Establishment of an in vivo-based assay using a silkworm infection model for phenotypic evaluation of antimicrobial drug combinations against Mycobacterium abscessus
Akiho Yagi, Motoko Shinohara, Yusuke Minato, Ryuji Uchida

TL;DR
A new silkworm model is developed to test antimicrobial drug combinations against Mycobacterium abscessus in a whole-organism setting.
Contribution
The introduction of a silkworm-based in vivo assay for evaluating antimicrobial drug interactions against M. abscessus.
Findings
The silkworm model produced interaction profiles consistent with in vitro results for tested drug combinations.
The assay is rapid, reproducible, and suitable for assessing synergistic or antagonistic drug effects.
Abstract
Mycobacterium abscessus exhibits high intrinsic drug resistance, requiring combination therapy. We developed a silkworm (Bombyx mori) infection model as a whole-organism, in vivo-based platform for quantitative, outcome-based evaluations of antimicrobial combinations. The system, examined using clarithromycin-amikacin and imipenem-cefoxitin combinations, showed interaction profiles that were qualitatively consistent with those observed in vitro. This rapid, reproducible, and ethical assay enables reliable phenotypic assessments of synergistic or antagonistic effects and may facilitate the evaluation and prioritization of antimicrobial combination regimens in preclinical studies.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
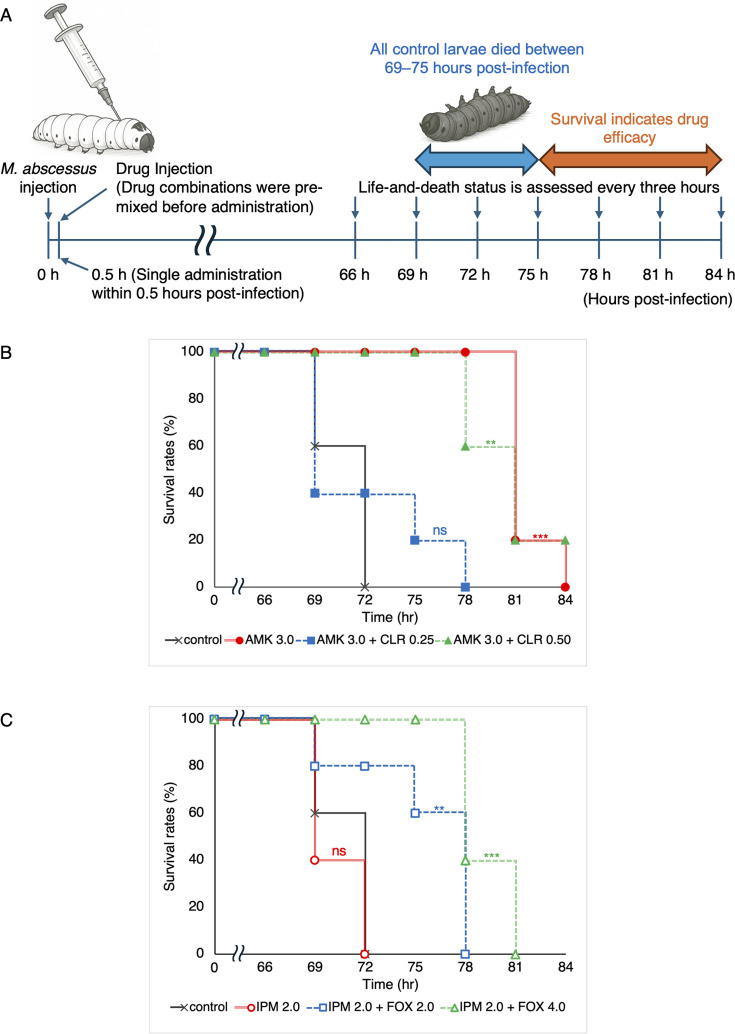 Fig 1
Fig 1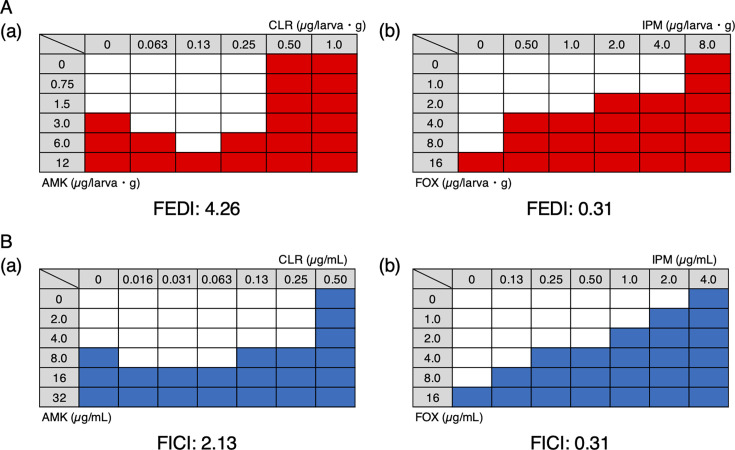 Fig 2
Fig 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycobacterium research and diagnosis · Tuberculosis Research and Epidemiology · Invertebrate Immune Response Mechanisms
INTRODUCTION
Infections caused by nontuberculous mycobacteria (NTM) are increasing worldwide and remain difficult to treat because of their intrinsic resistance to many commonly used antimicrobials (1). Among NTM, Mycobacterium abscessus is recognized as the most drug-resistant species, leaving few therapeutic options (1). Current clinical guidelines recommend prolonged multidrug combination therapy (2). Clarithromycin (CLR) resistance further limits available treatments, highlighting the need for additional agents that may be incorporated into combination therapy. Consequently, efforts to identify synergistic drug combinations are ongoing. Many combinations have been evaluated in vitro, and several, including dual β-lactams, have been examined in vivo (3, 4). Nevertheless, these in vivo studies are limited to specific dosing conditions, and comprehensive evaluations of dose-dependent drug interactions have rarely been conducted.
Although mammalian models, such as mice, are commonly used for these evaluations, the establishment of a persistent M. abscessus infection in these models is technically difficult and involves considerable cost and ethical concerns (5). Furthermore, it remains unclear whether in vitro drug interactions may be reliably reproduced in vivo. Therefore, the need for reliable alternative models that efficiently and quantitatively assess drug interactions in vivo is increasing. While in vitro combination assays are indispensable for analyzing direct antimicrobial interactions, they cannot capture host-dependent therapeutic outcomes. To address this limitation, we employed a silkworm infection model as a whole-organism system to quantitatively evaluate the effects of antimicrobial combinations on survival. This model is intended to complement in vitro assays by providing an outcome-based assessment of combination effects within an integrated biological context, rather than to recapitulate mammalian pharmacokinetics.
Alternative M. abscessus infection models using Drosophila melanogaster (6), zebrafish (7), and Galleria mellonella (8) have been reported, and each has its limitations. Drosophila and zebrafish cannot be maintained at 37°C, while the small size of G. mellonella hinders quantitative evaluations. In contrast, the silkworm infection model with M. abscessus can be maintained at 37°C and supports reproducible systemic infections, enabling quantitative assessments of antimicrobial efficacy within a short period (9). This model allows for the ethical testing of numerous individuals and aligns with the 3Rs principle. Furthermore, the silkworm’s small body weight enables evaluation with minimal compound quantities, making it advantageous for screening limited or valuable molecules. Based on these advantages, we selected the silkworm infection model and established a practical assay system for comprehensive, dose-dependent evaluations of antimicrobial combinations.
The efficacies of drug combinations were evaluated using a silkworm infection model with M. abscessus (Fig. 1A). Fifth-instar silkworm larvae (2.0 ± 0.1 g, n = 5) were infected with M. abscessus JCM13569 (ATCC19977) by injecting a bacterial suspension (approximately 2.5 × 10^6^ CFU/mL, 50 µL) into the hemolymph. We previously reported that this infection condition caused 100% mortality within 69–75 h at 37°C (9). To allow for a survival-based evaluation prior to natural death, silkworms were monitored for up to 84 h post-infection. Each drug was administered in a single 50 µL dose, either individually or as a premixed solution, within 30 min after infection. To select the appropriate dose range for the combination study, we initially confirmed that each drug exhibited dose-dependent therapeutic efficacy when administered alone (Table S1 and Fig. S1). Based on these results, five dose levels were selected for each drug, and all possible dose combinations were systematically evaluated. Kaplan-Meier survival curves were generated to assess life-prolonging effects (Fig. 1B and C), and detailed survival data and Kaplan-Meier curves for all combinations are provided in the Supplementary Information (Fig. S1 and Table S1).
*In vivo evaluation of drug efficacy using the silkworm infection model with M. abscessus. (A) Experimental scheme for the infection and treatment protocol. (B) Kaplan-Meier survival curves for AMK and CLR. Filled circles, AMK (3.0 µg/larva·g); filled squares, AMK (3.0 µg/larva·g) + CLR (0.25 µg/larva·g); filled triangles, AMK (3 µg/larva·g) + CLR (0.50 µg/larva·g); ×, control. (C) Kaplan–Meier survival curves for IPM and FOX. Open circles, IPM (2.0 µg/larva·g); open squares, IPM (2.0 µg/larva·g) + FOX (2.0 µg/larva·g); open triangles, IPM (2.0 µg/larva·g) + FOX (4.0 µg/larva·g); ×, control. Each experiment was performed twice, and reproducible results were obtained. Survival curves were compared with controls using the log-rank test (***P < 0.005, *P < 0.05, ns = not significant).
The results obtained confirmed that under the CLR and AMK combination, the co-administration of CLR at 0.25 µg/larva·g shortened survival significantly more than AMK alone (Fig. 1B). This antagonistic effect was also observed at lower CLR doses (0.13 and 0.063 µg/larva·g) (Fig. S1 and Table S1). However, no antagonism was observed when CLR was administered at 0.5 µg/larva·g, a dose that was as effective as monotherapy (Fig. 1B). Therefore, the antagonism between CLR and AMK reported in vitro (10) was also observed in the silkworm infection model. In contrast, for the IPM and FOX combination, the co-administration of IPM at 2 µg/larva·g with FOX at 2 or 4 µg/larva·g—doses that were individually ineffective—significantly prolonged survival (Fig. 1C). Similar results were observed across other dose levels, and the combination exerted clear synergistic effects in vivo (Fig. S1 and Table S1). This result is consistent with previously reported synergy in both in vitro studies and mouse models (3, 11), further supporting the validity of our evaluation system. This assay was also independently repeated on different dates and with different dosing lots, yielding reproducible results.
To further enhance the utility of this evaluation system, we established an index for quantitatively assessing drug combination effects in vivo. Survival rates at the time when all control larvae (injected without drug treatment) had died were visualized as heatmaps (Fig. 2A), with red cells indicating conditions achieving ≥80% survival, defined as 80% effective doses (ED_80_). Using this matrix, we calculated the fractional effective dose index (FEDI), defined as the sum of the ratios of each drug’s ED_80_ in combination with its ED_80_ as monotherapy.
Heat maps of the combinatorial efficacy of clinical drugs against M. abscessus. (A) (a) AMK and CLR, (b) IPM and FOX in vivo by a silkworm infection model. Red cells indicate wells showing ≥80% larval survival. (B) (a) AMK and CLR, (b) IPM and FOX in vitro by a microdilution method. Blue cells indicate wells showing ≥90% inhibition of bacterial growth.
This index allows intuitive interpretations, with values <0.5 indicating synergy, 0.5–1 indicating additivity, and >2 indicating antagonism—similar to the fractional inhibitory concentration index (FICI) used in vitro. As proof of concept, the CLR and AMK combination yielded FEDI of 4.26, demonstrating clear antagonism, whereas IPM and FOX resulted in FEDI of 0.31, indicating strong synergy. These values corresponded well with in vitro FICI values (2.13 and 0.31, respectively) (Fig. 2B), confirming the consistency of this in vivo system with established in vitro findings. Although optimal dosing ratios were not completely identical between the two systems, this may reflect pharmacokinetic and physiological factors not captured in vitro. Therefore, a quantitative in vivo evaluation is essential for bridging this gap and designing more clinically relevant combination regimens.
Previous studies have reported that pharmacokinetic parameters and ED₅₀ values obtained in the silkworm model show trends comparable to those observed in mammalian systems (12–14), supporting its practical utility for prioritizing drug combinations prior to mammalian testing. A limitation of the silkworm infection model is that drug exposure under the experimental conditions is relatively static and does not fully recapitulate mammalian pharmacokinetic profiles (15), as is often the case for alternative animal infection models that are not designed to reproduce mammalian pharmacokinetics. In this study, our objective was not to reproduce mammalian pharmacokinetics, but to evaluate drug-drug interactions as phenotypic therapeutic outcomes in a whole-organism infection context. Because single agents and their combinations were assessed under identical exposure conditions, relative changes in survival attributable to drug combinations could be quantitatively compared independent of PK matching.
Thus, while the silkworm model does not reproduce mammalian pharmacokinetics, it serves as a complementary platform for phenotypic evaluation of antimicrobial combinations, particularly in contexts where relative interaction effects rather than precise PK profiles are of interest.
In conclusion, we herein developed an in vivo-based assay system using a silkworm infection model for quantitative evaluations of antimicrobial drug combinations. This platform enables reproducible, dose-dependent assessments of synergistic and antagonistic interactions with a high throughput and minimal ethical burden. Thirty-six treatment conditions were evaluated twice for reproducibility, involving 360 silkworms (n = 5 per group × 36 groups × 2 experiments). This scale of whole-organism combination testing would be difficult to achieve using mammalian models, underscoring the practical utility of this system. In the future, this platform may be applied to evaluate and prioritize antimicrobial drug combinations prior to translation to mammalian models.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Johansen MD, Herrmann JL, Kremer L. 2020. Non-tuberculous mycobacteria and the rise of Mycobacterium abscessus. Nat Rev Microbiol 18:392–407. doi:10.1038/s 41579-020-0331-132086501 · doi ↗ · pubmed ↗
- 2Daley CL, Iaccarino JM, Lange C, Cambau E, Wallace RJ Jr, Andrejak C, Böttger EC, Brozek J, Griffith DE, Guglielmetti L, Huitt GA, Knight SL, Leitman P, Marras TK, Olivier KN, Santin M, Stout JE, Tortoli E, van Ingen J, Wagner D, Winthrop KL. 2020. Treatment of nontuberculous mycobacterial pulmonary disease: an official ATS/ERS/ESCMID/IDSA clinical practice guideline. Clin Infect Dis 71:e 1–e 36. doi:10.1093/cid/ciaa 24132628747 PMC 7768748 · doi ↗ · pubmed ↗
- 3Le Run E, Arthur M, Mainardi JL. 2018. Synergistic efficacy of β-lactam combinations against Mycobacterium abscessus pulmonary infection in mice. Antimicrob Agents Chemother 62:e 00623-18. doi:10.1128/AAC.00623-1831109979 PMC 6658775 · doi ↗ · pubmed ↗
- 4Lanni A, Iona E, Fattorini L, Giannoni F, Iacobino A. 2025. Activity of combinations of bactericidal and bacteriostatic compounds in Mycobacterium abscessus-infected mice: an overview. Front Microbiol 16:1616149. doi:10.3389/fmicb.2025.161614940822395 PMC 12354516 · doi ↗ · pubmed ↗
- 5Sui X, Oehlers SH. 2025. Animal models of Mycobacterium abscessus pulmonary infection phenotypes: What are we modeling? P Lo S Pathog 21:e 1013414. doi:10.1371/journal.ppat.101341440788902 PMC 12338768 · doi ↗ · pubmed ↗
- 6Oh CT, Moon C, Jeong MS, Kwon SH, Jang J. 2013. Drosophila melanogaster model for Mycobacterium abscessus infection. Microbes Infect 15:788–795. doi:10.1016/j.micinf.2013.06.01123831804 · doi ↗ · pubmed ↗
- 7Bernut A, Herrmann JL, Kissa K, Dubremetz JF, Gaillard JL, Lutfalla G, Kremer L. 2014. Mycobacterium abscessus cording prevents phagocytosis and promotes abscess formation. Proc Natl Acad Sci USA 111:E 943–E 952. doi:10.1073/pnas.132139011124567393 PMC 3956181 · doi ↗ · pubmed ↗
- 8Meir M, Grosfeld T, Barkan D. 2018. Establishment and validation of Galleria mellonella as a novel model organism to study Mycobacterium abscessus infection, pathogenesis, and treatment. Antimicrob Agents Chemother 62:e 02539-17. doi:10.1128/AAC.02539-1729437630 PMC 5914002 · doi ↗ · pubmed ↗