The RNA-binding protein NOVA-1 regulates circRNA expression, alternative splicing, and aging in Caenorhabditis elegans
Emmanuel Adeyemi, Hussam Z Alshareef, Jaffar M Bhat, Pedro Miura, Alexander M van der Linden

TL;DR
This study shows that the protein NOVA-1 controls circular RNA production and splicing in worms, and its absence extends lifespan and improves stress recovery.
Contribution
The study identifies NOVA-1 as a key regulator of circRNA biogenesis and alternative splicing in C. elegans, linking it to aging and stress resilience.
Findings
NOVA-1 regulates 686 circRNAs in C. elegans, acting as a negative regulator for a subset.
NOVA-1 influences alternative splicing and is linked to lifespan extension and stress recovery in mutants.
The circRNA circ-crh-1 is a shared target of NOVA-1's regulation of splicing and circRNA production.
Abstract
Circular RNA (circRNA) biogenesis is regulated by RNA-binding proteins (RBPs) that alter back-splicing of exons in protein-coding genes. However, few in vivo roles for RBPs in the regulation of circRNA biogenesis have been characterized. We previously showed that many circRNAs increase with age in Caenorhabditis elegans, and that loss of circ-crh-1, an abundant age-accumulated circRNA, extends mean lifespan. Given the established role of the mammalian RBP NOVA2 in promoting circRNA biogenesis, we investigated whether nova-1, the sole C. elegans homolog of NOVA1/2, similarly regulates circRNA expression and function in vivo. RNA-sequencing of nova-1 mutants compared to wild-type identified 686 circRNAs. Of these, 103 were differentially expressed in nova-1 mutants compared to wild-type, with 76 upregulated and 27 downregulated circRNAs, suggesting NOVA-1 acts as a negative regulator of a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
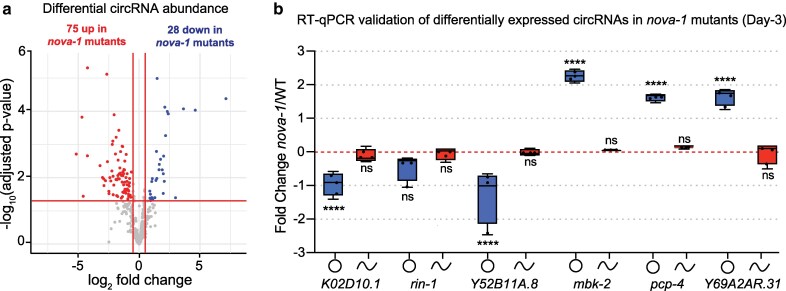 Fig. 1
Fig. 1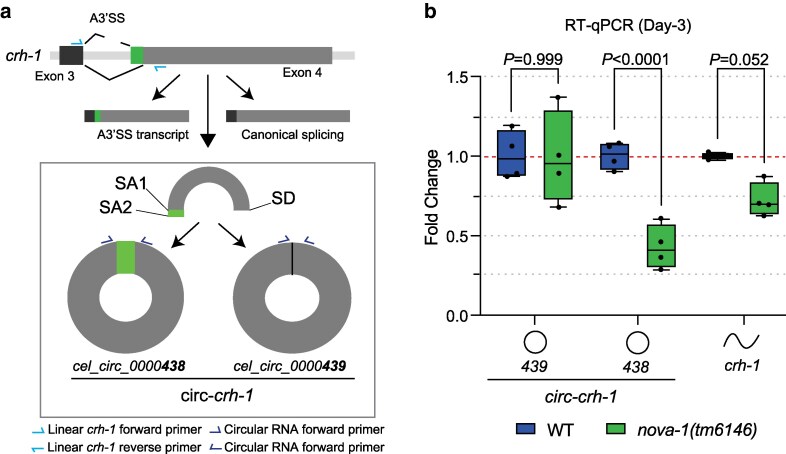 Fig. 2
Fig. 2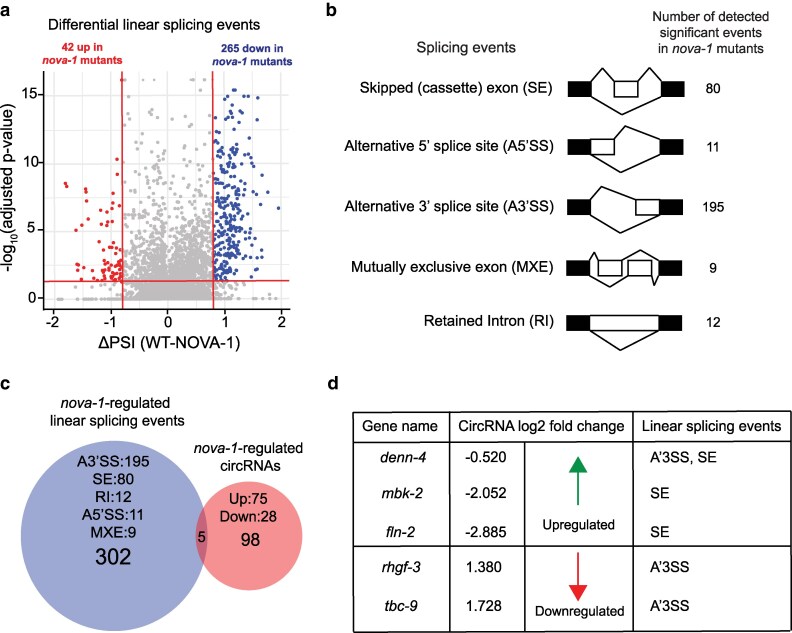 Fig. 3
Fig. 3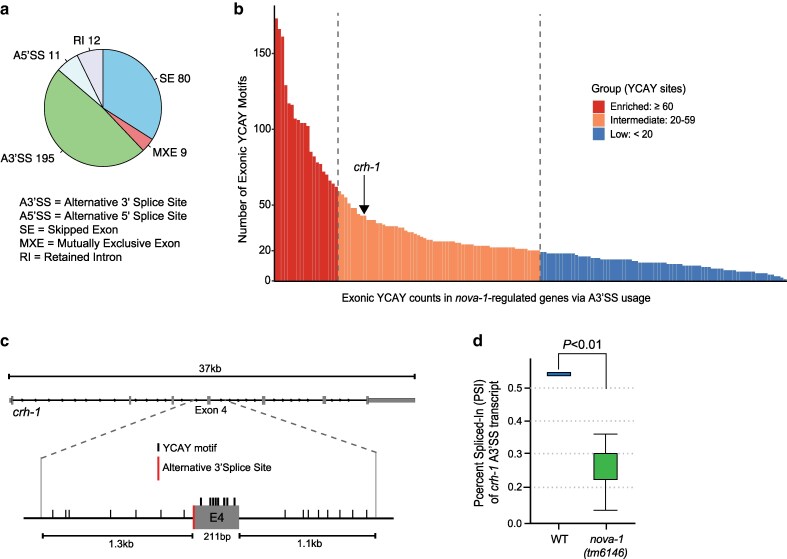 Fig. 4
Fig. 4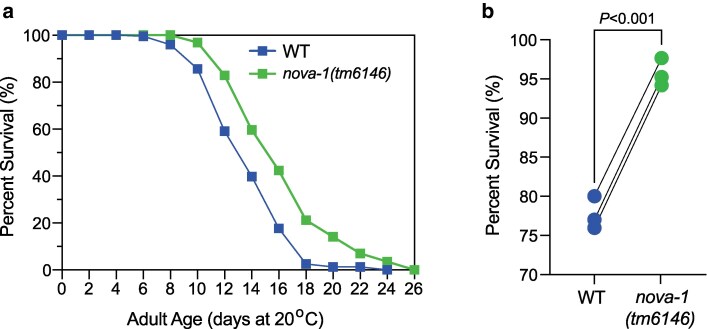 Fig. 5
Fig. 5- —National Institutes of Health10.13039/100000002
- —Cellular and Molecular Imaging (CMI)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA Research and Splicing · Circular RNAs in diseases · Genetics, Aging, and Longevity in Model Organisms
Introduction
Circular RNAs (circRNAs) are generated through back-splicing, a non-canonical splicing event in which a downstream 5′ splice donor (SD) joins an upstream 3′ splice acceptor (SA), forming a covalently closed loop marked by a unique back-splice junction (BSJ) (Cortes-Lopez and Miura 2016). Back-splicing is regulated by a complex interplay of cis-elements and trans-acting factors that promote looping of intronic regions flanking the circularizing exon(s) (Chen 2020), thereby facilitating circRNA formation. Inverted repeat sequences within introns can base-pair to bring SD and SA sites into proximity (Ashwal-Fluss et al. 2014; Ivanov et al. 2015), while RNA-binding proteins (RBPs) enhance back-splicing by stabilizing looped RNA structures and binding intronic motifs (Conn et al. 2015; Errichelli et al. 2017).
The NOVA family of splicing factors, NOVA1 and NOVA2, has emerged as key regulators of splicing in neuronal tissues (Piton 2025). NOVA2, a neuron-specific splicing factor that binds YCAY motifs (where Y indicates pyrimidines T or C), is essential for promoting circRNA formation during brain development (Knupp et al. 2021). Embryonic Nova2 knockout mice exhibit a global reduction in neuronal circRNAs, both in whole embryonic cortex and in isolated excitatory and inhibitory neurons (Saito et al. 2016; Knupp et al. 2021), underscoring NOVA2's role in facilitating circRNA formation in cortical neurons. In contrast, NOVA1, a paralog of NOVA2 with functions in alternative splicing and neurons (Buckanovich et al. 1993; Jensen et al. 2000), does not cause a widespread effect on circRNA levels when deleted (Saito et al. 2016; Knupp et al. 2021), indicating that circRNA regulation is a unique function of NOVA2. Notably, NOVA2-regulated circRNAs show minimal overlap with its exon-skipping targets (Knupp et al. 2021), suggesting a mechanistically distinct role for NOVA2 in circRNA biogenesis apart from alternative splicing activity. However, in vivo functional analysis of NOVA proteins remains limited due to the early lethality of NOVA-null mice.
The nematode Caenorhabditis elegans expresses a single NOVA homolog, nova-1, providing a genetically tractable model to investigate the in vivo role of NOVA proteins in circRNA regulation and alternative splicing. While mammalian NOVA2 promotes circRNA formation independently of its splicing function (Knupp et al. 2021), it remains unexplored whether C. elegans nova-1 exerts similar dual functions. Here, we show that nova-1 regulates circRNA expression and alternative splicing, largely through distinct mechanisms. RNA-seq of nova-1 mutants revealed 103 differentially expressed circRNAs, including the lifespan-associated circ-crh-1. Splicing analysis revealed significant changes in alternative 3′ splice site usage and exon skipping, with little overlap between circRNA regulation and splicing targets mediated by NOVA-1. However, the expression of circ-crh-1 represents a unique shared regulatory target in which NOVA-1 influences crh-1 circRNA formation through 3′-alternative splicing. Lastly, nova-1 mutants have an extended mean lifespan and improved heat stress recovery, highlighting NOVA-1 as a key regulator of aging and stress resilience in C. elegans. Together, these findings suggest that NOVA-1 is a key regulator of circRNA expression and alternative splicing in C. elegans, with likely downstream consequences for organismal lifespan and stress resilience.
Material and methods
elegans maintenance
C.
Worms were cultivated on the surface of NGM agar seeded with the Escherichia coli strain OP50 as the primary food source and grown in 20 °C incubators using standard protocols unless indicated otherwise. We used the wild-type strain N2, variety Bristol (Brenner 1974), and VDL1105 nova-1(tm6146) mutant strain, which was outcrossed at least six times. The nova-1 genotype was confirmed by PCR by identifying the 582 bp deletion using specific primers (Supplementary Table 1).
Total RNA collection and extraction
Worms were age-synchronized by hypochlorite treatment and collected eggs were hatched overnight at 20 °C in 1× M9 buffer. L1 larvae were then plated onto NGM plates seeded with 10× concentrated E. coli OP50 bacteria and allowed to develop to the L4 larval stage at 20 °C. L4 larvae were then collected, washed, and re-plated onto 25 µM 5-fluorodeoxyuridine (FUdR) (Milipore Sigma, Cat #50-91-9) containing NGM plates, unless indicated otherwise, seeded with 10× E. coli OP50 bacteria in order to prevent progeny formation. Three-day-old adult worms were collected by filtering through a 35 µM nylon mesh to remove bacteria (Sefar, Cat #7050-1220-000-10). Worm pellets were then transferred into green RINO tubes (Next Advance) and TRizol LS reagent (Thermo Fisher Scientific, Cat #10296028) was added in a 1:3 ratio. Worms were immediately lysed by bead beating them for 5 min using a Bullet Blender Pro Storm (Next Advance). Total RNA was extracted using the Purelink RNA mini-kit, followed by a DNAse I treatment following the manufacturer's protocol (Ambion, Cat #12183020). RNA was quantified by a Nanodrop. Bioanalyzer or tapestation (Agilent) was used for qualification as needed, and samples were stored at −80 °C.
RNA-Seq and circRNA prediction, mapping, and quantification
One microgram of total RNA was extracted from 5 independent biological replicates of wild-type (N2) and nova-1(tm6146) (VDL1105) animals, prepared and sequenced on an Illumina HiSeq 6000 system to obtain paired-end 150nt reads at the University of Connecticut genomics center. Raw FASTQ files were aligned to the WBcel235/ce11 reference genome using HISAT2 v2.2.1 (parameters: -no-mixed -no-discordant). Unmapped reads were realigned to the reference genome using BWA v0.7.18-r1243 mem followed by circRNA loci prediction using CIRI2 v2.0.6 (Gao et al. 2018). Predicted circRNAs with junction reads <12 were filtered out. From the predicted circRNA loci, a 200-nt circRNA junction scaffold was built and Bowtie2 v2.5.4 (Langmead and Salzberg 2012) was used for alignment and mapping to raw FASTQ datasets. Duplicate reads were removed after mapping with Picard (http://broadinstitute.github.io/picard) (parameters: MarkDuplicates ASSUME_SORTED = true REMOVE_DUPLICATES = true). Circular RNA counts in each sample were quantified using featureCounts v2.0.1 (Liao et al. 2014). edgeR v4.0.16 was used for differential expression analysis and to calculate Fold-change count values between nova-1 and wild-type samples (Chen et al. 2025). Differential expression testing was performed with quasi-likelihood F-tests (glmQLFit and glmQLFTest), which fits a generalized linear model under a negative binomial distribution. The resulting P-values were corrected for multiple testing using the Benjamini–Hochberg false discovery rate (FDR) method. CircRNAs were considered significantly upregulated in nova-1 mutants if log_2_ fold change (log_2_FC) < −0.5 with P < 0.05, and downregulated if log_2_FC > 0.5 with P < 0.05. This threshold corresponds to at least a ∼1.4-fold change in expression.
Multivariate analysis of transcript splicing (rMATS) and shiba
For alternative splicing analysis, we applied rMATS-turbo v4.3.0 to calculate significant skipped exon (SE), alternative 3′ and 5′ splice site usage (A3′SS & A5′SS), mutually exclusive exons (MXE), and retained intron (RI) splicing events in nova-1 mutants vs wild-type controls. rMATS utilizes a linear mixed model to compute the “Percent Spliced In” (PSI) value by evaluating the splicing levels of alternative/mutually exclusive exons, alternative 3′ or 5′ splice sites, or intron retention in the samples. rMATS uses a hierarchical framework based on a likelihood-ratio test, and the resulting P-values were adjusted for multiple testing using the Benjamini–Hochberg false discovery rate (FDR) method (Shen et al. 2014; Wang et al. 2024). Briefly, FASTQ files were mapped using STAR 2.7.10a using default parameters and subsequently aligned to the WBcel235/ce11 genome. rMATS-v.4.3.0 was used to discover significant linear alternative splicing events with the default parameters. For post-processing, junction count and exon count (*JCEC.txt) output files of all 5 splicing events were obtained and uploaded into R. To further process the output, the R package maser (Mapping Alternative Splicing Events to pRoteins) was used (https://www.bioconductor.org/packages/devel/bioc/vignettes/maser/inst/doc/Introduction.html). Significant splicing events were selected by applying filtering threshold of FDR ≤ 0.05 and |ΔPSI| ≥ 0.2 to the “topEvents” maser function, thereby selecting for splicing changes that were at least 20% different between nova-1 mutants and wild-type. The “plotTranscripts” function was used to visualize splicing events in R. Custom scripts were used to count the number of nova-1 YCAY ([C|T]CA[C|T]) binding sites in exons of alternatively spliced transcripts. Sashimi plot for splicing output visualization was generated following the scripts provided at https://github.com/Sika-Zheng-Lab/shiba2sashimi?tab=readme-ov-file after running Shiba (Kubota et al. 2025).
RT-qPCR analysis
To quantify and confirm individual circular or linear transcripts, 0.5 μg total RNA was reverse transcribed using Superscript III to prepare cDNA using random hexamers (Invitrogen, Cat #18080051). Next, cDNA samples were diluted and used with PowerUp SYBR Green Master Mix (Applied Biosystems, Cat #A25471) for RT-qPCR analysis, analyzed on a CFX96 Real-Time System (Bio-Rad). For RT-qPCRs of circRNAs, we used outward-facing primers. For host gene linear RNA counterparts, one primer was located in the circularizing exon, and the other was located in the upstream or downstream non-circularizing exon. For linear mRNAs, we used forward-facing primers. Fold-change values were calculated using wild-type (N2) ΔCt as control values for the 2^−ΔΔCt^ method. Data is normalized to housekeeping genes (cdc-42) mRNA. Primer sequences are listed in Supplementary Table 1.
Lifespan assay
Strains were maintained at 20 °C for at least 2 generations before the lifespan assay. Adult worms age-synchronized by hypochlorite treatment were allowed to lay eggs on NGM plates seeded with 10× concentrated E. coli OP50 bacteria over ∼3 h, and then removed. The resulting progeny synchronized by the timed egg-laying were allowed to develop into L4 larvae at 20 °C. At the L4 stage, 150 worms per genotype were transferred to new 6 cm NGM plates with freshly seeded 10× concentrated E. coli OP50 bacteria. Wild-type controls (N2) were assayed in parallel to mutants in the absence of FUdR and at 20 °C. Adult worms were transferred every 2 d during active reproduction, and each plate contained 10–15 worms. Worms that experienced ventral rupture, bagging, or walling were censored from the life-span analysis. A worm was considered dead when it did not respond to touch of the platinum wire pick, and was subsequently removed from the plate.
Heat shock viability assay
Animals age-synchronized by hypochlorite treatment were allowed to grow at 20 °C to reach L4 developmental stage. L4 animals were exposed to 35 °C heat-shock in a water bath for 4 h. Following the heat shock treatment, worms were recovered at 20 °C for 24 h on NGM plates seeded with E. coli OP50 before scoring viability.
Rigor and experimental statistics
To minimize experimenter bias in lifespan and stress assays, the experimenter was blinded to the genotypes and experimental conditions of the tested animals. Statistical analysis was performed using GraphPad Prism (version 10.6.1) software. Unpaired t-tests, two-way ANOVA, and Mantel-Cox tests, as well as likelihood-ratio tests within software packages (i.e. rMATS) were performed as indicated in the figure legends. A value of P ≤ 0.05 was used to determine statistical significance.
Results
NOVA-1 regulates a subset of circRNAs
Using RNA-sequencing, we analyzed circRNA expression in whole nova-1 mutants compared to whole wild-type worms in 3-d-old adults. CircRNA loci were identified from the RNA-seq dataset using CIRI2, a tool that detects back-splice junctions (BSJ) from RNA-seq data (Gao et al. 2018). In total, we detected 686 circRNAs (Supplementary Table 1). Of these, 103 circRNAs were differentially expressed in nova-1 mutants compared to wild-type, with 76 upregulated and 27 downregulated circRNAs (Fig. 1a; Supplementary Table 1) (|Fold-change| > 1.4, adj. P < 0.05). RT-qPCR validation of select circRNAs using outward-facing primers confirmed the directionality of expression changes in selected circRNAs, including 2 upregulated and 3 downregulated circRNAs (Fig. 1b). The rin-1 gene produces multiple circRNAs, and the selected circRNA did not exhibit a significant change in expression in nova-1 mutants (P = 0.0874) (Fig. 1b). Importantly, the corresponding linear transcripts of the host genes did not show significant expression changes (Fig. 1b). Notably, 73 of the 103 differentially expressed circRNAs overlapped with a previously characterized set of age-associated circRNAs (Supplementary Fig. 1d) (Cortés-López et al. 2018). 77% (56/73) of these circRNAs are upregulated in nova-1 mutants, and concurrently downregulated in wild-type, while 23% (17/73) are downregulated. For example, this included cel_circ_new_0000088, derived from the Y52B11A.8 gene, which was significantly downregulated in nova-1 mutants (P = 8.42e-05). Together, these findings suggest that NOVA-1 tends to inhibit the expression of a subset of age-regulated circRNAs in C. elegans.
*NOVA-1 regulates a subset of circRNAs. a) Volcano plot of differentially expressed circRNAs in whole animals of nova-1 mutants compared to wild-type control as determined by RNA-seq. Significantly downregulated and upregulated genes (Log2 fold-change >2, adj. P < 0.05), blue color represents down-regulated circRNAs, red color represents upregulated circRNAs. Significance was determined by a quasi-likelihood F-test under a negative binomial model. P-values were adjusted using the Benjamini–Hochberg false discovery rate (FDR) method. b) RT-qPCR analysis of circular and linear differentially regulated transcripts in day-3 adults of nova-1 mutants compared to wild-type. Shown are 3 upregulated and 3 downregulated circRNAs. n = 3 independent biological replicates. Data was normalized to cdc-24 mRNA, and is represented as the mean ± SEM; ns, not significant; ***P < 0.001. Statistical comparisons were performed with a two-way ANOVA followed by a Sidak's post hoc multiple comparisons test.
Distinct circ-crh-1 isoform regulation by NOVA-1
We previously identified 2 circRNAs derived from the crh-1 gene (i.e. cel_circ_0000438 and cel_circ_0000439) (Knupp et al. 2022). These circRNAs are 2 of the most abundant, age-accumulated circRNA isoforms derived from the crh-1 host gene. The circRNAs differ by only in 6 nucleotides, as a result of an alternative splice acceptor site (A3′SS) in exon 4 of the crh-1 locus, and are collectively referred to as circ-crh-1 (Fig. 2a). To determine whether NOVA-1 influences circ-crh-1 formation, we quantified the expression of the 2 circ-crh-1 isoforms using RT-qPCR in 3-d-old adults. We observed a significant reduction in cel_circ_0000438 in nova-1 mutants, while levels of cel_circ_0000439 and linear crh-1 were not significantly changed, although linear crh-1 showed a marginal trend toward reduction (P = 0.052) (Fig. 2b). These findings suggest that loss of NOVA-1 modulates the expression of one circ-crh-1 isoform (ie cel_circ_0000438) likely by selectively impairing back-splicing at one acceptor site without affecting the other circ-crh-1 isoform or linear crh-1. Alternatively, NOVA-1 may simply regulate the selection of the A3′SS at the crh-1 exon 4 locus, which in turn affects both circular and linear splicing outcomes.
NOVA-1 differentially regulates 2 circ-crh-1 isoforms. a) Schematic showing the alternative site usage in exon 4 of crh-1. Two circRNAs (cel_circ_0000438 and cel_circ_0000439) are generated by back-splicing of exon 4, using 2 alternative splice acceptors (SA) and one shared splice donor (SD). b) RT-qPCR analysis of circular and linear circ-crh-1 transcripts in 3-d-old adults at 20 °C of nova-1 mutants compared to wild-type. nova-1 mutants reduce cel_circ_0000438 expression but not cel_circ_0000439, whereas the linear crh-1 transcript is not significantly changed. Linear and circRNA oligo primers used for RT-qPCR analysis are shown in panel b. n = 4 independent biological samples. Data are normalized to cdc-24 mRNA and are represented as the mean ± SEM. Statistical comparisons were performed with a two-way ANOVA followed by a Sidak's post hoc multiple comparisons test.
NOVA-1 regulates alternative splicing events
We performed analysis of linear alternative splicing using rMATS-turbo v4.3.0 (Shen et al. 2014; Wang et al. 2024) on all wild-type and nova-1 mutant samples, identifying over 16,000 linear splicing events. After applying stringent filtering (FDR ≤ 0.05, |ΔPSI| ≥ 0.2), 307 total events were classified as differentially expressed in nova-1 mutants compared to wild-type controls (Figs. 3a and 4a; Supplementary Table 4). Of these, 195 (64%) involved alternative 3′ splice site (A3′SS) usage, and 80 (26%) were skipped exons (SE) (Figs. 3b and 4a). The remaining 10% comprised alternative 5′ splice sites (5A′SS), retained introns (RI), and mutually exclusive exons (MXE) (Figs. 3b and 4a), indicating widespread splicing disruption in the absence of NOVA-1.
NOVA-1 regulates distinct gene sets during pre-mRNA alternative splicing and back-splicing programs. a) Volcano plot showing total splicing changes in nova-1 mutants compared to wild-type control. Points represent transcripts from any of the 5 linear splicing events (i.e. A3′SS, A5′SS, SE, MXE, and RI). Upregulated and downregulated splicing events correspond to points with percent splice in changes (ΔPSI = WT_PSI—NOVA-1_PSI) ≤ −0.2 (red) and ΔPSI ≥ 0.2 (blue), respectively (FDR ≤ 0.05; adj. P < 0.05). b) Schematic representation of the 5 splicing events and the number of detected significant events in nova-1 mutants vs wild-type (FDR ≤ 0.05, |ΔPSI| ≥ 0.2). Alternative 3′ splice site usage (A3′SS) and Skipped exons (SE) are the most abundant of the AS events, both accounting for approximately 90% of nova-1-regulated linear splicing events. Alternative 5′ splice site usage (A5′SS), retained introns (RI) and exclusive exons (MXE) account make up the remaining 10% of splicing events regulated by nova-1. White exons (boxes) indicate alternatively “spliced in” exons during linear pre-mRNA alternative splicing. c-d) Venn diagram showing the relationship between nova-1-regulated linear splicing and back-splicing events. CircRNAs derived from 3 of the 5 host genes in this category are more abundant in nova-1 mutants compared to wild-type controls and all have lower skipped exon PSI (see d), with the exception of denn-4, which, in addition, has a lower A3′SS usage compared to control, alongside 2 nova-1-regulated low abundance circRNAs (see d).
NOVA-1 modulates alternative splice site selection at the crh-1 locus. a) Pie chart of 5 splicing events with the number of detected significant events in nova-1 mutants vs wild-type (FDR ≤ 0.05, |ΔPSI| ≥ 0.2) for alternative 3′ splice site usage (A3′SS), alternative 5′ splice site usage (A5′SS), skipped exons (SE), retained introns (RI), and exclusive exons (MXE). b) Number of exonic YCAY motif sites within alternatively spliced transcripts regulated by NOVA-1 categorized by the number of YCAY sites: low (<20), intermediate (20 to 59), and enriched (>60). c) Schematic of the crh-1 gene with a total of 43 YCAY motif sites with 8 shown in exon 4, which is subject to A3′SS usage. In addition, 7 and 8 YCAY sites are shown in upstream and downstream intronic sequences of exon 4, respectively. d) Reduced percent spliced In (PSI) at exon 4 of crh-1 (A3′SS) in nova-1 mutants. Differential splicing was quantified using rMATS, which applies a likelihood-ratio test on exon inclusion levels between samples with Benjamini–Hochberg false discovery rate (FDR) correction. PSI values were visualized using the maser R package.
Next, we examined YCAY motif sites in alternatively spliced transcripts regulated by NOVA-1. Exonic YCAY sites were categorized into 3 different groups: low (<20 sites), intermediate (20–59 sites), and enriched (>60 sites). This analysis revealed that 168 (55%) of the 307 significant splicing events contained 20 or more YCAY motif sites (Fig. 4b; Supplementary Table 4). Focusing on the crh-1 gene, we identified a total of 43 YCAY sites (Fig. 4c; Supplementary Table 4) with 8 located in exon 4 (Fig. 4c), which is subject to A3′SS usage. Notably, this represents at least 3 more YCAY sites than were identified in any other crh-1 exon, suggesting preferential NOVA-1 binding near exon 4. In addition, 7 and 8 YCAY sites were identified in the upstream and downstream intronic regions flanking exon 4, respectively (Fig. 4c). A3′SS selection at exon 4 of crh-1 generates 2 isoforms of the same exon differing by 6 nucleotides (205 nucleotides compared to 211 nucleotides, Fig. 2a). Our data suggest that loss of nova-1 favors canonical 205-nt exon 4 splicing and reduces selection of the A3′SS isoform (Fig. 4d; Supplementary Fig. 1b). Since the YCAY motif is short and its presence in A3′SS transcripts could occur by chance, we also examined YCAY occurrences in more than 2,000 transcripts that showed no difference in A3′SS usage (negative controls) between wild-type and nova-1 mutant animals. Among these non-significant transcripts, only 14% contained over 20 exonic YCAY motifs (intermediate-enriched), compared to more than 48% of the 195 transcripts in the significant dataset (Supplementary Fig. 1c; Supplementary Table 4). This enrichment pattern indicates that YCAY tetranucleotide occurrences are not random and likely play an important role in NOVA-1-mediated regulation of A3′SS usage. Together, these findings suggest that NOVA-1-mediated alternative splicing may intersect with its role in back-splicing.
Minimal overlap between NOVA-1-regulated circRNAs and alternative splicing targets
Since nova-1 mutations affect both back-splicing (103 circRNAs) and alternative splicing events (307 linear transcripts), we next examined potential overlap between these 2 regulatory processes. Of the 103 differentially expressed circRNAs, only 5 showed overlap with nova-1-regulated linear splicing events (Fig. 3c). Specifically, 3 of the 75 upregulated and 2 of the 28 downregulated circRNAs overlapped with nova-1-regulated linear splicing events (Fig. 3d). All overlapping splicing events fell within alternative 3′ splice site (A3′SS) or skipped exon (SE) categories, with denn-4 undergoing both A3′SS usage and SE (Fig. 3d). Interestingly, the crh-1 gene displayed a reduced Percent Spliced In (PSI) at exon 4 (A3′SS) in nova-1 mutants (Fig. 4d; Supplementary Fig. 1b). This together with our RT-qPCR analysis (Fig. 2b) supports a model in which NOVA-1 differentially regulates the expression of circ-crh-1 isoforms by modulating alternative splicing. Overall, although NOVA-1 impacts both alternative splicing and circRNA expression, their regulatory overlap is confined to a small subset of targets.
Loss of NOVA-1 extends mean lifespan and enhanced stress resistance
Given that we previously showed that loss of circ-crh-1 circRNAs extends mean lifespan (Knupp et al. 2022), we next tested whether nova-1 mutants exhibit similar lifespan phenotypes. Lifespan assays revealed that nova-1 mutant animals have a significant 14.75% increase in mean lifespan compared to wild-type controls (P < 0.0001, Fig. 5a). Because enhanced longevity is often associated with improved stress resistance (Zhou et al. 2011), we also examined thermotolerance as a potential contributing factor. Following a 4-h heat shock at 35 °C, nova-1 mutant adults showed markedly improved recovery relative to wild-type animals (Fig. 5b), suggesting that increased stress resilience may underlie the extended lifespan observed in these nova-1 mutants. Together, these findings suggest that the altered expression of circRNAs, together with changes in 3′ alternative splicing and exon skipping events, may contribute to the increased mean lifespan and stress resistance phenotype of nova-1 mutants.
nova-1 mutants extend mean lifespan and increase heat resistance. a) Lifespan curves for nova-1 mutant animals compared to wild-type. nova-1 mutants extend mean lifespan compared to wild-type (P < 0.0001, Mantel–Cox log-rank test). n = 2 independent lifespan assays were performed with n = 150 animals at 20 °C for each assay and genotype in the absence of FUdR. b) Loss of nova-1 during heat shock results in increased viability. Heat shock treatment was performed on synchronized L4 worms for 4 h at 35 °C and the percentage of survival was determined after 24 h recovery at 20 °C for nova-1 mutants compared to wild-type controls. nova-1 mutants exhibit increased resistance to heat exposure compared to wild-type. Three biological replicates with n = 90 animals for each survival assay and genotype in the absence of FUdR. Statistical comparisons between wild-type and nova-1 mutants were performed with an unpaired two-tailed t-test.
Discussion
Our study shows that the RNA-binding protein NOVA-1 regulates the expression of a subset of circRNAs in C. elegans, including the age-associated cel_circ_0000438 (circ-crh-1). Using RNA-seq and RT-qPCR analysis, we identified circRNAs whose expression are altered in nova-1 mutants. Interestingly, the majority of differentially expressed circRNAs were upregulated in nova-1 mutants, suggesting that unlike NOVA2 in mammals where it promotes circRNA production by binding intronic YCAY motifs to enhance back-splicing in neural tissues (Knupp et al. 2021), NOVA-1 in C. elegans may act as a negative regulator of circRNA expression. However, because our RNA-seq analysis was performed on whole worms, it remains possible that NOVA-1 in C. elegans promotes circRNA formation in select tissues such as neurons.
Beyond circRNA regulation, we observed widespread splicing disruptions in nova-1 with the majority of significant changes involving alternative 3′ splice site (A3′SS) selection and exon skipping (SE). This enrichment of A3′SS is consistent with findings from mammalian systems where it was observed that NOVA1/2 proteins bind YCAY sites to regulate diverse alternative splicing outcomes including cassette exons, intron retention and importantly A3′SS usage (Ule et al. 2005; Licatalosi et al. 2008; Zhang et al. 2010). More recently, over 2,000 NOVA1/2-regulated splicing events were identified in the developing mouse cortex, many of which involved A3′SS usage (Saito et al. 2016). Thus, the enrichment of A3′SS events in C. elegans nova-1 mutants is highly similar with NOVA's established splicing functions in vertebrates. Moreover, similar to mammalian NOVA2, where circRNA abundance is largely uncoupled from linear alternative splicing (Knupp et al. 2021), we observed that only a small subset of differentially expressed circRNAs in nova-1 mutants overlapped with genes exhibiting altered linear splicing. The crh-1 locus illustrates this partial convergence where reduced expression of cel_circ_0000438 in nova-1 mutants corresponds with a shift in splice acceptor site usage, whereas expression of the closely related cel_circ_0000439 and linear crh-1 remains unaffected. This may be explained by A3′SS selection at exon 4 of the crh-1 locus, which generates 2 isoforms of the exon that differ by only 6 nucleotides. The 2 crh-1-derived circRNAs (cel_circ_0000438 and cel_circ_0000439) originate from these same alternative exon 4 isoforms. Consequently, decreased A3′SS selection in nova-1 mutants corresponds to reduced production of the circRNA generated from the longer exon 4 (cel_circ_0000438), while the circRNA derived from the shorter exon 4 isoform (cel_circ_0000439) remains unaffected. Therefore, NOVA-1 loss negatively affects both A3′SS selection in the linear transcript and production of the corresponding circRNAs. This parallel regulation suggests that NOVA-1 influences 3′ splice site choice, which in turn impacts both linear and circular splicing. Several studies and reports (Knupp and Miura 2018; Li et al. 2018; Wang et al. 2025) have described competition between canonical splicing and back-splicing, and our findings are consistent with NOVA-1 acting to balance these splicing pathways at crh-1 exon 4. Upon NOVA-1 loss, the spliceosome preferentially selects the canonical 3′ splice site, leading to reduced A3′SS selection and back-splicing. These findings suggest that NOVA-1 in C. elegans functions through both shared mechanisms by modulating splice acceptor site choice that influences circRNA back-splicing and distinct pathways that regulate circRNA levels independent of linear splicing.
The position of YCAY motif sites in pre-mRNA determines how NOVA influences splicing regulation (Ule et al. 2006; Witten and Ule 2011). Additionally, NOVA selectively inhibits or promotes alternative splicing regulation depending on whether the sites are exonic or intronic, respectively (Ule et al. 2006). We identified YCAY sites in exon sequences and found enrichment of these sites in over half (55%) of the NOVA-1-regulated transcripts. Interestingly, this exonic enrichment did not result in widespread inhibition of NOVA-1-regulated alternative splicing events in wild-type C. elegans, as observed in mammals (Ule et al. 2006). A reasonable explanation may be that C. elegans introns are generally shorter and lack strong polypyrimidine tracts compared to mammals, which could alter the way NOVA-1 engages with the spliceosome and reduce the likelihood of reproducing the same splicing regulatory outcomes as observed in mammals. Despite this apparent difference, our analysis revealed that SE still represented the most affected alternative splicing category, with 24 of 80 events (30%) reduced in wild-type compared to nova-1 mutants. Overall, while the NOVA-mediated bidirectional control of alternative splicing established in mammals provides useful general insights (Ule et al. 2006), our findings suggest that NOVA-regulated alternative splicing in C. elegans follows the same underlying principles but plays out differently due to species-specific intron differences and splicing constraints.
Taken together, these findings extend our view of NOVA proteins as key regulators of alternative splicing and circRNA regulation. This study expands our understanding of RBP-mediated circRNA regulation and highlights NOVA-1's dual role in circRNA expression and splice site choice that may contribute to the regulation of lifespan and stress resistance in C. elegans.
Supplementary Material
jkag016_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ashwal-Fluss R et al 2014. circ RNA biogenesis competes with pre-m RNA splicing. Mol Cell. 56:55–66. 10.1016/j.molcel.2014.08.019.25242144 · doi ↗ · pubmed ↗
- 2Brenner S . 1974. The genetics of Caenorhabditis elegans. Genetics. 77:71–94. 10.1093/genetics/77.1.71.4366476 PMC 1213120 · doi ↗ · pubmed ↗
- 3Buckanovich RJ, Posner JB, Darnell RB. 1993. Nova, the paraneoplastic Ri antigen, is homologous to an RNA-binding protein and is specifically expressed in the developing motor system. Neuron. 11:657–672. 10.1016/0896-6273(93)90077-5.8398153 · doi ↗ · pubmed ↗
- 4Chen LL . 2020. The expanding regulatory mechanisms and cellular functions of circular RN As. Nat Rev Mol Cell Biol. 21:475–490. 10.1038/s 41580-020-0243-y.32366901 · doi ↗ · pubmed ↗
- 5Chen Y, Chen L, Lun ATL, Baldoni PL, Smyth GK. 2025. Edger v 4: powerful differential analysis of sequencing data with expanded functionality and improved support for small counts and larger datasets. Nucleic Acids Res. 53:gkaf 018. 10.1093/nar/gkaf 018.39844453 PMC 11754124 · doi ↗ · pubmed ↗
- 6Conn SJ et al 2015. The RNA binding protein quaking regulates formation of circ RN As. Cell. 160:1125–1134. 10.1016/j.cell.2015.02.014.25768908 · doi ↗ · pubmed ↗
- 7Cortes-Lopez M, Miura P. 2016. Emerging functions of circular RN As. Yale J Biol Med. 89:527–537.28018143 PMC 5168830 · pubmed ↗
- 8Cortés-López M et al 2018. Global accumulation of circ RN As during aging in Caenorhabditis elegans. BMC Genomics. 19:8. 10.1186/s 12864-017-4386-y.29298683 PMC 5753478 · doi ↗ · pubmed ↗