3 M fructose; likely due to differing laboratory conditions, a higher fructose concentration was required to obtain equivalent WT foraging behavior.) Two 1 µl dots of 1:350 diacetyl were then placed on opposite sides of the plate, 1 cm from the ring border (shown in purple in Fig. 1a). The plate was then set to the side for 5 min to allow an odor gradient to form. Meanwhile, 10 worms were moved from the culture plate to a clean NGM plate with no E. coli and left for at least 1 min to allow any remaining bacteria to wash off onto the clean plate. At the end of the 5 min, the 10 worms were moved from the cleaning plate to the center of the assay plate inside the fructose ring. The worms were allowed to forage for 15 min, and then the number of worms remaining in the ring was counted. “Percent exiting” was determined as (10−(# worms remaining)) × 10. (Unisensory osmotic avoidance assays were performed in essentially the same manner as our foraging risk assay, except that the diacetyl was not added to the assay plate.)
*Assay and experimental design. a) A 1 cm diameter circle of 3 M fructose (represented by the small torus) was drawn in the center of a 60 mm Petri dish of nematode growth media (NGM). Two dots of 1:350 diacetyl were placed outside the fructose circle, 1 cm from the edge of the Petri dish (represented by the dots). The dish sat for 5 min to allow an odor gradient to form. Meanwhile, 10 worms were moved from a feeding dish to a clean NGM dish for 1 min to wash off any bacteria adhering to the worms. At the end of the 5-min set up, the 10 worms were moved to the center of the fructose circle. Worms were allowed to forage for 15 min, and then the number of worms remaining in the circle was counted. b) Raw data were additionally analyzed by calculating the bootstrapped distribution of the difference between the fed and food deprived groups. Raw datasets of fed and food deprived worms were first transformed into bootstrapped datasets with 10,000 replications using resampling with replacement (the first 2 of 10,000 replications are shown here). A third bootstrapped dataset was created by calculating the difference of each data point from the fed and food deprived bootstrapped datasets. This third bootstrapped dataset of the difference (food deprived minus fed) was used to calculate a population average, standard deviation, and 95% confidence interval for the effect of food deprivation. c) Foraging behavior of fed and food deprived wild type worms in the foraging risk assay with varying durations of food deprivation (analyzed by 1-way ANOVA with Dunnett's post hoc tests, using “Fed” as the control condition). d) Bootstrapped distributions (means and 95% confidence intervals) of the differences between fed and food deprived wild type worms from (c) (calculated as described in (b); analyzed by 1-way ANOVA with Dunnett's post hoc tests, using “60 min Food Deprived difference” as the control condition). Adjusted P-values of post hoc tests: ns, P ≥ 0.05; *, P < 0.05; ***, P < 0.0001.
Food deprivation
For all assays that used food deprivation, worms were moved from a culture plate to a cleaning plate (as above) for at least 1 min, and then placed into a food-deprivation plate. The food-deprivation plate was a clean, dried NGM plate with an ∼4 cm diameter ring-shaped acetate stencil pressed gently into the agar to form a seal. The stencil reduced the number of worms that crawl up the side of the petri dish and desiccate. Worms remained in the food-deprivation plate for 1 h (unless otherwise specified), and then were moved directly onto the assay plate.
Unisensory chemotaxis assay
Chemotaxis assays were performed as in Ghosh et al. (2016). Fresh 60 mm NGM plates were prepared with 1 µl of 1:10,000 diacetyl in dH2O placed 1 cm from 1 edge of the plate, and 1 µl of dH2O placed 1 cm from the opposite edge of the plate; an odor gradient was allowed to form for 5 min before 10 worms were placed in the center of the plate. Chemotaxis was measured after 15 min. Chemotaxis index was measured as (worms at odor−worms at control)/total worms.
Statistics
Given the design of our experiments, we analyzed data in 2 ways. First, the raw data was analyzed in GRAPHPAD Prism by 3- or 2-way ANOVA. (Best practices for analysis of 2-way and 3-way ANOVAs begin with the highest possible level of interaction and proceed through lower levels only if the higher levels do not reach significance. We display the tested significance results of all 2- and 3-way ANOVA interactions in Table 1; in this table, significance results are reported starting as the highest-order interaction and continuing through lower-order interactions and then main effects, if no interaction was significant.) The highest level significant ANOVA result was reported in the text of the results. When the ANOVA analysis found an effect of or interaction with mutant genotype, we followed up with post hoc tests using Šidák's correction that compared each mutant group (fed and food deprived) to the matched WT control animals tested on the same days. (Dunnett's post hoc tests were used in experiments that had a single WT control group.) This analysis answered the questions, “Do fed mutant animals exit more, less, or about the same as fed WT animals?” and “Do food deprived mutant animals exit more, less, or about the same as food deprived WT animals?” We present this raw data as violin graphs with each raw data point overlaid on the violin graph. For the visual representation of significance in all graphs, we use the following designations: ns, P ≥ 0.05; *, P < 0.05; **, P < 0.01; ***, P < 0.001; ****, P < 0.0001.
While the ANOVA results are useful for identifying whether or not an interaction exists between genotypes and feeding conditions within the larger dataset, our research is primarily interested in the question of, “Is the effect of food deprivation in this mutant larger, smaller, or about the same as in WT animals?” This specific question—particularly when applied to individual genotypes within a larger dataset—is not directly answered by classical 2- or 3-way ANOVA with post hoc tests (examples of this are discussed in our results, notably in reference to the behavior of age-1 and daf-28 mutant worms). Additionally, our datasets show a high degree of skew, which violates the assumption of normal distribution for ANOVA analysis; unfortunately, there are no nonparametric tests that are quite equivalent to 2- or 3-way ANOVA. Bootstrapping allows for more direct comparisons of the effect of food deprivation within individual genotypes, and transforming our raw data into bootstrapped difference datasets reduced skew, increasing the robustness of ANOVA and post hoc analysis of the bootstrapped difference datasets. We directly tested the effect of food deprivation (ie the difference in exiting between fed and food deprived animals in a single genotype) by creating a bootstrapped difference in python (similarly to the bootstrapping of learning integration index in Koemans et al. 2017a, 2017b). Bootstrapping involves repeated random resampling with replacement over many replicates (typically 10,000 replicates). Each replicate makes a new dataset with the same N as the original dataset, resampling to take a random datapoint from the raw dataset to fill each row in the new dataset; for instance, if the raw dataset is “5, 6, 4,” a random resample with replacement could be “4, 6, 6” or “5, 4, 6” (an additional example is shown in Fig. 1b). To create a bootstrapped difference, our python script (included as Supplementary Information) first created a bootstrapped distribution of each raw dataset. After creating 10,000 resampled datasets for each raw dataset, a “difference” dataset was created by calculating the food deprived percent exiting minus the fed percent exiting for 10,000 paired resampled datasets (example in Fig. 1b). (Where the sample sizes of the fed and food deprived groups were not equal, we used the smaller N to calculate the difference dataset.) This allowed us to then calculate an average and standard deviation for each of the 10,000 datasets, followed by calculating the mean of the averages and standard deviations. Additionally, we sorted the means of the 10,000 resampled datasets, then removed the 2.5% highest and 2.5% lowest resampled replicate means. We used the remaining maximum and minimum values as the bounds of the 95% confidence interval of the population mean of the effect of food deprivation.
These bootstrapped calculations are robust measures of the effect of food deprivation in each genotype, which then allow us to present targeted statistical and visual analysis. We used the mean average and standard deviation of the bootstrapped difference in GRAPHPAD Prism to test a 2-way ANOVA for each data group, followed by post hoc tests with Šidák's correction. (For the N of each bootstrapped difference in the statistical analysis, we used the total fed and food deprived N of the raw data minus 1, as N−k equals N−1 when there are 2 groups, in this case “Fed” and “Food Deprived”). In our figures, we present the bootstrapped mean average of the difference between fed and food deprived for each genotype and WT control group as a mean and 95% confidence interval.
Graphical Abstract
The graphical abstract for this publication was created with BioRender.com (Price 2025).
Results
Foraging risk assay
Our foraging risk assay tests the propensity for worms to exit a ring of an aversive substance, a hyperosmotic fructose solution absorbed into the surface of an agarose plate, in order to approach an attractive food-related odorant (diacetyl). We ran this assay in WT worms after varying lengths of food deprivation (Fig. 1c, 1-way ANOVA P < 0.001; and Fig. 1d, 1-way ANOVA P = 0.0003). We found that worms significantly increase their exiting after as little as 30 min food deprivation (Fig. 1c, P < 0.0001), and continue to exhibit increased exiting compared with fed worms through 3 h of food deprivation (Fig. 1c, P < 0.0001). When comparing the effects of food deprivation of various durations, there was no significant difference between 30 min or 45 min food deprivation when compared with 60 min food deprivation (Fig. 1d, respectively P = 0.1768 and P = 0.9882). However, worms that were food deprived for 3 h exhibited a significantly higher effect of food deprivation than worms that had been food deprived for 1 h (Fig. 1d, P = 0.0166).
Targeted screen
We first ran a targeted screen of candidate genes that previous literature suggested might transduce information about the worm's metabolic state to the neurons involved in making foraging risk decisions (Ao et al. 2004; Apfeld et al. 2004; Murakami et al. 2005; Rottiers and Antebi 2006; Chalasani et al. 2007; Cohen et al. 2009; Dallière et al. 2016). Of the 7 gene mutants we tested, 3 exhibited a strong reduction in the effect of food deprivation (Fig. 2a; 3-way ANOVA interaction of mutant genotype × fed condition × WT control, P < 0.0001, and Fig. 2b, 2-way ANOVA interaction of mutant genotype × WT control, P < 0.0001; detailed significance for all 2- and 3-way ANOVAs is reported in Table 1), while 4 genes—the AMP-activated protein kinase aak-2, the nuclear hormone receptor daf-12, the NPY-like peptide flp-18, and the glutamate-gated chloride channel *glc-3—*did not impact foraging risk behavior (Fig. 2a, b, P > 0.05).
*Targeted screen of candidate genes. a) Foraging behavior in the foraging risk assay of fed and food deprived mutant worms and respective WT controls (analyzed by 3-way ANOVA, followed by post hoc tests with Šidák's correction for WT vs mutant at each condition and genotype). b) Bootstrapped distributions (means and 95% confidence intervals) of the differences between fed and food deprived worms from (a) (analyzed by 2-way ANOVA, followed by post hoc tests with Šidák's correction for each genotype). Adjusted P-values of post hoc tests: ns, P ≥ 0.05; **, P < 0.01; ***, P < 0.0001.
One of the 3 positive results, the odr-10 mutant, was an internal positive control that behaved as expected. Our lab has previously shown that food deprived WT animals show an increase in exiting a fructose ring both under unisensory conditions (fructose alone) and in the multisensory foraging risk assay (wherein diacetyl is placed outside of the fructose ring); however, the magnitude of the change in exiting is much greater in the foraging risk assay (Ghosh et al. 2016). Therefore, we expect a mutant animal that lacks the ability to sense the diacetyl odorant to behave similarly to WT animals in a unisensory assay of fructose aversion: we expect it will show a mild increase in exiting when food deprived, but less than the WT animals that can still sense the diacetyl (Ghosh et al. 2016). ODR-10 is an olfactory receptor necessary for sensing the diacetyl odorant used in the multisensory foraging risk assay (Sengupta et al. 1996). odr-10 mutants in the multisensory foraging risk assay display lower exiting than WT animals when food deprived (Fig. 2a, P < 0.0001) as well as a reduced effect of food deprivation when compared with WT animals (Fig. 2b, P < 0.0001). Effectively, the worms do not sense the attractive odorant, and so they do not display the odor-driven enhancement in exiting behavior seen by food deprived worms with normal chemosensory ability.
The proprotein convertase egl-3 mutant exhibits foraging behavior that, on a surface level, looks similar to the behavior of the odr-10 mutant (Fig. 2a). However, in view of the literature, changes in foraging behavior in the egl-3 mutant are more likely to be due to alterations in responses to food deprivation than to alterations in sensory capacity (Kass et al. 2001; Larsch et al. 2015; Thapliyal et al. 2023). The egl-3(ok979) mutant worms have near-normal GCaMP responses to pulses of diacetyl, with perhaps a slight enhancement of habituation and desensitization (Larsch et al. 2015), and the egl-3(nu349) mutant has typical responses to hyperosmolarity (Kass et al. 2001). Moreover, it was previously found that egl-3 mutation reduced a starvation-induced change in backwards movements and tail bending, a microbehavior, which may contribute to the changes we see in the foraging risk assay (Thapliyal et al. 2023). Thus, it may be more likely that egl-3 functions in a hunger signaling pathway, such that loss of the gene results in worms that take lower foraging risks (reduced exiting when food deprived, Fig. 2a; P < 0.0001) and exhibit reduced effects of food deprivation (Fig. 2b; P < 0.0001).
In contrast to egl-3, the daf-2 IIS receptor gene appears to function as a satiation signal, such that loss of daf-2 results in worms that exhibit high percent exiting regardless of metabolic state (Fig. 2a; higher exiting when well-fed P < 0.0001, higher exiting when food deprived P = 0.0065), with a strong reduction in the effect of food deprivation (Fig. 2b; P < 0.0001). DAF-2 is the sole IIS transmembrane receptor in C. elegans, and has long been known to play roles in various types of learning where worms avoid an environmental condition associated with food deprivation, such as high salt or altered ambient temperature (Murakami et al. 2005; Kodama et al. 2006; Tomioka et al. 2006; Vellai et al. 2006). Many genes in the IIS pathway in C. elegans are orthologous to insulin pathway genes in mammals, suggesting conserved effects of these genes that may translate into mammalian research (Hua et al. 2003). Moreover, it is possible that EGL-3 exerts its influence on foraging behavior via modulation of DAF-2 activity. EGL-3 has been found to process several ILPs which act on the DAF-2 receptor, including INS-3, INS-4, and INS-6 (Hung et al. 2014; Zheng et al. 2018). We thus focused further on the role of the IIS pathway in integrating metabolic state with foraging risk tolerance in the worm.
Validation of the IIS receptor daf-2
We performed a series of experiments testing for allele-specific effects of daf-2 and further validating it as a relevant gene in this behavior. First, we tested 5 additional daf-2 IIS receptor mutant alleles in the foraging risk assay (Fig. 3a, 3-way ANOVA interaction of mutant genotype × fed condition × WT control, P = 0.0003; and Fig. 3b, 2-way ANOVA interaction of mutant genotype × WT control, P < 0.0001). All of these additional mutant alleles exhibited an increase in exiting when fed (Fig. 3a; daf-2(m41) P = 0.0001, daf-2(m596) P < 0.0001, daf-2(m579) P < 0.0001, daf-2(m577) P < 0.0001, and daf-2(e1371) P < 0.0001), and all but one exhibited a reduced effect of food deprivation (Fig. 3b; daf-2(m41) P = 0.0338, daf-2(m596) P < 0.0001, daf-2(m579) P < 0.0001, daf-2(m577) P = 0.8501, and daf-2(e1371) P < 0.0001). daf-2 mutant alleles are known to be divisible into 2 classes: Class I alleles have a more restricted phenotype that only impacts dauer arrest, longevity, and thermotolerance, while Class II alleles have more widespread phenotypic effects, including altered body morphology and brood size, as well as sometimes arresting at embryonic and L1 life stages. It has been hypothesized that Class I alleles may only reduce the ability of daf-2 to transduce signals to 1 major downstream effector pathway, while Class II alleles may also impact a second downstream effector pathway (Gems et al. 1998). We found that Class II alleles had overall stronger impacts than Class I alleles in the foraging risk assay. The 2 alleles which exhibit effects of food deprivation most similar to WT—m41 exhibiting only a small reduction in the effect of food deprivation (Fig. 3a, P = 0.0001), and m577 exhibiting no reduction in the effect of food deprivation (Fig. 3b, P = 0.8501)—are both Class I alleles, while the Class II alleles (m596, m579, and e1370) all exhibited strong effects. This is consistent with the possibility that multiple downstream daf-2 effector pathways may be involved.
*The daf-2 mutant alleles. a and b) Foraging behavior of worms in the foraging risk assay with different daf-2 mutant alleles (daf-2(e1370) data here is the same as that presented in Fig. 2, shown here for comparison). c and d) Foraging behavior of daf-2 mutant worms in the foraging risk assay at increased (3.5 M) fructose concentration (using both 60 min and 3 h food deprivation). a and c) Show raw data of fed and food deprived mutant worms and respective WT controls (analyzed by 3-way ANOVA, followed by post hoc tests with Šidák's correction for WT vs mutant at each condition and genotype); b and d) show the bootstrapped distributions (means and 95% confidence intervals) of the differences between fed and food deprived worms from each respective raw data figure (analyzed by 2-way ANOVA, followed by post hoc tests with Šidák's correction for each genotype). Adjusted P-values of post hoc tests: ns, P ≥ 0.05; *, P < 0.05; **, P < 0.01; ***, P < 0.0001.
We next tested the daf-2 IIS receptor mutant for an effect of food deprivation in a higher-risk version of the foraging risk assay (Fig. 3c, 2-way ANOVA interaction of genotype × fed condition, P < 0.0001; and Fig. 3d, 2-way ANOVA effect of genotype, P < 0.0001; effect of duration of food deprivation, P = 0.0273). When a mutant worm shows overall very high exiting behavior, as most daf-2 mutants do (Fig. 3a, b), the high baseline behavior may mask an effect of food deprivation. The higher-risk foraging risk assay lowers the percent of worms exiting the circle by increasing the fructose concentration (from a standard 3 M to 3.5 M) and hence increasing the aversion of worms to exiting. Lowering the overall percent of worms exiting the circle thus allows better observation of any effect of food deprivation. We additionally performed this assay both at our typical (60 min) and a longer (3 h) duration of food deprivation. In the higher-risk foraging assay, daf-2 mutant worms maintained their increased exiting compared with WT when well fed (Fig. 3c, fed WT vs fed daf-2 mutant, P < 0.0001) but did not display substantially higher exiting than WT when food deprived for 1 h (Fig. 3c, P = 0.2728). At 3 h food deprivation, daf-2(e1370) mutant worms displayed significantly lower exiting than WT worms (Fig. 3c, P < 0.0001). Further, we saw that WT worms exhibited greater effect of food deprivation than daf-2 mutant worms in the 3.5 M fructose multisensory foraging risk assay at both 60 min and 3 h lengths of food deprivation (Fig. 3d, P = 0.0005 and P < 0.0001, respectively). While WT worms after 3 h food-deprivation exhibited a stronger effect of food deprivation than after 60 min food deprivation (Fig. 3d, P = 0.0127), daf-2 mutant worms after 3 h food deprivation exhibited a reduced effect of food deprivation compared with the effect found after 60 min food deprivation (Fig. 3d, P < 0.0001). While we do not currently fully understand why the daf-2 mutant showed reduced exiting after 3 h food deprivation, it is possible it is due in part to complex behavioral changes that occur during starvation, such as an increase in intermittent sleep episodes (Skora et al. 2018). Moreover, a reduced exiting phenotype of daf-2 mutant worms at a longer period of food deprivation is in line with previous literature (Matty et al. 2022). This is unlikely to be due to any sort of reduction in the ability of daf-2 mutant worms to survive starvation, as the daf-2 mutation is well-known to promote starvation survival (Baugh and Hu 2020). Together, these results support the conclusion that the higher exiting of fed daf-2 mutant worms is due to the mutant worms being unable to appropriately distinguish between a fed and 60 min food-deprived metabolic state.
Next, we tested daf-2 IIS receptor mutant animals in unisensory assays to test whether sensory deficits may be driving the smaller effect of food deprivation. As worms in unisensory assays exhibit a much lower effect of food deprivation compared with their behavior in the multisensory foraging risk assay, a lack of ability to sense either hyperosmolarity or diacetyl odor could produce results similar to the results we would expect from a loss of ability to distinguish between fed and food deprived conditions. However, our results indicate that daf-2 mutant worms are capable of sensing both hyperosmolarity and the diacetyl odorant (consistent with Matsuura et al. 2007). We tested both WT and daf-2 mutant animals in a unisensory assay of hyperosmolar avoidance at concentrations of both 3 and 2 M fructose (Fig. 4a, 3-way ANOVA interaction of genotype × fed condition, P = 0.1031; genotype × fructose concentration, P = 0.0045; fed condition × fructose concentration, P = 0.1518; and Fig. 4b, 2-way ANOVA effect of fructose concentration, P = 0.0342; effect of genotype, P = 0.0158). As has been shown previously (Ghosh et al. 2016), WT animals showed higher exiting of the unisensory fructose ring when food deprived (Fig. 4a, WT fed vs WT food deprived, P = 0.0099), although this increase was not as large as that of WT animals in the multisensory foraging risk assay (eg compare WT animals in Fig. 4a to WT animals in Fig. 3a). While daf-2 mutants continued to exhibit higher exiting than WT animals when well-fed, consistent with a “perpetually food deprived” metabolic state (Fig. 4a, 3 M fructose fed WT vs fed daf-2 mutant, P = 0.0397; 2 M fructose fed WT vs fed daf-2 mutant, P < 0.0001), they also exhibited higher exiting at 2 M fructose than at 3 M fructose, indicating functional hyperosmolar sensation (Fig. 4a, daf-2 mutant fed 3 M vs daf-2 mutant fed 2 M, P < 0.0001; daf-2 mutant food deprived 3 M vs daf-2 mutant food deprived 2 M, P < 0.0001). These results, alongside previous literature (Matsuura et al. 2007), suggest that the loss of an effect of food deprivation in the daf-2 mutant worm in the foraging risk assay is not caused by a simple failure of the daf-2 mutant worm to detect hyperosmolarity.
*The daf-2 unisensory assays. a and b) Unisensory hyperosmotic avoidance measured as exiting of the fructose ring by daf-2 mutants at typical (3 M) and reduced (2 M) fructose concentration. c and d) Chemotaxis of daf-2 mutants to 1 µl of 1:10,000 diacetyl. a and c) show raw data of fed and food deprived mutant worms and respective WT controls (analyzed by 3-way ANOVA, followed by post hoc tests with Šidák's correction for WT vs mutant at each condition and genotype); b and d) show the bootstrapped distributions (means and 95% confidence intervals) of the differences between fed and food deprived worms from each respective raw data figure (analyzed by 2-way ANOVA, followed by post hoc tests with Šidák's correction for each genotype). Adjusted P-values of post hoc tests: ns, P ≥ 0.05; *, P < 0.05; **, P < 0.01; ***, P < 0.0001.
We also tested both WT and daf-2 IIS receptor mutant animals in a unisensory assay of chemotaxis to the diacetyl odorant (Fig. 4c, 2-way ANOVA effect of genotype, P = 0.0011; effect of fed condition, P = 0.5771; and Fig. 4d, unpaired t-test P = 0.0507). WT animals did not show a significant change in diacetyl attraction when food deprived (Fig. 4c, P = 0.4448). We found that daf-2 mutant animals overall exhibited a strong chemotaxis response, while also exhibiting a significant increase in chemotaxis compared with WT animals when well-fed (Fig. 4c, WT fed vs daf-2 mutant fed, P = 0.0024). This data is both consistent with the hypothesis that the altered foraging behavior of daf-2 mutant worms is likely not due to a simple deficit in chemosensation, and consistent with previous literature finding no deficits in daf-2(e1370) chemotaxis (Kauffman et al. 2010).
Genes of the canonical IIS pathway
We next built on these findings by investigating components of the IIS pathway that are downstream of the daf-2 IIS receptor (Fig. 5a, 3-way ANOVA interaction of mutant genotype × fed condition × WT control, P < 0.0001; and Fig. 5b, 2-way ANOVA interaction of mutant genotype × WT control, P < 0.0001). Overall, given that our results in daf-2 mutants indicate that activation of the IIS pathway acts as a satiety signal, we expect that loss-of-function mutants of positive elements of the IIS pathway—ist-1, age-1, akt-1, and akt-2—will exhibit higher levels of exiting in the foraging risk assay (ie will behave as if they are more hungry), and loss-of-function mutants of negative elements of the IIS pathway—daf-18 and *daf-16—*as well as the gain-of-function mutant of the positive element *pdk-1(mg142)—*will exhibit lower levels of exiting (ie will behave as if they are less hungry).
*Insulin/IGF-1 signaling pathway mutants. a) Foraging behavior in the foraging risk assay of fed and food deprived mutant worms and respective WT controls (analyzed by 3-way ANOVA, followed by post hoc tests with Šidák's correction for WT vs mutant at each condition and genotype). b) Bootstrapped distributions (means and 95% confidence intervals) of the differences between fed and food deprived worms from (a) (analyzed by 2-way ANOVA, followed by post hoc tests with Šidák's correction for each genotype). Adjusted P-values of post hoc tests: ns, P ≥ 0.05; *, P < 0.05; **, P < 0.01; ***, P < 0.001; ***, P < 0.0001.
Contrary to our expectations, the ist-1 mutant exhibited overall lower exiting when food deprived (Fig. 5a, P < 0.0001) with a greatly reduced effect of food deprivation (Fig. 5b, P < 0.0001). This lower exiting when food deprived could be due to IST-1 acting via non-canonical IIS pathways, rather than simply as a positive element in the canonical IIS pathway. As we predicted given the canonical IIS pathway, age-1 mutants exhibited higher exiting than WT worms, both when fed and when food deprived (Fig. 5a, fed WT vs fed age-1 mutant P = 0.0031, food deprived WT vs food deprived age-1 mutant P = 0.0330). However, age-1 mutants exhibited a similar effect of food deprivation compared with WT worms (Fig. 5b, P = 0.6621). As age-1(hx546) mutants have reduced AGE-1 activity, not a complete loss of function (Morris et al. 1996), it is possible that the age-1(hx546) mutants retained sufficient AGE-1 function to still respond to changes in metabolic state. Alternatively, these results could suggest that while the effect of AGE-1 activity is to lower high-risk foraging activities overall, AGE-1 activity may not be required for metabolic state to alter the worm's behavior.
The behavior of age-1 mutant worms additionally illustrates 1 scenario where including bootstrapped effect data allow for more complex analysis than traditional statistical tests alone. Initially, the ANOVA and post hoc analysis of the raw data in Fig. 5a simply allowed us to determine that there was an interaction between conditions (3-way ANOVA interaction of mutant genotype × fed condition × WT control, P < 0.0001), and that age-1 worms showed higher exiting than WT worms under both fed and food deprived conditions (WT vs fed age-1 mutant P = 0.0031, food deprived WT vs food deprived age-1 mutant P = 0.0330). However, these results are ambiguous when it comes to the difference of the effect of food deprivation; ie how much does food deprivation change exiting in the mutant, and is that the same or different from how much food deprivation changes exiting in the WT worm? Conducting ANOVA and post hoc analysis on the bootstrapped dataset allowed us to determine that, despite differences between the raw exiting in both fed and food deprived groups, the effect of food deprivation on exiting in age-1 mutants did not differ from that of WT worms (Fig. 5b, P = 0.6621). Traditional ANOVA statistical tests with direct post hoc comparisons of raw data will not differentiate between data such as our age-1 mutant worm data (where changes in overall exiting do not impact the effect of food deprivation), and data where a difference in overall exiting does impact the effect of food deprivation (such as the behavior of daf-2 receptor mutant worms in Fig. 2a, b).
The remaining kinases and phosphatases that we tested in the IIS pathway also exhibited results that were overall consistent with our hypothesis of IIS pathway activation acting as a satiety signal. The daf-18 (PTEN) mutant exhibited reduced exiting when food deprived (Fig. 5a, P = 0.0003), as well as a reduction in the effect of food deprivation (Fig. 5b, P = 0.0179). While akt-1 mutant worms exhibited the expected increased exiting when food deprived (Fig. 5a, P < 0.0001), they were the only mutant worm we tested that exhibited a significantly increased effect of food deprivation compared with WT worms (Fig. 5b, P < 0.0001). This may suggest the involvement of a non-canonical pathway, as the worm is able to allow metabolic state to modulate its foraging behavior without the influence of akt-1. By contrast, the akt-2 mutant worm behaved similarly to WT (Fig. 5a, b, P > 0.05). These results are likely at least in part because AKT-1 and AKT-2 have some degree of redundancy in worms (Paradis and Ruvkun 1998); additionally, it is possible that akt-1 plays a larger role than akt-2 in neurons involved in modulating foraging behavior, as a significant subset of neurons express akt-1 but not akt-2 (Taylor et al. 2021). Finally, daf-16 FOXO transcription factor mutant worms exhibited reduced exiting when food deprived compared with WT worms (Fig. 5a, P < 0.0001) and a reduced effect of food deprivation (Fig. 5b, P < 0.0001). Together, these results support the canonical IIS pathway, from the receptor daf-2 to the transcription factor daf-16, as a necessary modulator of foraging risk in C. elegans.
We additionally tested several ILP (insulin-like peptide) mutants (Fig. 6a, 3-way ANOVA interaction of mutant genotype × fed condition × WT control, P = 0.0005; and Fig. 6b, 2-way ANOVA interaction of mutant genotype × WT control, P < 0.0001). C. elegans has 40 ILPs, which can have wide variation in their expression (Ritter et al. 2013). Some, like INS-1 and INS-7, act as antagonists, rather than agonists, of the DAF-2 receptor (Pierce et al. 2001; Chen et al. 2013). As our hypothesis is that activation of the IIS pathway acts as a satiety signal (to reduce high-risk foraging behavior, and hence reduce exiting in the foraging risk assay), we expected that mutants of ILPs that act as agonists would exhibit overall increased exiting, and mutants of ILPs that act as antagonists would exhibit overall reduced exiting.
*Insulin-like peptide mutants. a) Foraging behavior in the foraging risk assay of fed and food deprived mutant worms and respective WT controls (analyzed by 3-way ANOVA, followed by post hoc tests with Šidák's correction for WT vs mutant at each condition and genotype). b) Bootstrapped distributions (means and 95% confidence intervals) of the differences between fed and food deprived worms from (a) (analyzed by 2-way ANOVA, followed by post hoc tests with Šidák's correction for each genotype). Adjusted P-values of post hoc tests: ns, P ≥ 0.05; *, P < 0.05; **, P < 0.01; ***, P < 0.001; ***, P < 0.0001.
We found that a daf-28 ILP mutant, which is characterized as an agonist of the IIS pathway (Zheng et al. 2018), did not differ significantly from WT animals in either fed or food deprived conditions (Fig. 6a, P > 0.05). However, the daf-28 mutant did exhibit a reduction in the effect of food deprivation (Fig. 6b, P = 0.0076). Notably, the behavior of daf-28 mutants is an excellent example wherein small differences in the opposite direction in both fed and food-deprived groups result in no direct difference between the paired groups following classical ANOVA and post hoc analysis, yet the bootstrapped analysis of the effect of food-deprivation reveals an overall change. Slightly lower food deprived exiting and slightly higher fed exiting are not significant differences alone, but together they add up to a significantly smaller effect of food deprivation when bootstrapped effect analysis is conducted.
By contrast, ins-1 mutant worms exhibited an increase in exiting when food deprived (Fig. 6a, P < 0.0001), but did not differ significantly from WT in the effect of food deprivation (Fig. 6b, P = 0.0848). This mixed effect is consistent with previous work on ins-1, which has shown that ins-1 interaction with the IIS pathway is complex; it may act as an agonist or antagonist of the IIS pathway, depending on the context (Pierce et al. 2001; Kodama et al. 2006; Tomioka et al. 2006; Zheng et al. 2018). ins-18 mutant worms behaved similarly to WT worms both when fed and food deprived (Fig. 6a, P > 0.05) and in the effect of food deprivation (Fig. 6b, P = 0.7083). Finally, the ins-22 mutant worm (which Zheng et al. 2018 found to be an antagonist) exhibited both a reduction in exiting when food deprived (Fig. 6a, P < 0.0001), as well as a strong reduction in the effect of food deprivation (Fig. 6b, P < 0.0001). These results further support the IIS pathway as a modulator of risky foraging behavior and suggest complexity in the ILPs that may transmit signals about metabolic state. The ILP ins-22 is clearly necessary for the appropriate increase in exiting when food deprived, but other ILPs, such as daf-28 and ins-1, may also modulate risky foraging behavior in more subtle ways.
Validation of the FOXO transcription factor daf-16
For further validation of daf-16 (the FOXO transcription factor primarily regulated by the IIS pathway) in transducing metabolic state to foraging behavior, we tested 3 more daf-16 mutant alleles (Fig. 7a, 3-way ANOVA interaction of mutant genotype × fed condition, P < 0.0001; mutant genotype × WT control, P < 0.0001; fed condition × WT control, P < 0.0001; and Fig. 7b, 2-way ANOVA interaction of mutant genotype × WT control, P = 0.0102). All of these daf-16 mutants exhibited a decrease in exiting when fed (Fig. 7a; daf-16(mu86) P < 0.0001, daf-16(m26) P = 0.0003, daf-16(mgDf50) P < 0.0001), and all exhibited a reduced effect of food deprivation (Fig. 7b; daf-16(mu86) P < 0.0001, daf-16(m26) P = 0.0001, daf-16(mgDf50) P < 0.0001).
*The daf-16 mutant alleles. a and b) Foraging behavior in the foraging risk assay of worms with different daf-16 mutant alleles. c and d) Foraging behavior of daf-16 mutant worms in the foraging risk assay at reduced (2 M) fructose concentration. a and c) show raw data of fed and food deprived mutant worms and respective WT controls (analyzed by 3-way (a) or 2-way (c) ANOVA, followed by post hoc tests with Šidák's correction for WT vs mutant at each condition and genotype); b and d) show the bootstrapped distributions (means and 95% confidence intervals) of the differences between fed and food deprived worms from each respective raw data figure (b, analyzed by 2-way ANOVA followed by post hoc tests with Šidák's correction for each genotype; d, analyzed by Student's t-test). Adjusted P-values of post hoc tests: ns, P ≥ 0.05; ***, P < 0.001; ***, P < 0.0001.
We next tested the daf-16 FOXO transcription factor mutant for an effect of food deprivation in a lower-risk version of the foraging risk assay (Fig. 7c, 2-way ANOVA effect of genotype, P < 0.0001; effect of fed condition, P < 0.0001; and Fig. 7d, Student's t-test, P = 0.4956). When a mutant worm shows overall very low exiting behavior, as most daf-16 mutants do (Fig. 7a, b), the low baseline behavior may mask an effect of food deprivation. The lower-risk foraging risk assay increases the percent of worms exiting the circle by decreasing the fructose concentration (from a standard 3 to 2 M) and hence decreasing the aversion of worms to exiting. Raising the overall percent of worms exiting the circle thus allows better observation of any effect of food deprivation. At the lower 2 M fructose concentration, daf-16 mutant worms maintained their overall lower exiting when food deprived (Fig. 7c, P < 0.0001) but now also exhibited lower exiting when well fed (Fig. 7c, P = 0.0003), resulting in a similar effect of food deprivation compared with WT animals (Fig. 7d, Student's t-test, P = 0.4956). This suggests that, while DAF-16 transcription factor activity increases exiting and thus high-risk foraging behaviors, daf-16 (unlike the IIS receptor daf-2) is not necessary for the worm to distinguish between a fed and food deprived metabolic state. These results further highlight the possibility that the DAF-2 receptor-mediated effect of food deprivation on exiting may be transduced via a non-canonical pathway (ie not solely via modulation of DAF-16 transcription factor activity).
Tests of a non-canonical pathway
To further test the hypothesis that the IIS receptor DAF-2 acts via a non-canonical pathway to modulate foraging risk in response to food deprivation, we tested double mutants of daf-2 and daf-16 (Fig. 8a, 3-way ANOVA effect of interaction of mutant genotype × fed condition, P < 0.0001; interaction of mutant genotype × WT control, P < 0.0001; interaction of fed condition × WT control, P < 0.0001; and Fig. 8b, 2-way ANOVA effect of interaction of mutant genotype × WT control, P = 0.0002). If DAF-2 acts solely via the canonical pathway that culminates by inhibiting DAF-16 transcriptional activity, then we would expect a daf-2; daf-16 double mutant to have a behavioral phenotype very similar to the daf-16 single mutant (eg reduced exiting, especially when food deprived). However, we found that 2 independent daf-2; daf-16 double mutants both exhibited a phenotype that was more similar to the daf-2 IIS receptor single mutant than the daf-16 FOXO transcription factor single mutant. daf-2; daf-16 double mutant worms exhibited increased exiting when fed, but lacked a large or significant decrease in exiting when food deprived (Fig. 8a, daf-16(m26); daf-2(e1370) fed vs WT fed P < 0.0001; daf-16(mgDf47); daf-2(e1370) fed vs WT fed P = 0.0021; daf-16(m26); daf-2(e1370) food deprived vs WT food deprived P = 0.9892; daf-16(mgDf47); daf-2(e1370) food deprived vs WT food deprived P = 0.0668). The daf-2; daf-16 double mutant worms additionally exhibited a strong reduction in the effect of food deprivation (Fig. 8b, daf-16(m26); daf-2(e1370) vs WT P < 0.0001; daf-16(mgDf47); daf-2(e1370) vs WT P < 0.0001).
*Mutants testing a non-canonical daf-2 pathway. a and b) Foraging behavior in the foraging risk assay of daf-16; daf-2 double mutant worms and casy-1 mutant worms. (Single mutants daf-2(e1370) and daf-16(m26) are the same data as shown in Figs. 2a and 5a, respectively, shown here for comparison.) c and d) Foraging behavior in the foraging risk assay of daf-2c mutant worms. a and c) show raw data of fed and food deprived mutant worms and respective WT controls (analyzed by 3-way (a) or 2-way (c) ANOVA, followed by post hoc tests with Šidák's correction for WT vs mutant at each condition and genotype); b and d) show the bootstrapped distributions (means and 95% confidence intervals) of the differences between fed and food deprived worms from each respective raw data figure (b, analyzed by 2-way ANOVA followed by post hoc tests with Šidák's correction for each genotype; d, analyzed by Student's t-test). Adjusted P-values of post hoc tests: ns, P ≥ 0.05; *, P < 0.05; **, P < 0.01; ***, P < 0.001; ***, P < 0.0001.
We then tested casy-1 as a potential mediator of a non-canonical daf-2 pathway in the foraging risk assay. The daf-2 IIS receptor gene in worms is expressed primarily as 2 isoforms, daf-2a and daf-2c. These isoforms differ only in that daf-2c includes the ligand binding domain exon 11.5 (Ohno et al. 2014). Previous research has found that inclusion of exon 11.5 allows DAF-2c to bind to CASY-1, which then binds to a kinesin-1 complex. DAF-2c is trafficked to the axon following food deprivation, where it may mediate synaptic activity; its presence is necessary for salt avoidance learning following food deprivation (Ohno et al. 2014). casy-1 mutants have been previously found to show defects in learning (Hoerndli et al. 2009) and sensory integration (Ikeda et al. 2008), while displaying typical naïve chemotaxis and hyperosmotic avoidance (Ikeda et al. 2008; Hoerndli et al. 2009). In our foraging risk assay, casy-1 mutant worms exhibited both increased exiting when fed (Fig. 8a, P = 0.0134) and decreased exiting when food deprived (Fig. 8a, P = 0.0034), leading to the strongest reduction in the effect of food deprivation that we observed in any single mutant, with a near-zero effect of food deprivation on exiting (Fig. 8b, casy-1 effect of food deprivation vs WT effect of food deprivation, P < 0.0001). We additionally tested a mutant worm, which lacks specifically the DAF-2c specific exon 11.5 (Nagashima et al. 2019), thus making it a daf-2c null mutant (Fig. 8c, 2-way ANOVA effect of mutant genotype × fed condition, P = 0.0124; and Fig. 8d, unpaired t-test P = 0.0002). In our foraging risk assay, the daf-2c mutant worms exhibited decreased exiting when food deprived (Fig. 8c, P < 0.0001) and showed a strong reduction in the effect of food deprivation (Fig. 8d, P = 0.0002). Altogether, these results further support the role of a non-canonical daf-2 pathway in mediating the effects of food deprivation on high-risk foraging behaviors and suggest a new avenue of research regarding the exact mechanisms by which a CASY-1 mediated DAF-2c signal may modulate behavior.
Discussion
Together, these results suggest a complex, 2-pronged pathway by which the IIS receptor may mediate foraging risk behaviors in response to food deprivation. When metabolic state is satiated (worms are well-fed), overall expression of agonistic ILPs is high (and expression of antagonistic ILPs is low) (Chen and Baugh 2014). High levels of agonistic ILPs leads to activation of the IIS receptor DAF-2, which inhibits the FOXO transcription factor DAF-16 and thereby reduces high-risk foraging behaviors. Additionally, when worms are well-fed, the DAF-2c isoform of the IIS receptor is primarily localized to the cell body (Ohno et al. 2014), which reduces DAF-2c activity in axons and also reduces high-risk foraging behaviors. When food deprivation alters metabolic state, IIS activation is reduced. Food deprivation then increases CASY-1 trafficking of DAF-2c to axons (Ohno et al. 2014), which increases high-risk foraging behaviors. Additionally, the disinhibition of DAF-16 transcriptional activity leads to an increase in exiting and further strengthens the worm's response to food deprivation.
We consider it an intriguing possibility that the canonical and non-canonical DAF-2 pathways may in part function to modulate the worm's response to food deprivation via different time scales. Multiple papers have reported effects of starvation that differ depending on the length of starvation (Murakami et al. 2005; Ghosh et al. 2016; Skora et al. 2018), and previous observations from our lab and others have demonstrated changes in foraging strategy in as little in 30 min (Hills et al. 2004; Ghosh et al. 2016). However, changes in cellular localization of the FOXO transcription factor DAF-16 due to food deprivation are generally thought to be slow, with most studies observing an effect after several hours to a day or more (Henderson and Johnson 2001; Weinkove et al. 2006), and no studies that we could find demonstrating a change in cellular localization in less than 60 min. Notably, 1 study of daf-16::GFP cellular localization observed nuclear localization after 75 min of food deprivation, while they did not see clear nuclear localization after 60 min of food deprivation (Henderson and Johnson 2001). Another study (Nagashima et al. 2019) found DAF-16::GFP localized to the nucleus of the ASER neuron following 1 h of food deprivation on a high-salt plate; the methods note that worms were imaged 10–60 min after incubation, suggesting an effective food deprivation time of 70–120 min. However, even assuming a sub-60-min time for DAF-16 to localize to the nucleus, it is unlikely that behavioral effects downstream of DAF-16 would occur immediately. Transcription factor mediated effects on behavior require time following nuclear translocation, as genes that may function as downstream mediators must be transcribed, and usually also translated, before altering the worm's behavior.
In contrast to the longer timescales of transcription factor activity, the speed of fast axonal transport and the length of C. elegans axons suggest proteins can be transported from the soma to synapses in 15–30 min (Wu et al. 2007, 2024; Niwa 2017; Feng et al. 2018; Lipton et al. 2018), and activation of kinase pathways present at the synapse likely occurs in seconds to minutes (Murphy et al. 1994; Blazek et al. 2015). Thus, it is possible that the DAF-2c/CASY-1 pathway is an early modulator of foraging risk following short-term food deprivation (ie in the 15–60 min time range), while the DAF-2/DAF-16 pathway is a mechanism that may be slower to act but may also be more strongly maintained over time. In support of this time-separated hypothesis, the food-deprivation-driven salt learning seen in Tomioka et al. 2006 (which used 1 h of food deprivation) required the daf-2 IIS receptor but did not seem to be affected by loss of the FOXO transcription factor daf-16. A 2-pronged foraging modulation strategy over multiple time scales may be especially relevant for an organism like C. elegans that has a boom-and-bust population growth strategy. For these worms, very low-risk foraging is ideal during the “boom” phase when food is abundant, but a strongly risk-tolerant foraging strategy that is both timely and sustained is imperative for animals to disperse and some to eventually find more food during the “bust” phase, when a food source has been depleted (Frézal and Félix 2015).
These differences in time scale may in part explain some of the differences between our results, and the results obtained in a recent paper that also used a multisensory foraging risk assay (Matty et al. 2022). Matty et al. (2022) made important contributions to our understanding of foraging regulation in the worm, notably identifying HLH-30 and MML-1 as transcription factors that modulate foraging risk behavior and likely modulate expression of ILPs. Similarly to our assay, they separated worms from a food-related (diacetyl) odor using an aversive barrier (copper sulfate) and counted animals that crossed the barrier vs animals that did not cross the barrier. Notably, both copper and high-osmolarity fructose are primarily sensed by the ASH neuron (Hilliard et al. 2005); the neural circuits for both assays are likely to be similar. However, while we used an assay test time of 15 min and saw changes with as little as 30 min of food deprivation, their assay used a test time of 45 min and a typical food deprivation duration of 3 h (Matty et al. 2022). As worms are not fed during the assay test, and the time at which we see alterations in behavior is (including the assay test time) 45 min past the initiation of food deprivation, this means that “well-fed” worms in a 45 min test assay are likely to behave as if they are slightly food deprived (solely by any short-term mechanisms) by the end of the assay. Using the 45 min test, they did not find a change in foraging behavior at 1 h food deprivation, but did see a difference at 2 h (Matty et al. 2022). Thus, we argue that our foraging risk assay using a 15-min test period and 30- to 60-min of food deprivation is primarily testing short-term mechanisms of food-deprivation-modulated risky foraging behavior. By contrast, their foraging risk assay using a 45-min test period may be testing long-term mechanisms of food-deprivation-modulated risky foraging behavior, which may not begin until 2–3 h after the initiation of food deprivation.
These results suggest areas of further research into how the insulin signaling pathway in the brain may modulate mammalian food-seeking behaviors and dietary choices. Strong evidence suggests that neuronal insulin is an important signaling pathway. Insulin in the mammalian CNS is known to play a role in feeding behavior (Baldini and Phelan 2019), metabolic diseases (Porte et al. 2005), Alzheimer's disease and other types of neurodegeneration (Kellar and Craft 2020; Alberini 2023), and learning and memory (Spinelli et al. 2019). The isoform of the IIS receptor trafficked by CASY-1 (DAF-2c) has similarities to the mammalian insulin receptor B isoform (Ohno et al. 2014). While the insulin receptor B isoform is not found in mammalian neurons (Kenner et al. 1995), previous research has shown that the insulin receptor can localize to rat synapses (Abbott et al. 1999). It is not yet fully clear in which cells and under which circumstances the mammalian insulin receptor may be trafficked to synapses, which cellular pathways may be modulated at those synapses, and how synaptic effects of the insulin receptor may differ in timescale and physiological effect from actions of the insulin receptor in the cell soma or nucleus. Our research, however, suggests the possibility that an improved understanding of these questions may provide insights into the neural control of mammalian foraging behavior.
Supplementary Material
jkag042_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abbott M-A, Wells DG, Fallon JR. 1999. The insulin receptor tyrosine kinase substrate p 58/53 and the insulin receptor are components of CNS synapses. J Neurosci. 19:7300–7308. 10.1523/JNEUROSCI.19-17-07300.1999.10460236 PMC 6782521 · doi ↗ · pubmed ↗
- 2Alberini CM . 2023. IGF 2 in memory, neurodevelopmental disorders, and neurodegenerative diseases. Trends Neurosci. 46:488–502. 10.1016/j.tins.2023.03.007.37031050 PMC 10192130 · doi ↗ · pubmed ↗
- 3Ao W, Gaudet J, Kent WJ, Muttumu S, Mango SE. 2004. Environmentally induced foregut remodeling by PHA-4/Fox A and DAF-12/NHR. Science. 305:1743–1746. 10.1126/science.1102216.15375261 · doi ↗ · pubmed ↗
- 4Apfeld J, O’Connor G, Mc Donagh T, Di Stefano PS, Curtis R. 2004. The AMP-activated protein kinase AAK-2 links energy levels and insulin-like signals to lifespan in C. elegans. Genes Dev. 18:3004–3009. 10.1101/gad.1255404.15574588 PMC 535911 · doi ↗ · pubmed ↗
- 5Baldini G, Phelan KD. 2019. The melanocortin pathway and control of appetite-progress and therapeutic implications. J Endocrinol. 241:R 1–R 33. 10.1530/JOE-18-0596.30812013 PMC 6500576 · doi ↗ · pubmed ↗
- 6Baugh LR, Hu PJ. 2020. Starvation responses throughout the Caenorhabditis elegans life cycle. Genetics. 216:837–878. 10.1534/genetics.120.303565.33268389 PMC 7768255 · doi ↗ · pubmed ↗
- 7Blazek M, Silva Santisteban T, Zengerle R, Meier M. 2015. Analysis of fast protein phosphorylation kinetics in single cells on a microfluidic chip. Lab Chip. 15:726–734. 10.1039/C 4LC 00797 B.25428717 · doi ↗ · pubmed ↗
- 8Blüher M et al 2023. New insights into the treatment of obesity. Diabetes Obes Metab. 25:2058–2072. 10.1111/dom.15077.37055715 · doi ↗ · pubmed ↗