Pseudomonas aeruginosa tRNA nucleotidyltransferase Cca controls resistance and tolerance to aminoglycoside antibiotics by regulating the MexXY multidrug efflux pump
Chenyu Shen, Lepeng Wang, Yu Zhang, Linyan Zhang, Zhihui Cheng, Weihui Wu, Un-Hwan Ha, Shouguang Jin, Yongxin Jin

TL;DR
This study reveals how a tRNA enzyme in Pseudomonas aeruginosa influences antibiotic resistance by regulating a drug efflux pump.
Contribution
The study identifies a novel regulatory mechanism involving Cca enzyme and the MexXY efflux pump in antibiotic resistance.
Findings
Mutation of the cca gene increases resistance and tolerance to aminoglycosides in P. aeruginosa.
Cca controls the transcription of armZ through the leader peptide PA5471.1, which regulates mexXY.
Abstract
Pseudomonas aeruginosa displays high resistance to a wide range of antibiotics, greatly impeding clinical treatment. MexXY efflux pump plays an important role in bacterial resistance and tolerance to aminoglycoside antibiotics. tRNA nucleotidyltransferase (Cca, CCA-adding enzyme) is a universal polymerase responsible for the synthesis and repair of the 3′-terminal CCA sequence of tRNA. In this study, we found that mutation of the cca gene increases resistance and tolerance to aminoglycosides in P. aeruginosa. We demonstrate that the upregulation of mexXY genes contributes to the increased resistance and tolerance of the Δcca mutant. Furthermore, our experimental results revealed that Cca controls the transcription of armZ, which encodes a positive regulator of mexXY, through its leader peptide PA5471.1. An increased amount of PA5471.1 mRNA was found to be associated with ribosomes in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
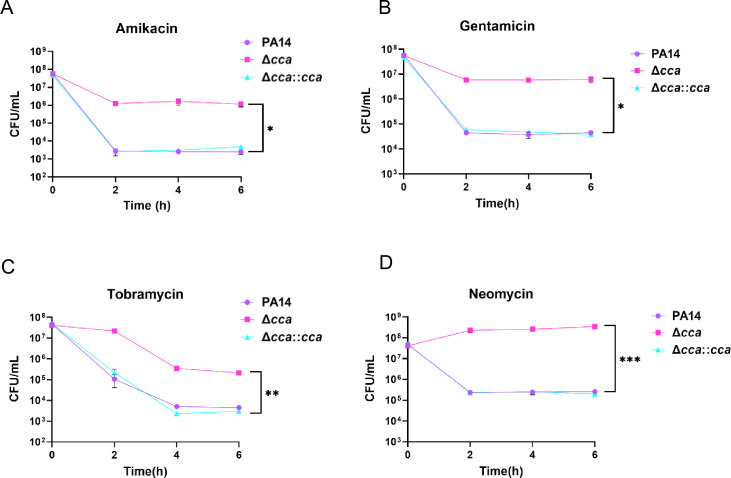 Fig 1
Fig 1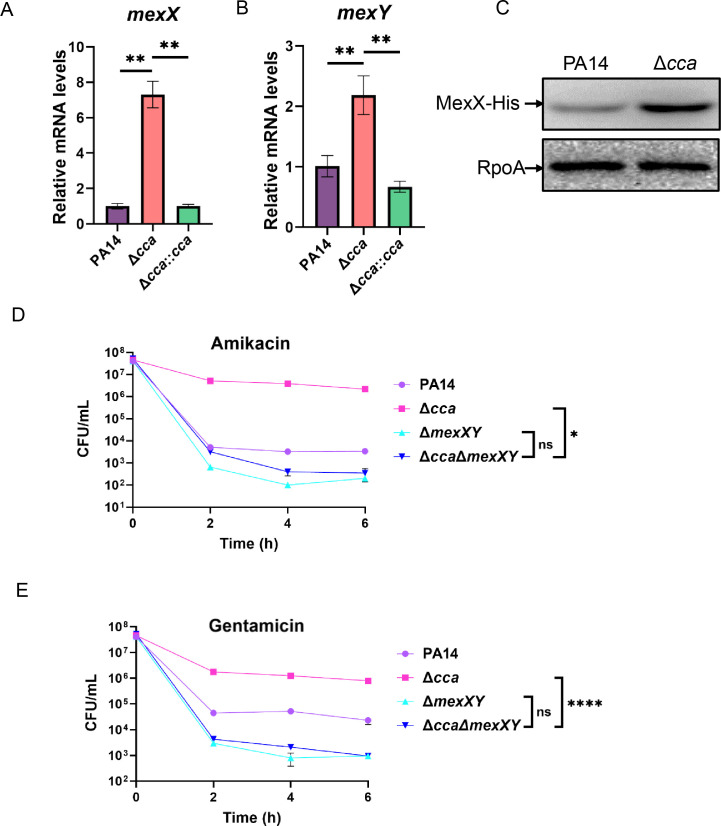 Fig 2
Fig 2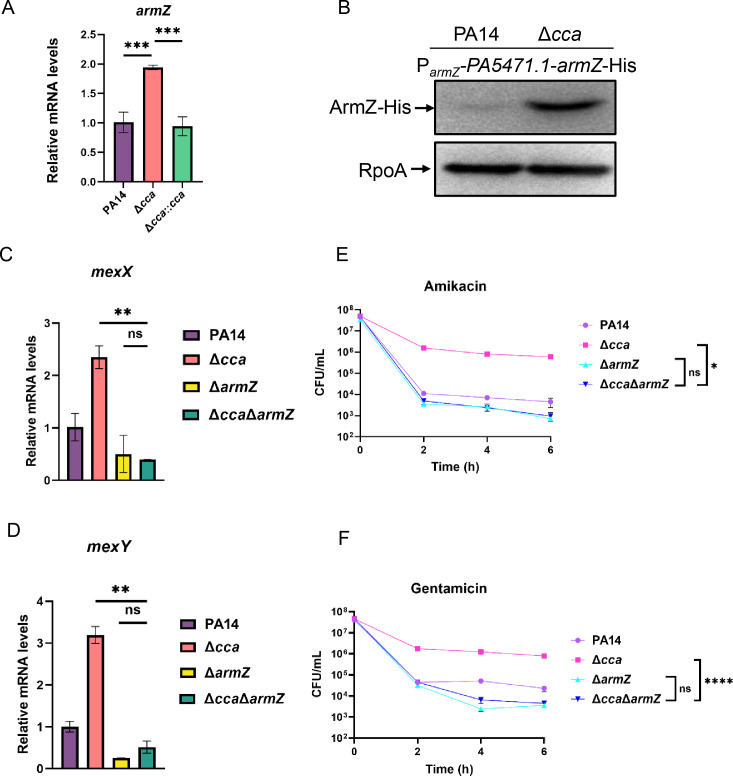 Fig 3
Fig 3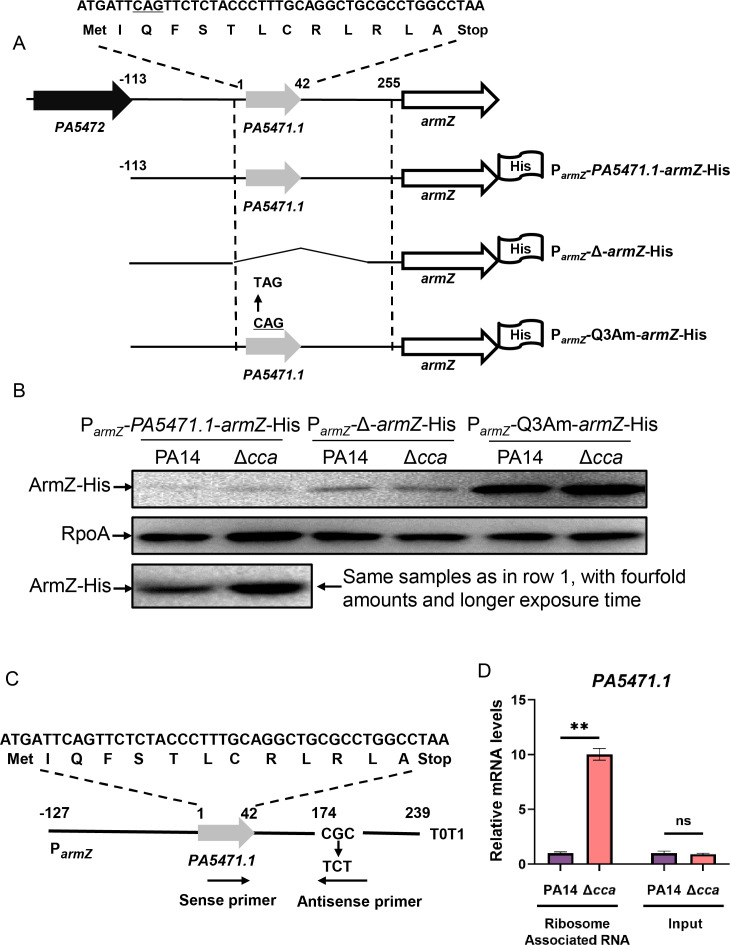 Fig 4
Fig 4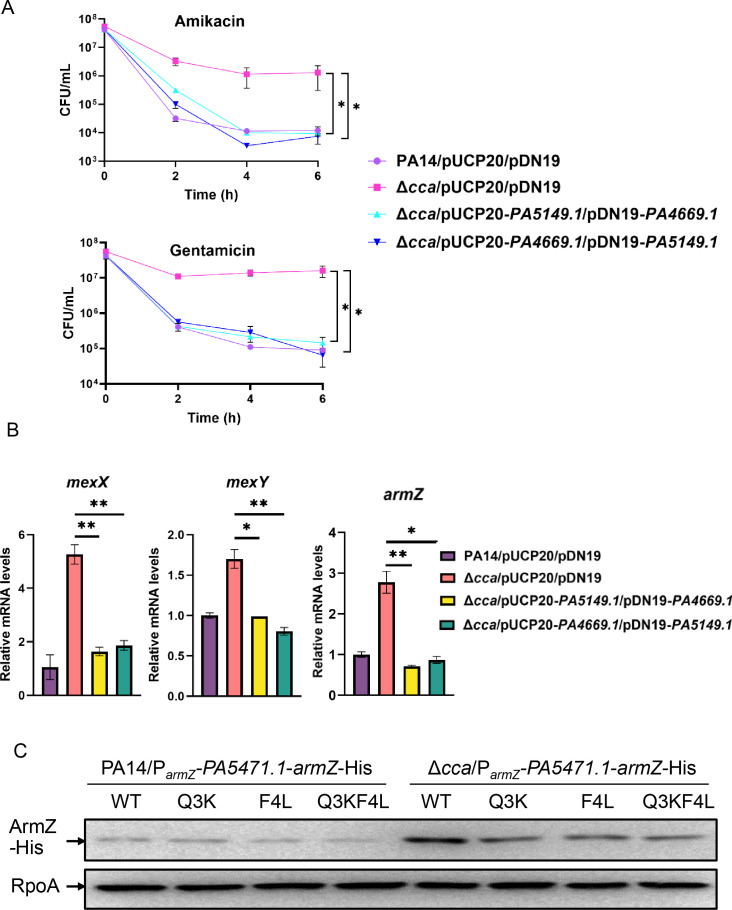 Fig 5
Fig 5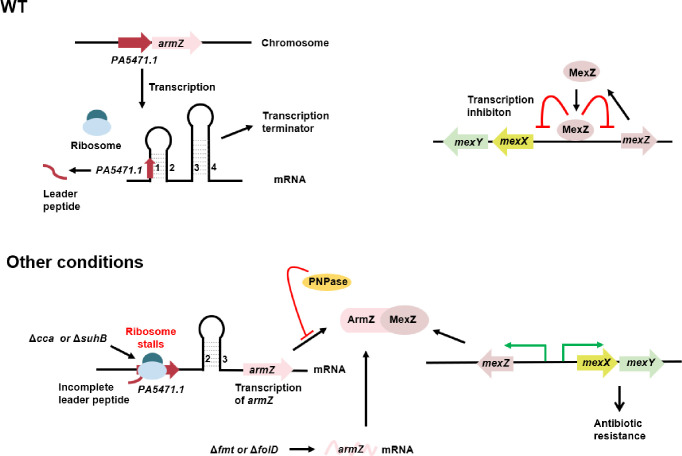 Fig 6
Fig 6| Strain | Amk | Gm | Tob | Neo | Mem | Cef |
|---|---|---|---|---|---|---|
| PA14 | 2.0 | 1.0 | 0.3 | 3.0 | 0.25 | 2.0 |
| Δ | 4.0 | 2.0 | 0.6 | 6.0 | 0.25 | 2.0 |
| Δ | 2.0 | 1.0 | 0.3 | 3.0 | 0.25 | 2.0 |
| Δ | 1.0 | 0.25 | 0.15 | 1.5 | ND | ND |
| Δ | 1.0 | 0.25 | 0.15 | 1.5 | ND | ND |
| Δ | 1.0 | 0.25 | 0.15 | 1.5 | ND | ND |
| Δ | 1.0 | 0.25 | 0.15 | 1.5 | ND | ND |
| PA14/pUCP20/pDN19 | 2.0 | 1.0 | ND | ND | ND | ND |
| Δ | 4.0 | 2.0 | ND | ND | ND | ND |
| Δ | 2.0 | 1.0 | ND | ND | ND | ND |
| Δ | 2.0 | 1.0 | ND | ND | ND | ND |
- —Science and Technology Committee of Tianjin
- —Science and Technology Committee of Tianjin
- —Fundamental Research Funds for the Central Universities, Nankai University
- —Fundamental Research Funds for the Central Universities, Nankai University
- —National Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —National Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —National Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA and protein synthesis mechanisms · Bacterial Genetics and Biotechnology · Antibiotic Resistance in Bacteria
INTRODUCTION
Pseudomonas aeruginosa is a gram-negative opportunistic human pathogen that can cause various acute and chronic infections in cystic fibrosis patients, immunocompromised individuals, and burn victims (1). It has intrinsic resistance and easily acquires resistance to a wide range of antibiotics, posing a great challenge to clinical treatment (2). P. aeruginosa possesses multiple mechanisms to defend against antibiotics, including multidrug efflux systems that export antimicrobial drugs outside bacterial cells.
Four multidrug efflux systems associated with resistance to antibiotics have been well elucidated in P. aeruginosa, including MexAB-OprM, MexCD-OprJ, MexEF-OprN, and MexXY-OprM (3). MexXY is an inducible efflux system that contributes to the bacterial resistance to aminoglycosides (4–6). Expression of the mexXY is regulated by its repressor MexZ, encoded by the immediately upstream gene mexZ (7). MexZ binds to the intergenic region of mexX-mexZ to repress expression of mexXY and mexZ itself (7, 8). PA5471, also named ArmZ for anti-repressor MexZ, functions as an anti-repressor by binding to the MexZ and reducing its DNA-binding ability (7, 9). Expression of armZ is induced by antibiotics, mediated by its leader peptide-encoding gene PA5471.1 upstream of the armZ (10). In the presence of ribosome-targeting antibiotics, such as aminoglycosides, the ribosome stalls on the mRNA of PA5471.1, altering the RNA secondary structure and leading to the transcription of armZ and, ultimately, mexXY, thereby conferring bacterial resistance to aminoglycosides (10).
tRNAs are adapter molecules that translate the genetic codes of mRNA into protein sequences. To be charged with their cognate amino acids, mature tRNAs carry the universally conserved CCA sequence at their 3′ termini (11). tRNA nucleotidyltransferase, also known as CCA-adding enzyme, is responsible for the synthesis and repair of the 3′-terminal CCA sequence (12). Most organisms rely on the posttranscriptional addition of the CCA triplet mediated by CCA-adding enzymes (13). However, some bacteria, like Escherichia coli, carry the CCA triplet in their tRNA genes (13). Therefore, acting as a tRNA repair enzyme, inactivation of the CCA-adding enzyme is not lethal for E. coli but only decreases the growth rate (14). While the function and structure of the CCA-adding enzyme in tRNA modification have been characterized (13, 15–17), other roles of the Cca in bacteria have not yet been elucidated.
Previously, we demonstrated that the ribosome-associated protein SuhB controls susceptibility to aminoglycoside antibiotics by modulating ribosome stalling and consequent expression of the MexXY multidrug efflux pump in P. aeruginosa (18). Polynucleotide phosphorylase (PNPase) controls bacterial tolerance to aminoglycosides by influencing the translation of armZ and thereby regulating MexXY (8). In this study, we found that mutation of the cca gene resulted in increased resistance and tolerance to aminoglycosides in P. aeruginosa. MexXY is upregulated in the Δcca mutant, which contributes to the increased resistance and tolerance to aminoglycosides. We further demonstrated that PA5471.1 is involved in the upregulation of armZ in the Δcca mutant. An increased amount of PA5471.1 mRNA was found to be associated with ribosomes purified from the Δcca mutant, indicating enhanced ribosome stalling. Furthermore, we found that the glutamine and phenylalanine at the third and fourth codons of PA5471.1, respectively, are involved in the Cca-mediated transcriptional upregulation of armZ and consequent upregulated expression of the mexXY efflux pump, as well as resistance and tolerance to aminoglycoside antibiotics.
RESULTS
Cca influences bacterial resistance and tolerance to aminoglycosides
To assess the role of Cca in antibiotic resistance to aminoglycosides and β-lactams, we examined the resistance of a Δcca mutant to amikacin, gentamicin, tobramycin, neomycin, meropenem, and ceftazidime. Compared with the wild-type strain PA14, the Δcca mutant exhibited the same susceptibility to meropenem and ceftazidime, but decreased susceptibility to amikacin, gentamicin, tobramycin, and neomycin, with twofold increases in minimal inhibitory concentrations (MICs; Table 1). Complementation with a cca gene restored susceptibility to these antibiotics (Table 1). Next, we determined the tolerance of PA14 and the Δcca mutant to aminoglycoside antibiotics. As shown in Fig. 1, mutation of the cca gene results in approximately 1,000-fold increased survival rates compared to the wild-type PA14 strain following treatment with amikacin, gentamicin, tobramycin, and neomycin, which were reversed by complementation with a cca gene. These results indicate a role for Cca in bacterial resistance and tolerance to aminoglycoside antibiotics.
*Cca influences tolerance to aminoglycoside antibiotics in P. aeruginosa. PA14, the Δcca mutant, and the complemented strain Δcca::cca were grown to an OD600 of 1.0 at 37°C and treated with 8 μg/mL amikacin (A), 4 μg/mL gentamicin (B), 2.4 μg/mL tobramycin (C), and 24 μg/mL neomycin (D). At the indicated time points, the survival bacterial numbers were determined by serial dilution and plating. *, P < 0.05; **, P < 0.01; and **, P < 0.001 by Student’s t-test.
Upregulation of mexXY contributes to the increased resistance and tolerance to aminoglycosides in the Δcca mutant
To understand the mechanism of Cca-mediated regulation of antibiotic susceptibility, we examined the expression levels of the mexX and mexY genes, which encode the multidrug efflux pump MexXY, a major determinant of aminoglycoside resistance in P. aeruginosa. Real-time qPCR results revealed significant upregulation of mexX and mexY in the Δcca mutant (Fig. 2). Complementation with a cca gene restored the relative mRNA levels of mexX and mexY in the Δcca mutant (Fig. 2). To further confirm the consecutive increase in MexX production due to gene upregulation, we utilized a C-terminal His-tagged mexX driven by its native promoter and examined the protein levels of MexX (8). Consistently, the MexX-His protein level was much higher in the Δcca mutant than in PA14 (Fig. 2C). Furthermore, deletion of mexXY in the Δcca mutant reduced the MICs and tolerance to the aminoglycosides to the same levels as those in the PA14ΔmexXY mutant (Table 1; Fig. 2D and E). These results demonstrate that upregulation of the mexXY efflux pump contributes to the increased resistance and tolerance to aminoglycoside antibiotics in the Δcca mutant.
*Upregulation of mexXY contributes to the increased tolerance to aminoglycoside antibiotics in the Δcca mutant. (A and B) Indicated bacterial strains were grown to an OD600 of 1.0 at 37°C, followed by RNA purification. Relative mRNA levels of mexX (A) and mexY (B) were determined by real-time qPCR with rpsL as an internal control. **, P < 0.01 by Student’s t-test. (C) Protein level of MexX-His in PA14 and the Δcca mutant with chromosomally integrated mexX-His. The bacterial cells were cultured to an OD600 of 1.0 at 37°C, and the MexX-His amounts were examined by western blot with RpoA as the loading control. (D and E) PA14, Δcca, ΔmexXY, and ΔccaΔmexXY were grown to an OD600 of 1.0 at 37°C and treated with 8 μg/mL amikacin (D) and 4 μg/mL gentamicin (E). At the indicated time points, the survival bacterial numbers were determined by serial dilution and plating. ns, not significant, *, P < 0.05 and ***, P < 0.0001 by Student’s t-test.
Upregulation of armZ contributes to the increased expression of MexXY and resistance/tolerance to aminoglycosides in the Δcca mutant
Transcription of the mexXY operon is directly repressed by its repressor MexZ (7). ArmZ controls mexXY transcription by interacting with and relieving the repression by MexZ (7). We then examined the transcription levels of armZ by real-time qPCR. As shown in Fig. 3A, the relative mRNA level of armZ was significantly increased in the Δcca mutant and was restored to the level seen in PA14 when complemented with a cca gene. To confirm the upregulation of armZ, we utilized a His-tagged armZ driven by its native promoter to determine the protein level of ArmZ. Consistent with the increased transcriptional level, ArmZ-His protein was increased in the Δcca mutant (Fig. 3B).
*Upregulation of armZ contributes to the increased expression of MexXY and tolerance to aminoglycosides in the Δcca mutant. (A, C, and D) Indicated bacterial strains were grown to an OD600 of 1.0 at 37°C, followed by RNA purification. Relative mRNA levels of armZ (A) mexX (C) and mexY (D) were determined by real-time qPCR with rpsL as an internal control. ns, not significant, **, P < 0.01 and ***, P < 0.001 by Student’s t-test. (B) Protein level of the ArmZ-His in PA14 and the Δcca mutant containing ParmZ-PA5471.1-armZ-His. The bacterial cells were cultured to an OD600 of 1.0 at 37°C, and the ArmZ-His amounts were examined by western blot with RpoA as the loading control. (E and F) PA14, Δcca, ΔarmZ, and ΔccaΔarmZ were grown to an OD600 of 1.0 at 37°C and treated with 8 μg/mL amikacin (E) and 4 μg/mL gentamicin (F). At the indicated time points, the survival bacterial numbers were determined by serial dilution and plating. ns, not significant, *, P < 0.05 and ***, P < 0.0001 by Student’s t-test.
To determine if the upregulated ArmZ contributes to the increased expression of MexXY, we deleted armZ in the Δcca mutant and determined the transcription levels of mexX and mexY. Real-time qPCR revealed that deletion of armZ reduced the mRNA levels of mexX and mexY in the Δcca mutant to the levels seen in the ΔarmZ mutant (Fig. 3C and D). Consistently, the MICs and tolerance to amikacin and gentamicin of the Δcca mutant were reduced by armZ deletion to the same levels seen in the ΔarmZ mutant of wild-type PA14 (Table 1; Fig. 3E and F). These results suggest that the upregulation of armZ contributes to the increased expression of mexXY operon and resistance/tolerance to aminoglycosides in the Δcca mutant.
Cca controls the transcription of armZ through its leader peptide PA5471.1
The expression of armZ is regulated through its leader peptide PA5471.1 (10). To explore whether PA5471.1 is involved in the Cca-mediated regulation of armZ, we deleted the PA5471.1 coding region in the ArmZ-His fusion construct, generating ParmZ-Δ-armZ-His (Fig. 4A). In another construct, a C was substituted with T at position 7 of PA5471.1 (encoding a glutamine, Q) to generate an amber mutation (designated ParmZ-Q3Am-armZ-His as shown in Fig. 4A), which had previously been shown to promote ribosome stalling and subsequently activate the transcription of armZ (10, 18). In agreement with the previous studies (10, 18), deletion or the point mutation of PA5471.1 largely increased the amounts of ArmZ-His (Fig. 4B). Notably, both mutations led to similar protein amounts of His-tagged ArmZ in wild-type PA14 and its Δcca mutant (Fig. 4B). These data indicate a role for PA5471.1 in the Cca-mediated regulation of armZ.
*Cca controls the expression of armZ through its leader peptide PA5471.1. (A) Structures of ArmZ-His fusions. The armZ, PA5471.1, and upstream regions were highlighted with numbers representing positions relative to the first base pair of the PA5471.1 start codon as 1. In ParmZ-Δ-armZ-His, the fragment from position 1 to 226 was deleted from ParmZ-PA5471.1-armZ-His. In ParmZ-Q3Am-armZ-His, a C at position 7 was substituted with a T, generating an amber mutation Q3Am. All the ArmZ-His fusion proteins were constructed in promoterless pUCP20 and transferred into wild-type PA14 and its Δcca mutant. (B) ArmZ-His protein amounts from the indicated constructs in PA14 and Δcca were examined by western blot assay. Samples from equivalent bacterial cell numbers were loaded onto sodium dodecyl sulfate-polyacrylamide gel electrophoresis gels and probed with an anti-His or anti-RpoA antibody. Row three shows the same samples as in row 1, but with fourfold amounts and longer exposure time in western blot for easier detection. (C) Structure of PA5471.1-T0T1. The first base pair of PA5471.1 start codon was designated as 1. Positions of real-time qPCR primers are indicated by arrows (18). (D) Quantification of ribosome-associated PA5471.1 mRNA. PA14 and the Δcca mutant containing pUCP24-rplL-His and pMMB67EH-ParmZ-PA5471.1-T0T1 were lysed by sonication and subjected to Ni-NTA chromatography, followed by RNA extraction. The purified RNA from lysed samples without Ni-NTA chromatography served as Input. The relative mRNA levels of PA5471.1 were examined by real-time qPCR with 16S ribosomal RNA (PA0668.1) as the internal control. ns, not significant, *, P < 0.01 by student’s t-test.
Mutation of cca enhances ribosome stalling at the PA5471.1 mRNA
Our previous study demonstrated that a suhB mutation increased ribosome stalling at the PA5471.1 mRNA, leading to increased transcription of armZ (18). The involvement of PA5471.1 in Cca-mediated regulation of armZ, as well as its tRNA nucleotidyltransferase function, suggests a role for Cca in modulating ribosome stalling at PA5471.1 mRNA. Therefore, a modified RNA-binding protein immunoprecipitation assay in combination with real-time qPCR was utilized to examine the amount of ribosome-bound PA5471.1 mRNA as previously described (18–20). Plasmids pMMB67EH-PPA5471 (armZ)-PA5471.1-T0T1 and pUCP24-rplL-His, previously constructed and utilized for the modified RNA-binding protein immunoprecipitation assay (18), were introduced into PA14 and its Δcca strains. In the plasmid pMMB67EH-ParmZ-PA5471.1-T0T1, two nucleotide substitutions were introduced into the PA5471.1 downstream region (Fig. 4C), ensuring that RNA transcribed from the chromosome would not be detected by real-time qPCR primers designed for the substituted region. Ribosomes from the two bacterial strains were isolated using Ni-NTA chromatography, and the associated RNA was purified and subjected to real-time qPCR. The mRNA levels of PA5471.1 produced from pMMB67EH-ParmZ-PA5471.1-T0T1 were similar between PA14 and the Δcca mutant (Input in Fig. 4D). However, a significantly increased amount of PA5471.1 mRNA was associated with ribosomes isolated from the Δcca mutant (Fig. 4D), indicating enhanced ribosome stalling at the PA5471.1 mRNA.
3Q4F of PA5471.1 is involved in the Cca-mediated resistance/tolerance to aminoglycosides
Ribosome stalling at the leader peptide PA5471.1 resulted in an increase in ArmZ levels in the Δcca mutant compared to those of PA14. As a tRNA nucleotidyltransferase, Cca is responsible for adding the CCA triplet at the 3′ termini of tRNAs to facilitate aminoacylation (21). It is possible that in the absence of Cca, some specific tRNAs lacking the 3′ CCA triplet have reduced or no aminoacylation, resulting in ribosome stalling at PA5471.1, increased armZ expression, and consequently increased resistance and tolerance to aminoglycosides. Therefore, we analyzed the tRNA coding genes in P. aeruginosa (www.pseudomonas.com). We found 63 tRNA encoding genes (Table S1), among which tRNA-Gln (glutamine) and tRNA-Phe (phenylalanine) were each encoded by only one gene, namely PA4669.1 and PA5149.1, respectively, which have only the CC sequence at their 3′ terminal ends. Glutamine is located at the third, while phenylalanine is located at the fourth position of the PA5471.1. To investigate if PA4669.1 and PA5149.1 are involved in the Cca-mediated regulation of resistance and tolerance to aminoglycosides, we overexpressed 3′ terminal A-added PA4669.1 and PA5149.1 in the Δcca mutant. Individual expression of the two modified tRNAs had no influence on the tolerance to aminoglycosides (Fig. S1). However, simultaneous expression of them (in two different plasmids, pUCP20 and pDN19) restored the MICs and tolerance to amikacin and gentamicin of the Δcca mutant to that of the wild-type PA14 strain (Table 1; Fig. 5A). In addition, simultaneous expression of the two modified tRNAs decreased the expression of mexX, mexY, and armZ (Fig. 5B).
*Glutamine at the third and phenylalanine at the fourth position of PA5471.1 are involved in the Cca-mediated tolerance to aminoglycosides. (A) Indicated bacterial cells were grown to an OD600 of 1.0 at 37°C and treated with 8 μg/mL amikacin or 4 μg/mL gentamicin. At the indicated time points, the survival bacterial numbers were determined by serial dilution and plating. *, P < 0.05 by Student’s t-test. (B) Indicated bacterial strains were grown to an OD600 of 1.0 at 37°C, followed by RNA purification. Relative mRNA levels of mexX, mexY, and armZ were determined by real-time qPCR with rpsL as an internal control. *, P < 0.05 and *, P < 0.01 by Student’s t-test. (C) Protein level of the ArmZ-His in PA14 and its Δcca mutant. Indicated bacterial cells were cultured to an OD600 of 1.0 at 37°C, and the ArmZ-His amounts were examined by western blot with RpoA as the loading control.
To further confirm that glutamine at the third and phenylalanine at the fourth (3Q4F) position of the PA5471.1 plays a role in the Cca-mediated regulation of ArmZ, we introduced a C to A point mutation at the 7th position (Q3K), a C to A point mutation at the 12th position (F4L), as well as simultaneous C to A at the 7th and C to A at the 12th position mutations (Q3KF4L) in PA5471.1 of the ParmZ-PA5471.1-armZ-His construct, resulting in Q3K, F4L, and Q3KF4L amino acid substitutions. Western blot assay was performed to examine the production of ArmZ. Consistent with a previous report that the Q3K mutation did not enhance armZ expression (10), individual Q3K and F4L substitutions did not increase the expression of ArmZ in the wild-type PA14 strain, suggesting that these two substitutions did not result in ribosome stalling at PA5471.1 (Fig. 5C). Importantly, the Q3K, F4L, and Q3KF4L simultaneous mutation decreased the expression of ArmZ in the Δcca mutant, while having no obvious effect in the wild-type PA14 strain (Fig. 5C). These results suggest that Q3 and F4 are involved in the Cca-mediated regulation of resistance and tolerance to aminoglycosides in P. aeruginosa.
DISCUSSION
The MexXY multidrug efflux pump is an important contributor to aminoglycoside resistance and tolerance in both P. aeruginosa reference strains and clinical isolates (22). Several regulators of MexXY have been found in P. aeruginosa, including regulators MexZ, ArmZ, SuhB, PNPase, Fmt, and FolD (Fig. 6) (7, 8, 10, 18, 23–25). In this study, we showed that tRNA nucleotidyltransferase Cca also controls the expression of MexXY in P. aeruginosa. Similar to SuhB and PNPase, Cca regulates the expression of MexXY by modulating the expression of ArmZ (Fig. 6).
Proposed model of mexXY regulation mediated by MexZ, ArmZ, SuhB, PNPase, Cca, Fmt, and FolD in P. aeruginosa based on this and previous studies (7, 8, 10, 18, 23–25). The armZ is regulated via a transcription attenuation mechanism. armZ and its leader peptide (PA5471.1) are transcribed under a constitutive promoter. In the wild-type strain, complete translation of PA5471.1 results in the formation of a transcription terminator (the hairpin structure formed by the base pairing between segments 3 and 4), which blocks the transcription of downstream armZ (10). Meanwhile, MexZ binds to the intergenic region and inhibits transcription of mexXY and mexZ (24). In the absence of suhB or cca, ribosome stalls at the PA5471.1 mRNA, which alters the RNA secondary structure and abolishes the formation of the transcription terminator, resulting in armZ transcription (18). PNPase represses translation of the armZ (8). The mutation of fmt and folD also leads to upregulation of armZ (25). The ArmZ protein binds to MexZ and releases its repression on its own gene and mexXY (7, 23).
tRNAs are initially transcribed as precursor tRNAs that undergo extensive processing at both the 5′ and 3′ ends to generate functional tRNAs that can be charged with their cognate amino acids at the 3′-ends (26). At the 3′-end, each functional tRNA includes the universally invariant CCA triplet that is either encoded in the tRNA coding gene or posttranscriptionally added by a highly specialized RNA polymerase, tRNA nucleotidyltransferase (CCA-adding enzyme) (26). In some bacteria, such as E. coli, Vibrio cholerae, and Yersinia pestis, the CCA 3′ sequence is already encoded in all tRNA genes (14, 27). Therefore, disruption of the cca gene in E. coli is not lethal, resulting only in a decreased growth rate (14). In the present study, we found that deletion of cca did not affect bacterial survival but led to a reduced growth rate in P. aeruginosa (Fig. S2). However, based on a previous study and the analysis of all the tRNA genes on the Pseudomonas website (www.pseudomonas.com) (27), not all cognate tRNAs for amino acids encode the 3′ CCA terminus. For example, PA5149.1, the only tRNA-Phe encoding gene, lacks the 3′ CCA sequence. Therefore, it is possible that other 3′ CCA-adding enzymes or other modification mechanisms exist for tRNA 3′ CCA modification and repair in P. aeruginosa.
Of note, besides their canonical role in mRNA translation, tRNAs have also been reported to perform additional functions in the regulation of gene expression (28). Uncharged tRNAs have been demonstrated to play a role in the bacterial stringent response (28, 29). Uncharged tRNAs function as effector molecules to regulate the expression of genes involved in aminoacyl-tRNA synthetases, amino acid biosynthesis, and amino acid uptake in Bacillus subtilis and other gram-positive bacteria (30). In addition, recent studies have revealed that tRNA modifications play important roles in regulating bacterial pathogenicity, virulence factors, and oxidative stress response (31–34). To our knowledge, this study provides the first evidence that tRNA nucleotidyltransferase Cca controls susceptibility to aminoglycosides through the MexXY efflux pump in P. aeruginosa. However, its global regulatory role in P. aeruginosa is still unknown and needs further investigation.
Interestingly, in some bacterial species, such as Bacillus halodurans, Deinococcus radiodurans, and Aquifex aeolicus, the incorporation of C and A residues is split between two tRNA nucleotidyltransferases (35–37). One CC-adding enzyme adds two C residues to the 3′-end of the tRNA, and the other A-adding enzyme incorporates the terminal A, whose collaboration leads to the complete CCA-end (36, 37). According to website prediction (www.pseudomonas.com) and a previous study, only one tRNA nucleotidyltransferase was found in P. aeruginosa (35). Therefore, it is possible that Cca acts as a tRNA nucleotidyltransferase with dual specificity for adding both C and A in P. aeruginosa.
Growth rate also influences bacterial susceptibility to antibiotics (38). The increased resistance and tolerance to aminoglycosides might be due to the decreased growth rate of the Δcca mutant. However, deletion of the mexXY efflux pump genes maintained the reduced growth rate (data not shown) while restoring the resistance and tolerance to aminoglycoside antibiotics of the Δcca mutant. Therefore, the decreased susceptibility to aminoglycosides of the Δcca mutant is not due to the reduced growth rate. In addition, simultaneous expression of the 3′ A-added PA4669.1 and PA5149.1 restored susceptibility to aminoglycosides but not the growth rate of the Δcca mutant (data not shown). The inability to complement the growth rate indicates that additional functions, other than A-addition to PA4669.1 and PA5149.1, affect bacterial growth in P. aeruginosa.
In summary, we found that tRNA nucleotidyltransferase Cca controls susceptibility to aminoglycoside antibiotics in P. aeruginosa and revealed its regulatory role on armZ expression and thus the mexXY efflux pump.
MATERIALS AND METHODS
Bacterial strains, plasmids, and primers
The bacterial strains, plasmids, and primers used in this study are listed in Tables S2 and S3. Bacterial strains P. aeruginosa (PA14) and E. coli used in this study were cultured in lysogeny broth (LB) medium (containing 1% NaCl, 1% tryptone, and 0.5% yeast extract; all [wt/vol]) or on LB agar plates (supplemented with 1.5% agar) at 37°C. For maintenance of plasmids within bacteria, appropriate antibiotics were supplemented in the medium at the following final concentrations: for P. aeruginosa, 50 μg/mL gentamicin, 150 μg/mL carbenicillin, and 50 μg/mL tetracycline; for E. coli, 10 μg/mL gentamicin, 100 μg/mL ampicillin, and 10 μg/mL tetracycline.
Construction of plasmids and deletion mutants
The cca gene was knocked out in frame by homologous recombination as previously documented (39). Upstream and downstream fragments (approximately 1,000 bp) of the cca gene were amplified by PCR with PA14 genomic DNA as a template and primers shown in Table S3. The PCR products were cloned into the EcoRI*-XbaI sites of pEX18Tc plasmid. Then the deletion construct pEX18Tc-cca was transferred into E. coli S17-1 by electroporation, followed by conjugal transfer into PA14. After that, single-crossover strains were selected on plates containing 50 μg/mL tetracycline and 25 μg/mL kanamycin. An overnight culture of the single-crossover strain was plated onto LB agar plates containing 7.5% sucrose to select double-crossover strains. Deletion of the cca gene was confirmed by PCR. Deletion of mexXY or armZ in PA14 or Δcca* was carried out with a similar procedure using the plasmid pEX18Tc-mexXY or pEX18Tc-armZ constructed in a previous study (40).
To complement the cca gene, the cca gene with its native promoter was amplified by PCR using specific primers (Table S3) with PA14 genomic DNA as the template. The PCR product was digested with EcoRI-BamHI and then cloned into pUC18T-mini-Tn7T (41). Then the resultant plasmid was introduced into the chromosome of the Δcca mutant by electroporation along with the helper plasmid pTNS3 (42). Insertion of the cca gene into the chromosome was verified by PCR amplification with primers PglmS-down and PTn7R (Table S3).
To construct plasmid pUCP20-PA5149.1, the fragment of PA5149.1 with a 3′ adenine addition was amplified by PCR using specific primers (Table S3) with PA14 genomic DNA as a template. The PCR products were digested with EcoRI*-Bam*HI and then cloned into pUCP20 plasmid. pUCP20-PA4669.1, pDN19-PA5149.1, and pDN19-PA4669.1 were constructed using a similar strategy.
To generate the construct of ParmZ-PA5471.1-armZ-His, the C-terminal His-tagged armZ with its native promoter region was PCR amplified with PA14 genomic DNA as a template and inserted into SacI*-BamHI sites of the promoterless pUCP20 (43, 44). To construct ParmZ-Δ-armZ-His, the 113 bp DNA fragment upstream of PA5471.1 and the C-terminal His-tagged armZ gene with its SD sequence were amplified using specific primers and ParmZ-PA5471.1-armZ*-His as a template (Table S3). The fragments were ligated by overlapping PCR and subsequently cloned into the SacI*-BamHI site of the promoterless pUCP20. For ParmZ-Q3Am-armZ-His, a point mutation (leading to the substitution of a glutamine to an amber stop codon, Q3Am) was introduced into primers ParmZ-Q3Am-armZ-UR and ParmZ-Q3Am-armZ-DF. Two DNA fragments were PCR amplified with ParmZ-Q3Am-armZ-UF/ParmZ-Q3Am-armZ-UR and ParmZ-Q3Am-armZ-DF/ParmZ-Q3Am-armZ-DR, ligated by overlapping and then cloned into SacI-BamHI site of the promoterless pUCP20. ParmZ-PA5471.1Q3K-armZ-His, ParmZ-PA5471.1F4L-armZ-His, and ParmZ-PA5471.1*Q3KF4L-armZ-His were constructed with similar procedures.
MIC determination
P. aeruginosa strains were subcultured in LB to an OD_600_ of 1.0 (5 × 10^8^ CFU/mL). MICs were determined with an initial inoculum 1 × 10^5^ bacterial cells each well in Cation-adjusted Mueller–Hinton Broth (Ca-MHB, QDRS Biotec, Qingdao, China) using a twofold dilution method in accordance with the guidelines of the Clinical and Laboratory Standards Institute (45). MICs were defined as the lowest concentration of antibiotic that inhibits visible growth following 24 h incubation at 37°C. Each experiment was repeated three times.
Bacterial survival assay
Bacterial cells were subcultured to an OD_600_ of 1.0 and 10-fold diluted into 2 mL of fresh LB medium. Then bacterial cells were treated with aminoglycoside antibiotics at final concentrations of twofold MIC (amikacin and gentamicin) or fourfold MIC (tobramycin and neomycin) at 37°C with shaking at 200 rpm. At indicated intervals (0, 2, 4, and 6 h), viable bacterial numbers were determined by serial dilution and plating assay. All experiments were performed in triplicate.
RNA isolation and real-time qPCR
Overnight bacterial cultures were 50-fold diluted into fresh LB medium and grown to an OD_600_ of 1.0. Total RNA was isolated utilizing the Bacterial Total RNA Isolation Kit (Zomanbia, Beijing, China), and cDNA was synthesized using a reverse transcriptase (Vazyme, Nanjing, China) and random primers (Vazyme, Nanjing, China). Real-time qPCR was performed using the ChamQ Universal SYBR qPCR Master Mix (TransGen Biotech, Beijing, China) and specific forward and reverse qPCR primers (Table S3) in a CFX Connect Real-Time system (Bio-Rad, United States). rpsL, the 30S ribosomal protein S12 encoding gene, was used as an internal control (46).
Western blot assay
Overnight bacterial culture was 50-fold diluted into fresh LB medium and grown to an OD_600_ of 1.0. Then samples from equivalent numbers of bacterial cells were collected by centrifugation, resuspended in 1× SDS loading buffer (62.5 mM Tris-HCl [pH 6.8], 10% [vol/vol] glycerol, 2% [wt/vol] SDS, 1% [vol/vol] β-mercaptoethanol, and 0.02% [wt/vol] bromophenol blue), boiled at 99°C for 10 min, and then separated by 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis gels. Proteins were transferred onto a polyvinylidene difluoride membrane and probed with a mouse monoclonal His antibody or RNA polymerase α antibody (RpoA, Biolegend). The signals were detected using the Immobilon Western Chemiluminescent HRP Substrate kit (Millipore). RpoA here serves as a loading control.
Detection of PA5471.1 mRNA associated with ribosomes
The amount of PA5471.1 mRNA associated with ribosomes was determined using a modified RNA-binding protein immunoprecipitation method followed by real-time qPCR as previously described (18–20). pMMB67EH-ParmZ-PA5471.1-T0T1 and pUCP24-rplL-His, previously utilized in the RNA-binding protein immunoprecipitation (18), were introduced into PA14 and its Δcca mutant. Bacterial cells were grown to an OD_600_ of 1.0 and collected by centrifugation. Ribosomes were purified by Ni-affinity chromatography as in a previous study with modifications (47). In brief, the collected bacterial cells were suspended in lysis buffer (150 mM NaCl, 20 mM Tris-HCl, 10 mM imidazole, 3 mM β-mercaptoethanol, 0.5% NP-40, pH 8.0) and lysed by sonication. Supernatants were collected after centrifugation and incubated with Ni-NTA agarose beads at 4°C for 1 h. The beads were washed four times with lysis buffer, and then the ribosomes were eluted using elution buffer, followed by RNA purification with an RNA Prep Pure Cell/Bacteria Kit (TransGen Biotech, Beijing, China). The amount of PA5471.1 mRNA was determined by real-time qPCR with specific primers (Table S3) using the 16S ribosomal RNA encoding gene PA0668.1 as the internal control for normalization.
Statistical analysis
Statistical analyses were conducted using GraphPad Prism 9.0 software. Two-tailed unpaired Student’s t-tests were employed to calculate P values, with statistical significance set at P < 0.05.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Balasubramanian D, Schneper L, Kumari H, Mathee K. 2013. A dynamic and intricate regulatory network determines Pseudomonas aeruginosa virulence. Nucleic Acids Res 41:1–20. doi:10.1093/nar/gks 103923143271 PMC 3592444 · doi ↗ · pubmed ↗
- 2Qin S, Xiao W, Zhou C, Pu Q, Deng X, Lan L, Liang H, Song X, Wu M. 2022. Pseudomonas aeruginosa: pathogenesis, virulence factors, antibiotic resistance, interaction with host, technology advances and emerging therapeutics. Signal Transduct Target Ther 7:199. doi:10.1038/s 41392-022-01056-135752612 PMC 9233671 · doi ↗ · pubmed ↗
- 3Poole K, Srikumar R. 2001. Multidrug efflux in Pseudomonas aeruginosa: components, mechanisms and clinical significance. Curr Top Med Chem 1:59–71. doi:10.2174/156802601339560511895293 · doi ↗ · pubmed ↗
- 4Jeannot K, Sobel ML, El Garch F, Poole K, Plésiat P. 2005. Induction of the Mex XY efflux pump in Pseudomonas aeruginosa is dependent on drug-ribosome interaction. J Bacteriol 187:5341–5346. doi:10.1128/JB.187.15.5341-5346.200516030228 PMC 1196038 · doi ↗ · pubmed ↗
- 5Westbrock-Wadman S, Sherman DR, Hickey MJ, Coulter SN, Zhu YQ, Warrener P, Nguyen LY, Shawar RM, Folger KR, Stover CK. 1999. Characterization of a Pseudomonas aeruginosa efflux pump contributing to aminoglycoside impermeability. Antimicrob Agents Chemother 43:2975–2983. doi:10.1128/AAC.43.12.297510582892 PMC 89597 · doi ↗ · pubmed ↗
- 6Rehman A, Patrick WM, Lamont IL. 2019. Mechanisms of ciprofloxacin resistance in Pseudomonas aeruginosa: new approaches to an old problem. J Med Microbiol 68:1–10. doi:10.1099/jmm.0.00087330605076 · doi ↗ · pubmed ↗
- 7Yamamoto M, Ueda A, Kudo M, Matsuo Y, Fukushima J, Nakae T, Kaneko T, Ishigatsubo Y. 2009. Role of Mex Z and PA 5471 in transcriptional regulation of mex XY in Pseudomonas aeruginosa. Microbiology (Reading) 155:3312–3321. doi:10.1099/mic.0.028993-019589837 · doi ↗ · pubmed ↗
- 8Fan Z, Pan X, Wang D, Chen R, Fu T, Yang B, Jin Y, Bai F, Cheng Z, Wu W. 2021. Pseudomonas aeruginosa polynucleotide phosphorylase controls tolerance to aminoglycoside antibiotics by regulating the Mex XY multidrug efflux pump. Antimicrob Agents Chemother 65:e 01846-20. doi:10.1128/AAC.01846-2033257447 PMC 7849007 · doi ↗ · pubmed ↗